CRISTIANA ENGELMANN
FATORES QUE INFLUENCIAM A NEOFOBIA
ALIMENTAR EM FÊMEAS E FILHOTES DE SAGUIS
(
Callithrix jacchus
)
Natal 2010
CRISTIANA ENGELMANN
FATORES QUE INFLUENCIAM A NEOFOBIA ALIMENTAR EM FÊMEAS E FILHOTES DE SAGUIS (Callithrix jacchus).
Natal 2010
Dissertação apresentada à Universidade Federal do Rio Grande do Norte para obtenção do título de Mestre em Psicobiologia.
iii Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de Biociências
Engelmann, Cristiana.
Fatores que influenciam a neofobia alimentar em fêmeas e filhotes de sagüis (Callithrix jacchus) / Cristiana Engelmann. – Natal, RN, 2010.
42 f. : Il.
Orientadora: .Profa. Fívia de Araújo Lopes.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1. Callithrix jacchus – Dissertação 2. Sagui – Dissertação. 3. Neofobia alimentar – Dissertação. I. Lopes, Fívia de Araújo. II. Universidade Federal do Rio Grande do Norte. III. Título.
Título: “Fatores que influenciam a neofobia alimentar em fêmeas e filhotes de
saguis (Callithrix jacchus).”
Autor: Cristiana Engelmann
Data da defesa: 08-10-2010
Banca Examinadora:
______________________________________________________________________ Prof. Francisco Dyonisio C. Mendes (UnB)
______________________________________________________________________ Profª Maria de Fátima Arruda (UFRN)
v
“A vida, Senhor Visconde, é um pisca-pisca. A gente nasce, isto é, começa a piscar. Quem pára de piscar, chegou ao fim, morreu. Piscar é abrir e fechar os olhos - viver é isso. É um dorme e acorda, dorme e acorda, até que dorme e não acorda mais. [...] A vida das gentes neste mundo, senhor sabugo, é isso. Um rosário de piscados. Cada pisco é um dia. Pisca e mama; pisca e brinca; pisca e estuda; pisca e ama; pisca e cria filhos; pisca e geme os reumatismos; por fim pisca pela última vez e morre.
- E depois que morre? - perguntou o Visconde. - Depois que morre, vira hipótese. É ou não é?”
AGRADECIMENTOS
Agradeço a todos aqueles que de alguma forma contribuíram para esta minha conquista
e em especial:
Ao meu pai Lotário pelo apoio constante e, particularmente, pelo investimento e
cuidado que sempre teve com a minha dedicação ao estudo, e por ser o grande
responsável pelo amor que tenho à leitura e aos conhecimentos. A minha mãe, Ursula,
que além de ser uma grande amiga, sempre foi meu exemplo de perseverança, de
dedicação e por ter me ensinado sobre a importância em conduzir tudo que se faz com
amor. Agradeço a ambos por toda ajuda, compreensão e apoio sempre.
A minha irmã Juliana e ao meu cunhado Gilberto, fontes de constante inspiração, pelo
incentivo, por todo o auxilio prestado durante meus estudos e pela confiança depositada
em mim, com certeza sem eles o caminho teria sido muito mais difícil.
Aos meus sobrinhos, Tomas e Alice, motivo constante de alegrias em todos os
momentos, amo demais!!
A minha irmã Daniela, e meu sobrinho Gabriel, que apesar da saudade que sinto,
sempre estiveram presentes em pensamento.
A todos os meus familiares e amigos distantes pelo carinho prestado, mesmo que via
Embratel.
A minha orientadora, Fívia Lopes, por quem tenho grande admiração e carinho,
agradeço pela paciência, por suas valiosas orientações, e pelo exemplo, que espero
seguir por toda minha vida profissional.
Aos meus amigos, pelos momentos de descontração, por me aturarem nos meus
períodos de “nada vai dar certo”, e que apesar das minhas aporrinhações estiveram
vii Ao meu amigo Bruno por ter me estimulado tantas vezes a continuar, e pelos valiosos
conselhos.
