Angela M.F. Fa ll eiros I
Elisa A. Gregório 2
AJ3STR .. \('T I'II\C;I)("YIII III ,\I()I'\II, II' I ,\R\\I ()I n/II/!({I SICU/II<({/S
11'\IlRII'II,l ILI I'IIHII'II lU. P\I/\lIIJ\I). The ullla'lruClural "h.,en·all<)l1 ,,( lhe hl.'IlHlCytl!\ \lI' n . . \(/("t'!/(I,.O/l" I(ln:tl.' ... h(l\\'l.'d Li 11 fl.'l"l.'llt ィエZ{ャhャcセ@ te lypc\ III \,H10U'" "dagl.' 01 intl.'raCl]OIl \\'1th Il11Cr()Orgalll ... Ill .... \\'111(11 ||Gj|セ@ lI11l.'rpn.:tcLl (I'" haCll.'rta. Tlll' Illlcroor galll\m ... \\'l.'l"e ()h...,cned l'llhel Ircc 111 lhe ィャNGエャャ|ャャセューィ@ (lI' l!ll!) thc IIl\CC( 11Ct1Hli. . NZ セ@ lc" Thc ha((Cni\ \\'cr\: dCll'ctl'd IlIO\tJ:- ]I!ll) lhe セイ[ャャQエゥャャャ」j@ te ... :llld pla:-.malpc:--1l' .... (Ill'
'pheruk cei" and (11L' UC1HIC} [oILl .... ílh(lIHe ... Cll(çd 1111]";1(.\ Illpla"IllH.': 1111Cr(lOrgillll\1ll\.
hUI lê .... , IreqUl'llll.\ . The lagoc.\tlc ((lp'lell.\ {lI lhe dlflerelll 1t1'll1ilC.\tl' (-'PC" I' di\clI\\l'd
KLY WOI{DS Lel',d"PICI'''. 1' ) 1,i1ld«e. f)iu/l'II<'u \U('I'/iUru!i\· . l1eI11"Clle.
ーィ。ァサIHセ@ エHIセャセN@ ultra .... truclun:
As células sanguíneas dos insetos. os hemócitos. desempenham papéis ele grande importância para a vida destes organismos, participando ativamente nos mecanismos de defesa contra elementos estranhos.
Os mecani smos de defesa dos insetos contra patógenos que pelletram sua hemocele tem sido considerados como parte dc um "sistcma imune". muito embora n<1o tenham sidos relacionados. até () prese nte. com a proeluç<1o de imunoglobulina s (Gl IPTA 1985; RATCI.II'I'I' 1'1 a/. 1985)
Esscs mecanismos de defcsa podem ser do tipo celular e humoral. As reações celulares. envolvcndo diretamenle os hemócitos , compreendem a coagu -lação da hemolinra , a ragocitose. a rormaç<1o de nódulos e o encapsulamento (GIÜ:GOIRJ: & GOFII\ET 1979; L..\CKIJ Ó 1983: Y .\GLR & KNIGJIT 1983: RIí',KI &
RUK I 1984: R;\TCLlI ;I'I: 1'1 a/. 1985; G()T? 1986). As rcações humorais. com o envolvimento inelireto dos hemócitos. caracteri/.amse pelo aparccimcllto na hemo -linra de ratores naturais ou induzidos tais como: aglutininas. ratores líticos. antibacterianos. lisosimas e a ativaç<1o do sistema de prorcnoloxidase (HOI ''!'VlA'iN
e/ a!. 1970: BLAl 'I.ATOt'\ & MO'-iI'EYSSIN 1977: PE\DL;\\D el a/. 1988: BROOK
-Vli\N 1'1 a/. 1989: Dl 't'\I'IJY & CII;\J)WICK 1989), ent re outros.
I) D"panalllel1lll de Ill slo logi«. CCl1lm lI.: HBセiQcj。L@ 8ioJlígic;". l 'l1l1'c r,idade ["adll,tI de LIlIllJnl1a. Caixa P'''lal WOJ. XC,O'i I -LJ70 1.'"1drI11'1. P«ral1,í. I1ra,iI.
2) Depanalllel110 de \J"rr" l"gla. II1"ilul" de iャャ ッ」ゥセ iQcBiGN@ t 'l1i\ er,,,Jade L"adll:tI Paull'l:l. Cal\a P"slal )1/1. IXC,JX-OtX) Iloluc« lu. S,1o I'«lllo. Br«,iI .