Ao meu colega Ricardo pelos puxões de orelha que me estimularam inclusive a
conseguir minha primeira bolsa de mestrado.
Ao Felipe pela paciência e pela grande e valiosa ajuda na analise dos dados.
A todos os meus colegas de mestrado.
A todos os professores do departamento de fisiologia, em especial Maria Emília
Yamammoto, Arrilton Araújo e Fátima Arruda pelas ótimas aulas e pelo exemplo
enquanto profissionais.
Aos funcionários do Núcleo de Primatologia, Edinólia, Zé Rubens, Geniberto, Tota,
Luís Veríssimo, Francisco, Márcio André e ao veterinário José Flavio, pelo apoio
técnico, por serem tão prestativos, enfim, por terem me auxiliado muito na execução
desse trabalho.
A funcionária Rose pelos cafés, tão importantes nas manhãs de coleta.
Ao CNPQ pela bolsa concedida, que me permitiu que esta pesquisa fosse realizada.
A Darwin (não seria eu se não houvesse esse agradecimento).
SUMÁRIO
Pág. RELAÇÃO DE ILUSTRAÇÕES...
RESUMO... ABSTRACT...
viii
x
xi
1. INTRODUÇÃO... 1
2. OBJETIVO GERAL... 9
2.1. OBJETIVOS ESPECÍFICOS... 9
3. HIPÓTESES E PREDIÇÕES... 10
4. MATERIAL E MÉTODOS... 13
4.1 Animais e Alojamento... 13
4.2 Dieta dos Animais... 14
4.3 Escolha dos Alimentos... 14
4.4 Procedimento... 15
4.4.1 Fêmea Gestante... 16
4.4.2 Filhotes... 16
4.5 Comportamentos Registrados... 17
4.6 Análise dos dados coletados... 18
5. RESULTADOS... 19
5.1 Fêmea ... 19
5.2 Filhotes... 23
6. DISCUSSÃO... 28
7. CONCLUSÃO... 33
ix
LISTA DE ILUSTRAÇÕES
Pág.
Figura 1: Mediana e desvio interquartílico da quantidade de itens consumidos em
relação a gestação...
Figura 2: Mediana e desvio interquartílico da latência para o consumo do primeiro
item, independente da qualidade gustativa, em relação a gestação...
Figura 3: Mediana e desvio interquartílico da latência para o consumo do primeiro item
apresentado em relação a qualidade gustativa deste item (doce ou não doce)...
Figura 4: Mediana e desvio interquartílico da quantidade de itens consumidos por
fêmeas em relação à qualidade gustativa do alimento. ...
Figura 5:. Média e intervalo de confiança (95%) para os itens consumidos doces e não
doces em relação a gestação...
Figura 6: : Média e intervalo de confiança (95%) para a latência em segundos para o
consumo do primeiro item...
Figura 7: Mediana e desvio interquartílico da latência para o consumo do primeiro item
em relação à experiência intra-uterina (consumo do item pela mãe durante o período
gestacional)...
Figura 8: Mediana e desvio interquartílico da quantidade de itens consumidos por
filhotes em relação ao consumo prévio da mãe destes mesmos itens durante o período
gestacional...
Figura 9: Mediana e desvio interquartílico da quantidade de aproximações do filhote
em relação ao consumo prévio do item pela mãe durante o período
gestacional...
Figura 10: Mediana e desvio interquartílico da latência para o consumo do primeiro
item relacionado ao sexo do filhote...
Figura 11: Mediana e desvio interquartílico da quantidade de itens consumidos em
relação ao sexo do filhote...
Figura 12: Mediana e desvio interquartílico da quantidade de aproximações do
alimento apresentado em relação ao sexo do filhote...
Tabela I: Caracterização dos animais experimentais de acordo com as condições de
alojamentos, presença de gestação, tipo de alimento oferecido às fêmeas e aos filhotes,
bem como o sexo destes últimos. ...
Tabela II: Tabela nutricional dos alimentos selecionados para o experimento, para 100g,
conforme a Tabela Brasileira de Composição de Alimentos- TACO, UNICAMP (Lima et al.
2006)...
26
26
27
13
xi
RESUMO
Considerando as constantes mudanças ambientais, a capacidade de introduzir
alimentos novos na dieta é essencial para a sobrevivência de animais onívoros. Para a
otimização do forrageio e diminuição dos riscos de intoxicação, é necessário ao animal
detectar sinais que indiquem quais itens são adequados ou não para o consumo.
Investigamos alguns fatores que interferem na resposta aos alimentos não familiares,
modulando comportamentos neofóbicos em sagüis Callithrix jacchus, primatas nativos
do nordeste brasileiro, animais onívoros e generalistas, porém cautelosos na ingestão
de itens desconhecidos. Foi analisada a influência da qualidade gustativa dos alimentos
(doce e salgado), da gestação, e do sexo no comportamento alimentar e na resposta
neofóbica desses animais. Selecionamos inicialmente 10 fêmeas de origem cativa,
sendo cinco grávidas e cinco não. As fêmeas apresentadas aos itens doces ingeriram
maior quantidade do que aquelas apresentadas ao item salgado, estando ou não
gestantes. Contudo, fêmeas grávidas se mostraram menos neofóbicas para ambos os
alimentos, embora demonstrassem forte neofilia para os doces. Verificamos então a
influência da alimentação das grávidas no comportamento alimentar pós-natal de seus
filhotes fêmeas e machos. Observamos 10 filhotes: um grupo cuja mãe teve contato
com o alimento na gestação e um cuja mãe não tivera esse contato. No primeiro grupo
o alimento foi mais facilmente aceito pelos filhotes, sugerindo que a neofobia e a
preferência alimentar possuam influência pré-natal. Além disso, filhotes fêmeas
ingeriram mais alimentos e mostraram-se menos neofóbicas que filhotes machos,
diferença já observada em adultos da mesma espécie. Nossos resultados sugerem que
o comportamento de baixa neofobia aos alimentos doces apresentado pelas fêmeas
possa ser adaptativo, e tenha conferido uma maior aptidão àquelas que o
ABSTRACT
Considering the constant environmental changes, the ability to introduce new
food items in the diet is crucial to omnivore animal survival. For optimal nourishment
and lessening of intoxication risks, the animals must detect signs that indicate which
items are adequate for their intake. We investigate some factors that interfere in the
responses to non familiar food, modulating their neophobic behavior, of marmosets
Callithrix jacchus, an omnivore and generalist primate, native to Northeast Brazil,
known for being cautious in ingesting not known food. We analyzed the influence of
food taste (sweet or salty), pregnancy and sex in feeding behavior and neophobic
responses in these animals. 10 captive females were first selected, 5 of them being then
pregnant. The females, pregnant or not, ate more when presented to the sweet items
than to the salty ones. Pregnant females, however, themselves were less neophobic to
both tastes, being also strongly neophilic to the sweets. We verified then the influence
of nourishment during pregnancy on young males and females post natal feeding
behavior. We observed 10 young divided in two groups, one whose mother ate that
food item during pregnancy and one whose mother had no contact to it. In the first
group that food was more easily accepted by the young, suggesting that neofobia and
feeding behavior had a pre natal influence. Female young also ingested more food and
were less neophobic than males, a difference already observed in behavior of adults of
these specie. These results suggest that the low neophobic behavior to sweet food
showed by females can be adaptive, and might have bestowed more fitness to those
1
1. INTRODUÇÃO
A introdução de novos itens alimentares na dieta de animais onívoros é um fator de
grande importância na estratégia alimentar. Considerando que o ambiente natural está
em constantes mudanças, a capacidade de inovar e de aceitar alimentos novos e
diversificados desempenha um papel essencial para a sobrevivência de um animal em
vida livre, pois a flexibilidade dos comportamentos permite aos animais lidarem com as
mudanças ambientais (Day et al. 2003, Fragazy & Mason 1978). Entretanto, a ingestão
de itens desconhecidos também apresenta riscos potenciais como o envenenamento. Este
dilema quanto a ingestão de alimentos foi denominado por Rozin (1966, citado por
Wise, 2009) de “Dilema do Onívoro”. Com a finalidade de otimizar a eficiência de
forrageamento e também de evitar a intoxicação, os animais devem estar, portanto, aptos
para detectar, discriminar e aprender sobre os sinais que indicam o que poderia se
apresentar como um alimento em potencial (Laska & Freist 2006).