752 FALLEIROS & GREGÓRIO
A larva da DiOlrafo sacclwralis (Fabricius , 1794), conhecida como broca da cana-de-açúcar , é a principal praga da cultura da cana-de-açucar em nosso país. Seis tipos de hemócitos foram identificados em sua ィ・ュッャゥョャセGャ@ por BOMBONATO
( 1994). a saber: prohemócito. plasmatócito. granulócito, esferulócito. oenocitóide e célula vermi forme . Os mccani smos de defesa deste inselO e suas relações com os hemócitos. entretanto. não foram estudados até a presente data.
Neste trabalho. apresenta-sc achado ocasional de fagocitose e destruição de patógenos . por diferentes tipos de hcmócitos em larvas de D. socc!wralis.
MATERIAL E MÉTODOS
Os excmplares de larvas de D. sacclwra/is utilizados em nosso trabalho foram obtidos do Laboratório de Controle de Pragas da Usina da Barra Açúcar e Álcool S.A. , Barra Bonita (São Paulo).
As larvas da D. sacchora/is foram cultivadas em tubos de vidro previamente estcrilizados, eontendo dieta artificial desenvolvida por HENSLEY & HAMMO'.'D (1968), mantidas em temperatura de 25 -27°C e umidade relativa do ar de
aproximadamente 70 %.
A hemolinfa de 20 larvas com 24 dias de desenvolvimento foi colet ada com auxílio de pipeta Pasteur, através de punção na região abdominal. imediatamente eolocada em solução anti -coagulante de insetos (LtóONARD fi a/. 1985) e centri-fugada a 5000rpm por cinco minutos. O sedimento obtido foi fixado por duas horas em glutaraldeído a 2,5 '1r. em tampão fosfato O, I M pH 7,2 e pós-fixado em tetróxido
de ósmio a I % no mesmo tampão, sendo posteriormente processado rotineiramente
para microscopia eletronica de transmissão.
RESULTADOS
A análise morfológica ultracstrutural dos hemócitos ele D. saccharalis mostrou, cm algumas elas preparações. microorganismos quc foram interprctados como bactérias , livrcs ou em vár ios estágios dc interação com diferentes tipos dc hemócitos circulantcs. As larvas infcctadas não aprcscntavam qualqucr alteração anatómica ou comportamental que as distinguissem das não contaminadas.
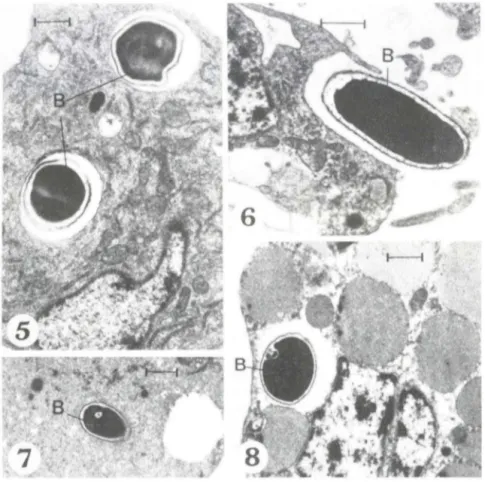
Os microorganismos foram observados preferencialmente nos granu lócitos, que mostraram vacúolos em diversos estágios ele digestão intracelular (Figs 1-4). Notamos adesão de grânulos citoplasmáticos. próprios deste tipo celular, ao vacúolo digestivo (Fig. 3) , bem como vár ios patógenos destruídos (Fig. 4).
Imagens ele fagocitose elas bactérias pelos plasmatóciros foram observadas (Fig. 6); os patógenos intracelulares não estavam alteraelos. nestas células (Fig. 5)
Os oenocitóides (Fig. 7) e os esferulócitos (Fig. 8) também apresel1laram microorgan ismos i nt raci toplasmát icos. embora com menor frequêneia. Nessas células. entretanto, nem se mpre foi possível visualizar membrana vacuolar nítida, separando o citoplasma celular cio patógeno, que se apresentava íntegro.