A resposta aos alimentos não familiares pode ser classificada como neofilia ou
neofobia, sendo a neofilia caracterizada como uma espontânea atração ao novo (Voelk et
al. 2006), enquanto a neofobia alimentar é caracterizada pela evitação de alimentos
novos (Visalberghi 1994). A neofobia é uma forma eficiente de reduzir os riscos de
ingestão de itens tóxicos (Galef 1996, Johnson 2007, Visalberghi 1994, Yamamoto &
Lopes 2004). De uma forma geral, a maior parte das espécies é neofóbica perante
alimentos desconhecidos evitando-os completamente ou alimentando-se inicialmente
apenas com pequenas porções (Day et al. 2003, Visalberghi & Fragazy 1995, Voelk et
São frequentes os estudos referentes ao tema da neofobia alimentar. Entretanto,
artigos nessa área costumam prestar pouca atenção na origem dos alimentos utilizados,
uma vez que algumas características como textura e consistência poderiam afetar a
resposta neofóbica (Johnson 2007). Muitas das informações obtidas nesses estudos se
baseiam unicamente na qualidade nutricional dos itens utilizados ignorando o lado
sensorial do indivíduo. Porém essas qualidades nutricionais são abstratas demais para o
entendimento não humano. Sendo assim, para entender o motivo da preferência de um
item específico seria favorável criar associações entre a qualidade nutricional e o sistema
sensorial, considerando, por exemplo, o tato, o olfato e a sensação da textura dos
alimentos (Dominy et al. 2001).
A qualidade gustativa caracterizada pelo sabor de um alimento pode ser também
um forte fator de influência na hora da escolha dos alimentos pelos animais. A maior
parte dos vertebrados possui sistema gustativo que responde às quatro qualidades
gustativas (Carlson 2001) e diferenciam o sabor doce do salgado, por exemplo. Os
receptores para doce são, claramente, detectores de alimento e a maior parte dos
alimentos doces, tais como frutas, podem, em geral, ser ingeridos com segurança
(Carlson 2001, Ramirez 1990). Ainda segundo Carlson (2001), a maioria das espécies
animais prontamente comerá substâncias que contenham gosto doce ou salgado;
contudo, evitará substâncias mais ácidas ou amargas. Portanto, podemos esperar que
diferentes características dos alimentos realmente despertem mais ou menos
comportamentos qualificados como neofóbicos.
Quando nos referimos especificamente a primatas, podemos observar que o seu
comportamento de seleção de alimentos de fato sugere que estes utilizam receptores de
3 prováveis itens alimentares (Laska 2001, Laska et al. 2003). De acordo com testes
realizados por Visalberghi (2003) utilizando macacos-prego (Cebus apella), ao
receberem a oportunidade de escolha entre sete alimentos desconhecidos, a preferência
foi positivamente correlacionada aos alimentos contendo maiores quantidades tanto de
glicose quanto de frutose. Johnson (2007) prevê ainda que exista uma preferência inata
para doces e que isso poderia alterar as respostas neofóbicas de um animal. Porém essa
afirmação se mostra contrária à definição proposta por Wise (2006), que sugere que
unicamente as manifestações cognitivas e comportamentais de fome sejam inatas, mas a
preferência por tipos específicos de alimento não.
Dentro do grupo dos primatas, podemos considerar o sagui comum, Callithrix
jacchus como uma espécie perfeitamente adequada ao estudo de neofobia alimentar,
pois apesar de serem onívoros e terem a capacidade de utilizar uma grande variedade de
itens na sua alimentação, apresentam comportamentos de cautela frente a itens
desconhecidos. Além disso, essa espécie tem se destacado como sujeito experimental
nas mais diversas áreas da ciência. Possivelmente esse fato se deve a algumas
características particulares, como o fácil manejo e a fácil manutenção em cativeiro, além
da alta taxa de fecundidade.
O sagui comum é um pequeno primata nativo do nordeste brasileiro que vive em
grupos sociais normalmente formados de 3 a 15 animais, contendo indivíduos adultos
reprodutores e adultos não reprodutores, subadultos, juvenis e infantes (Stevenson &
Rylands 1988). De acordo com estudos em cativeiro e também em condição natural, os
saguis do gênero Callithrix podem viver em famílias, agrupamento de indivíduos
aparentados e prole, ou em grupos, consistindo de animais aparentados e mais um ou
1978). Koening e Rothe (1991) discutem que assim como os micos, estes animais se
distinguem de outros grupos de primatas por serem bem descritos pelo cuidado
cooperativo, podendo esse comportamento ser realizado inclusive por indivíduos não
aparentados; esse auxílio é frequente inclusive na provisão de itens alimentares para os
infantes (Ferrari 1988, Koening & Rothe 1991, Rylands 1996). Tal provisão pode
representar em alguns casos mais de 17,5% da comida ingerida pelos filhotes (Voelk et
al. 2006).
Assim como outros Calitriquídeos, os saguis são animais onívoros e possuem um
amplo repertório alimentar (Ahlborn & Rothe 1999, Castro 2003). Boa parte de sua
alimentação é obtida a partir de exsudados extraídos de troncos e galhos de árvores, mas
também se alimentam de frutas e insetos (Caldwell & Whiten 2004, Rylands & Faria
1993) e até mesmo de pequenos vertebrados. Segundo Ferrari (1996), a alimentação por
exsudado provê uma fonte estável de recurso e é conseguida por esses animais devido a
uma especialização que consiste na habilidade de provocar a produção de goma em
vegetais. Porém a obtenção desta forma de recurso pode ser muito dispendiosa do ponto
de vista energético; sendo assim, há preferência por outros alimentos, como frutas e
vegetais, quando estes se encontram disponíveis (Rylands & Farias 1993).
As fêmeas dessa espécie apresentam várias peculiaridades no que se refere ao
período gestacional. Primeiramente é importante considerar que o sistema de
acasalamento pode se mostrar flexível. Contudo, normalmente há apenas uma fêmea
reprodutora, sendo esta a dominante do grupo. As demais fêmeas, subordinadas, tendem
a ter sua reprodução suprimida e costumam atuar auxiliando no cuidado dos infantes ou
não (Abbott et al. 1993, Yamamoto et al. 2009). As fêmeas dominantes apresentam
5 ano, com nascimento normalmente de gêmeos a cada gestação (Torri et al. 1996),
embora casos de nascimento de trigêmeos e quadrigêmeos já tenham sido observados,
especialmente em cativeiro (Rutherford & Tardif 2009). O período gestacional é de
aproximadamente 148 dias e as fêmeas apresentam receptividade sexual ao longo de
todo o ciclo ovariano, mesmo durante esse período, diminuindo essa frequência apenas
após as primeiras 6-8 semanas de gestação (Hearn et al. 1978, Hearn 1983). Esse fato
pode ser em decorrência do desenvolvimento fetal tardio, ou seja, o feto apresenta pouco
crescimento no decorrer dos primeiros meses, havendo uma intensificação (boom) desse
crescimento nos últimos dois meses (Chambers & Hearn 1985). Durante o período de
baixo desenvolvimento fetal a fêmea costuma estar, inclusive, amamentando a prole
anterior.
No que se refere à introdução de novos itens alimentares à dieta, os saguis têm sido
descritos como cautelosos. No entanto, alguns fatores parecem exercer forte influência na
neofobia de C. jacchus, como o sexo do animal, a sua faixa etária ou a sua posição
hierárquica dentro do grupo. De acordo com Yamamoto et al. 2004, existem diferenças
entre os sexos no que diz respeito à resposta dirigida aos alimentos desconhecidos. As
fêmeas apresentam uma maior prontidão na interação com estes itens. Isso sugere que
machos sejam mais cautelosos perante a possibilidade de inserção de alimentos novos, ou
seja, apresentem maior neofobia. Além disso, fêmeas aparentemente apresentam maior
motivação para encontrar potenciais recursos alimentares do que machos (Michels 1998).