Flgs 1-4. Granulócitos em diferentes fases da fagocitose de bactérias (8). (1) Emissão de
pseudópodos (barra = 0, UセャュI[@ (2) bactéria íntegra em vacúolo (barra = 1 ,Ofim); (3) detalhe
de bactéria morta em vacúolo, notar imagem de fusão de grânulo com membrana vacuolar
(seta) (barra = 0, 5fim); (4) bactéria morta (barra = 1 ,Ofim).
DISCUSSÃO
A fagocitose, considerada como um complexo e sofisticado processo biológico, é o mais comum dos mecanismos de imunidade celular, formando a primeira linha de defesa em muitos insetos (RIZKI & RIZKI 1984; RATCLlFFE &
ROWLEY 1979; RATCLlFFE el alo 1985; ANDERSON & CHAIN 1987 ; FENOGLIO et
alo 1993).
Um grande número de microorganismos como vírus, fungos, bactérias e protozoários são fagocitados pelos hemócilos , tanto in vivo como in vitro
(RATCLlFFE & ROWLEY 1975 1979; RATCLlFFE & WALTERS 1983; CHIANG el alo
1988 ; RAHM ET-ALLA & ROWLEY 1990, entre outros). Os plasmatócitos (PL) e/ou
granulócitos (GR) são os hemócitos apontados como as células fagocitárias , na grande maioria dos insetos (ROWLEY & RATCLlFFE 1976; RIZKI & R1ZKI 1984 ; G UPTA 1985 ; KAA Y A el alo 1986; SANTOS el alo 1990; BAINES el al. 1992 , EHLERS
el alo 1992; HUNG et alo 1993) , por possuirem a capacidade de reconhecer, aderir
e englobar elementos estranhos ao organismo (RATCLl FFE & ROWLEY 1979).
Alguns trabalhos mostraram que os PL são as células predominantemente envolvidas no processo de fagocitose (RIZKI & RIZKI 1984 ; KAA YA el alo 1986 ;
SANTOS el al. 1990; BAIN ES el alo 1992), enquanto outros autores descrevem como células fagocíticas predominantes os GR, (ROWLEY & RATCLlFFE 1976; EHLERS
el alo 1992 ; KURIHARA el alo 1992; HUNG el alo 1993). Entretanto, ANGGRAENI
754 FALLEIROS & GREGÓRIO
& RATCLIFFE (1991), trabalhando com monocamada de hemócitos purificados de
Galleria lIlellonella (Linnaeus, 1758) (Lepidoptera, Pyralidae) coberta por
suspen-são bacteriana, observaram que, embora a ingestão fosse atributo dos PL, a atividade fagocitária máxima, nessas células, só foi obtida após a readição de GR ao meio, ilustrando a necessidade da cooperação célula-célula neste processo.
Outros tipos de hemócilOs também foram apontados como fagócitos em
insetos. NEUWlRTH (1974), trabalhando com inoculação de partículas inertes em
larvas de Calpodes elhlius (Stoll , 1782) (Lepidoptera, Pyralidae), conclue que os
GR e os oenocitóides (OE) são as células fagocíticas; os PL e os esferulócitos (ES) não fagocitam, embora partículas possam ser encontradas nas proximidades do primeiro tipo celular.
Figs 5-8. Diferentes tipOS de hemócitos com bactéria (8) intracitoplaslllática. (5) Plasma
-tócito contendo bactérias íntegra s em vacúolo definido (barra = O, UセャャャャI[@ (6 ) plaslllatócito
englobando bactéria livre (barra = 1 LoセュI[@ (7) oenocitóide (barra = 1 LPセャiョI[@ (8) esferulócito
(barra = 1 LoセュI N@
Em hemoliní"a de larvas de D. saccharalis com 24 dias de desenvolvimento , as observações mostram que além dos PL e dos GR, também os ES e os OE se encontram, aparentemente, envolvidos na fagocitose de patógenos.
Estudos iII \'i\'O e iII \'irra sugerem que a fagoci tose em inse tos, a exemplo dos vertebrados, ocorre em quatro estágios: quimiotaxia. aderência , ingestão e morte do agcnte estranho (RATe!.II'!'!: & ROWLl,y 1979: RATCLlFIT 1986). Nas observações nota -se imagens que apontam para o G R e PL como células fagoei -tárias: hactérias fo ram vistas aderidas. sendo envoltas por pseudópodos c Íntegras em vacúolos intracitoplasmáticos. Entrctanto. somcnte o GR evidenciou bactérias em vá rios graus de dege neração. dentro dos \·acúolos d igest i vos.
A habilidade de destruir os patógenos parece estar na dependência da liberação de elvimas lisosomais para os vaeúolos de fagocitose (RATCLlFIT & Ro\\'u :Y 1979; KROSClII\SKI & RE:-.JWRi\:"JTí'. 1988: FEi\'OGLlO ef (II. 1993). Nesse sentido. estudos citoquÍmicos ultraestruturais de detecção de atividade li sosomal de\'erão ser cfetuados nos diferentes tipos de hemóeitos . para comprovar com certeza aqueles que detém essa capacidade de destruição direta de patógenos.
Com relação aos ES e OE. pode-se obsen·ar bactérias no citoplasma. po rém nem se mpre com membrana \'acuolar evidente. Segundo GI"P"r,\ (1985). estes últimos tipos celulares podem. oeasionalmente, serem apontados como fagócitos. porém, este fato não é demonstrado pela grande maioria dos aLIlores. A hipótese de uma super infccçao bacteriana, que levasse ao co mprometimento pass ivo de todas os tipos de hemócitos. inclusi\'e ES e OE. pode ser descartada. uma vel que não se pôde distinguir. pelo aspecto macroscó pico das larvas. aquelas que esta\'am contaminadas.
Não se conseguiu detectar bactérias no citoplasma de células vermilormes ou de prohemócitos. Esses tipos celulares aparecem em quantidade redu/ida na hemolinra da broca da cana com 2-1 dias de dese!1\·ol\'imento (80f\1130:\ ,\TO 199-1). Assim. ii dificuldade natural de dCletú -los nas prcparações para microscópio eletrónico. não permite exclUI -los da relação dos hemóeitos fagocitúrios em D. sl/(e/lllm/is.
/\ capacidade ragocitária dos diferentcs tipos de helllócitos da D .
socc!/Omlis dever<Í ser expcrilllelllalmellle avaliada, bem como o papel de cada um
deles nos mecanismos de defesa deste inscto .
. I\GRADECIMU\TOS. 0, \ャlャiBiB\セG@ LLセイ。、lG」」ャャQ@ ii, Sr;I'. :\'Llrta Eukda LII1<) Peru é f\.larr;t
I kklla vturellu. pL'lu prnCé.\\all1elllot':CIlIC<l d" materral. Tr;lh;llIlil parclalmcllle fill<lllcl;ldu pelo C:\PlI Il'rnc. I.+OI7l)(')()-'))
REFERÊNCIAS BIBLIOGRÁFICAS
A\DERSON. R. & B.M. 01.\1"'. 1987 Mac rophage func tio ns in insects, p.77-RR.
/11: A.P. Gl 'PTA (Ed.). Hcmoeytic 3ml hum oral immunit y in arthropods .
New York. Willey- Interscicnce. XII + 5:l:\p.
Ai\'Cj(iR.\E \1. T. & N.A. R·\TC LlFrT. 1991. Studies on cell-cell co-operation during
756 FALLEIROS & GREGÓRIO
phagoeytosis by puriried hacmoeyte populations 01' the moth. Cal/eria
II/el/one//a. Jour. Insect Physiol. 37 (6): --l53---+60.
BAI:'-JES , D.; T. DESANTIS & R.G.H. DOW:'-JER. 1992. Octopaminc and 5-hydro-xytryptamine enehance the phagoeytic and nodule rormation aetivities 01' eoekroaeh (Perip/ane{([ all/erical/a) haemoeytes. Jour. lnsect Physiol. 38 (II): 905-914.
beaャ i latoセ L@ J. & M. MO\,PEYSSI N. 1977. Ultrastrueture et eytochimie des hémoeytes d'AlIlheraea pemri (Guér) (Lepidoptera, Attacidae). II Cellules à sphcrules et oenocytoides. Riol. Cell. 28: 13- 18.
BOMBONATO, M.T.S. I 99--l. Análise morfológica c quantitativa dos hemócitos de larvas de Diatraea saccliaralis (Lepidoptera: Pyralidae) frente ao pa-rasitóide Metagol/isty/ulIl lIlin el/se (Diptera: Tachinidae). Disse rtação ele Mestrado , não publicada , Instituto de Bioeiências, Universidade Estadual Paulista , Botucatu , 9Op.