Essa prontidão das fêmeas na aquisição de alimentos parece se justificar pelo elevado
investimento que sua reprodução demanda (Lacerda 2004).
A interação social tem se mostrado crucial para o aprendizado alimentar do infante
resposta neofóbica, tal como proposto por Vitale e Queyras (1997). Isso pôde ser
evidenciado nos estudos realizados por Voelk et al. (2006) que demonstraram que na
ausência de adultos os indivíduos infantes se recusavam mais a comer itens
desconhecidos, ou apresentavam uma latência maior para a ingestão, investigando mais do
que quando o alimento era previamente conhecido. Porém na presença de membros do
grupo essa latência se apresentou diminuída, e os itens não familiares foram mais
prontamente aceitos, sugerindo que a facilitação social atua na diminuição da neofobia em
infantes. Experimentos realizados por Yamamoto e Lopes (2004) com adultos, subadultos
e indivíduos juvenis em C. jacchus, parecem estar de acordo com essa afirmação,
sugerindo ainda que haja diferenças no comportamento alimentar correspondente a faixa
etária. Estas autoras evidenciaram que, quando isolados, os adultos de saguis ingerem
mais alimentos novos, e iniciam a alimentação mais rapidamente do que os indivíduos
jovens em isolamento. Porém, quando em seu grupo familiar, a ingestão de alimentos
novos em indivíduos jovens aumenta, sugerindo que a neofobia para estes é maior quando
se encontram sozinhos e que a facilitação social exerce uma forte influência no que se
refere ao sucesso da introdução de novos itens alimentares na dieta.
A familiarização com o alimento, parece também influenciar a neofobia
alimentar, visto que animais podem “aprender” sobre a palatabilidade de um alimento
novo por meio da experiência direta (Galef 1996, Vitale & Queyras 1997). É provável
que, assim como a influência social, a experiência individual possibilite ao indivíduo o
aprendizado do que comer, ou seja, a neofobia é atenuada pelo contato constante com o
alimento, desde que a ingestão deste não cause nenhuma forma de desconforto
(Visalberghi & Adessi 2006). Esse aspecto foi testado em C. jacchus por Lacerda
7 animais tendem a detectar pistas confiáveis sobre a palatabilidade e a não toxidade,
passando a reduzir a cautela em relação ao alimento apresentado.
Outro mecanismo que pode atuar na compreensão e na redução da neofobia
alimentar sugerido a partir de pesquisas, sobretudo com primatas, é a influência do
ambiente intra-uterino. Como é característico entre os mamíferos, nos primatas os
embriões se desenvolvem no interior do útero materno, estabelecendo dessa forma uma
comunicação materno-fetal direta através tanto de hormônios quanto de nutrientes
(Rosenblatt 1967). Os saguis, da mesma forma que outros primatas antropóides,
possuem placenta hemomonocorial, o que significa que existe uma única camada
coriônica separando a circulação materna da circulação do feto (Luckett 1974). Por esse
motivo o tecido fetal é constantemente nutrido, de forma direta e intensa, pelos gases e
nutrientes contidos no sangue materno (Goehl & Tardif, 2009). Em várias outras
espécies de mamíferos, incluindo os seres humanos, muito já fora descrito sobre a
aprendizagem fetal a partir da dieta materna na gravidez. Schaal et al. 2000,
demonstraram que bebês humanos aparentemente são capazes de reconhecer odores
após o nascimento, quando esses foram previamente inseridos na alimentação materna
durante o período de gestação. Dessa forma, podemos supor que a alimentação da fêmea
durante esse período deve influenciar fortemente a alimentação da futura prole, podendo
inclusive aumentar ou diminuir a neofobia. Logo, poderíamos questionar até que ponto a
neofilia aos alimentos doces é de fato inata ou se ela se desenvolve como um resultado
do comportamento alimentar materno, durante o período da gestação.