BROOKMAN, J.L.; N.A. RATCL1I:I'E & A.F. ROWLEY. 1989. Stuelies on the act ivat ion 01' the prophenoloxidase sys telll 01' insects by bacterial cell wall components. Insect BiochclIl . 19 : --l7-57.
Ctlli\ NG, A.S.: A.P. Gl 'PTA & S.S H/d\'. 1988 . Arthropod immune sys tell1: I. Comparati \'e I igth and elcctron mieroscopie accounts 01' i mll1unocy tes anel Olhe r hemoeytes o
r
Blallel/a gemwllica (Dict yo ptera: Blattell idae). JOllr. 1\ lorphol.198 : 257 -267.
DL':-.!PIIY , G. B. & J .S. CII ADW ICK. 1989. EtTccts 01' selected earbohyelrate anel the eontribution 01' the prophenoloxielase easeade system to lhe adhesion 01' slrains 01' Pselldoll/OIlOS aemginoso anel Prolells II/irabi li S 10 hemoc ytes 01' noni mll1une larval Ca/leria lIIe//oneL/a. Cano Jour. I\licrohiol. 35: 524-527.
EIILERS. D.; B. ZOSEL; W. MOlIRIG; E. KAL ISC IIKE & M. EIIL ERS. 1992. COlllparison 01' iII ri\'() anel in \'i!ro phagoeYlosis in Cal/eria lIIel/olle/la L.. Parasitol. Res . 78 : 354 -359.
FENOG LlO .
c.;
P. BER'-!ARDI:-.!I & M.V. GERVASO. 1993 . CYloehemieal charae-leril.ation 01' the hellloc ylcs 01' Lellcophaea lIIaderoe (Dietyoptera: Blaberoielea). Jour. I\lorphol, 218: 115-126.GOTZ, P. 1986. Meehanisllls oCencapsulalion in Diplera hosts. p.I -19. /n: A.M. LACKI E (Ed.). Immune Illcchanisllls in invertehrate vectors. Ox forel, Clareelon Press, V + 285[1.
GIÜ:COIRE. C.H. & G. GOI'FI :-.IET. 1979. Conlroversies aboul lhe coaguloeyles, p.189-229. /n : A.P. GUPTA (Ed.). Insect hemocytes: ele"c1opment, forms , fllnctions anel techniqucs. Cambridge. Cambridge Univcrsil)' Press. XVI + 61--lp.
Gl ·I'TA. !\.P. 1985 . Cellular elcmellls in lhe hemolymph. p.-.f02---l--l-.f . III : G.A. KERKl 'T & L. I. GIL13ERT (Eds). Comprehensive insect physiology hiochem-istry anel pharmacolog)'. Oxford. Pergamon Press. vaI. 3. XV I+625p. HEi\S1.EY . S.D. & A.M. H:\:-"I\I10:\D JR. 1968. Laboratory lechniques ror rearing
the sugareane borcr on an art i ricial diel. Jour. Econ. Entolllol61: 1742- 17--l3. HOl'l'iYIA NN. J.A. ; A. PORTE & P . .I0LY. 1970. On lhe localization
dase aetivity in eoagulation or LoclIsra lIIigra/oria (L.) (Orthoptera). C.R. Hebd. Séanees Aead. Se. Paris. 270: 629-631.
HUNG, S.Y.; D.G. BoueIAs&A.J. VEY. 1993. EffeelofBealll'eriabassianaand
COlidida albicall.\" on lhe eellular defense response 01" Spodop/era e.\"iglla. Jour. Invertebr. Pathol. 61: 179- 187.
KAAYA. G.P.; N.A. RATCLlFFE & P. ALEM U. 1986. Cellular and humoral
de fcn ses or C/ossil/a: reactions against baeteria, lrypanosomes and
experimental illlplant s. Jour. Mcd. Entomol. 23: 30-43.
KROC IIINSKL J. & L. RFNWRANTZ. 1988. Delermination or the pH values insidc the cligestive vacuoles of hemocytes rrom M\"/i/II.\" edlllis . .lour. Invertebr. Pathol. 51: 73-79.
Kl'RIIIARA, Y.; T . SIIIMAZU & H. WAGO. 1992. Classiriealion or heIlloeytes in the eOllllllon eutworm, Spodop/I!ra Ii/lira (Lepieloptera: Noeluidae) I. Phase Illieroseopic stuely. Appl. Entomol. Zool. 27 (2): 225-235.
LM'K 110 , A. M. 1983. Elleets of sUbSlratLIlll wettabi I ity ancl eharge on adhesion iI/ "il/'o ancl eneapsulation iII l'i"o by inseet haellloeyles . .lour. Ccll Sei. 63: 181 - 190.
LEONARD, c.; K. SODERIIALL & N.A. RATCLlFFE. 1985. Studies or pro-phe-noloxiclase and protease or B/a!Jems crallii{er haemoeytes. Inseet Bioehcm. 15: 803-810.
NEl"WIRTII, M. 1974. Granular hemoeytes, lhe Illain phagocytie blood eclls in Ca/podes e/hlills (Lepidoptera, Hesperi ielae). Cano .lour. Zool. 52: 783-784. PL"iDLAND , J.C.; M.A. HEAT&D.G. BO lle IAS. 1988. Funelions ora galaetose-bincling leelin rrom Spodop/I!I'O exiglla larval haemolymph: opsoni/.ation or blastospores rrom enlomogeneous hyphomyceles . .lour. Inseel Physiol. 34: 533 -540.
RMIMET-AuA M. & A.F. ROWI.LY. 1990. Studies on lhe cellular delcnsc
reaetions 01' the macieira eockroach, Lellcopha(!(/ I/lOderae: iII "i/ro phagocy-losis or ditlcrent strains 01' Bacilllls cerells aml thcir elTect on hemocyte viability. Jour. Invertehr. Pathol. 55: 350-356.
RATCLlFFE, N.A. 1986. Inseet eellular immunity anel the reeognition of roreign -ness, p.21 -43. /11: A.M. LACKIE (EcI.l. IJl1Jl1une Jl1cehanisms in invertc-bratc vcelors. Oxforcl, C1areclon Press, V
+
285p.R/\TCUFFE, N.A. & A.F. ROWLEY. 1975. Cellular defense reaetions 01' insect hellloeyles iII "il/'o: phagoeytosis in a new suspension eulture system . Jour. Invertchr. Pathol. 26: 225 -233.
1979. Role 01' hemoeyles in defense againsl biologieal agents, p. 331-414.
III: A.P. Gl'PTA (EeI.). Inseel hcmoeytes: devclopment, I'orms, funetions
allCl teehniqucs. Cambriclge, Cambriclge University Press. X V 1+ 614p. RATCUFIE, N. A. & J. B. W AL TERS. 1983. Stuclies on the iII l'il'o eellular reaetions
01' inseets: elearanee or pathogenie ancl non-pathogenie baeteria in Cal/eria lIIellollel/a larvae . .lour. Inscel Physiol. 29: 407-415.
RI\TCLlFFE, N.A.; A.F. ROWLLY; S.W. FITZGERALD & c.P. RIIODFS. 1985, Invertebrate immunity: basie coneepts and reeent aclvanecs. Int. Rcv. Cytol.
758 FALLEIROS & GREGÓRIO
97: 183-349.
RIZKI, T. M. & R. M. RIZKI. 1984. The eeliular defense 01' DrosophiLa lIIe/aI10-gasler, p.579-604. III: R.C. KING & H. AKAI (Eds) . Inseet UItl'astrueture, New York, Plenull1 Press, vol. 2, XXV+62.tp.
ROWLEY. A.F. & N.A. RATCLlFFE. 1976. The granular eelis 01' Gal/eria lIIel/a -lIel/a during elotting phagoeYlie reaelions iII \'i!ro. Tissuc Cell, 8 (3) : 437 -446. SANTOS, R.R.; M .M.G. ESTE; E.S. GARCIA & P. AZA MBUJA. 1990. Binding siles for IgG -Fc in hell10eYle aclherenl ecIls 01' hemalophagous bugs (Rhodllills proLi\lIs). I\lcm, Inst. Oswaldo Cruz 85 (4): 469-470.
Yi\GER, J.F. & H.H. KNIGHT. 1983. Mieroseopie observalion bloocl eoagulalion in several clitlerenls speeies 01' inseels. Jour. Ent. Soe. AmeI'. 26: 591 -602.
Recebido em 1 O.VI.1995. 。cヲNセiio@ em 211111996