Diferentes fatores podem influenciar, portanto, o comportamento alimentar em
filhotes de C. jacchus. A forte influência exercida pela presença do grupo social na
bastante evidente. Porém fazem-se necessários mais estudos sobre a origem dessa
preferência, assim como a respeito dos aspectos neofóbicos e neofílicos, sem a
9
2. OBJETIVO GERAL
Caracterizar o comportamento de neofobia alimentar em fêmeas de Callithrix
jacchus e investigar a influência do comportamento alimentar da fêmea durante o
período da gestação na aceitação de alimentos novos pela sua prole após o nascimento.
2.1 OBJETIVOS ESPECÍFICOS
I. Investigar a reação da fêmea grávida a alimentos novos em comparação a fêmeas
não grávidas;
II. Investigar se a neofobia apresenta-se reduzida para alimentos com alto teor de
açúcar em comparação a alimentos com baixo teor de açúcar, e verificar se esta
diminuição apresenta-se tanto para a fêmea grávida quanto para a não grávida;
III. Investigar a frequência de comportamentos neofóbicos em filhotes, considerando a
alimentação materna durante a gravidez;
IV. Verificar se ocorre diferença no que se refere aos comportamentos neofóbicos dos
3. HIPÓTESES E PREDIÇÕES
Esperamos verificar as seguintes hipóteses:
Hipótese 01
O período de gestação influencia o comportamento neofóbico das fêmeas de Callithrix jacchus.
Predição
Fêmeas gestantes apresentam menos comportamentos neofóbicos do que as fêmeas fora
do período de gestação, tanto para alimentos doces quanto para não doces.
Justificativa
Ao apresentar alimentos não familiares aos indivíduos, fêmeas em gestação devem
responder mais prontamente a estes itens. Isso pode ser esperado, pois a gestação é um
período muito dispendioso energeticamente e a não aceitação de um recurso alimentar
pode acarretar em mais danos do que benefícios.
Hipótese 02
A qualidade gustativa do alimento influencia o comportamento de neofobia alimentar em fêmeas de Callithrix jacchus.
Predição
A fêmea de sagui comum é menos neofóbica a alimentos com maior teor de açúcar do
que a alimentos salgados.
11 Esse fato pode ser esperado devido à maior aceitabilidade de alimentos doces de uma
forma geral, visto que em vida livre, alimentos doces, como frutos, são em sua maioria
palatáveis.
Hipótese 03
A experiência intra-uterina influencia a neofobia/neofilia de filhotes após o nascimento.
Predição
Filhotes cuja mãe recebera um determinado item alimentar, independente da qualidade
gustativa deste item, durante a gravidez devem apresentar menos comportamentos
neofóbicos a este alimento.
Justificativa
Ao inserir um alimento novo na alimentação da fêmea durante os últimos momentos da
gravidez, este mesmo alimento será, posteriormente, aceito mais facilmente pelos
filhotes em decorrência da transmissão de informações sobre os alimentos ainda no
ambiente intra-uterino. De maneira geral, os comportamentos que caracterizam a
neofobia quanto a alimentos novos serão menos frequentes em filhotes cujas mães
tiveram contato com esse mesmo alimento, o oposto sendo verdadeiro (filhotes cujas
mães não receberam determinados alimentos durante o período de gestação serão mais
neofóbicos).
Hipótese 04
Predição
Os filhotes cujas mães tiveram acesso a uma dieta com maior teor de açúcar terão maior
facilidade para aceitar alimentos doces do que aqueles cujas mães receberam uma dieta
com restrição desse alimento.
Justificativa
Como é nossa expectativa que fêmeas grávidas apresentem neofobia diminuída para
itens doces em relação as não grávidas, e consumam grande quantidade desses itens
quando oferecido, acreditamos que tal comportamento terá um efeito sobre o filhote.
Esta diminuição do comportamento neofóbico no filhote deve ser esperada, pois durante
a gestação há troca de substâncias entre mãe e feto, o que forneceria pistas ao filhote
quanto a propriedades dos alimentos oferecidos. Considerando que a neofobia para itens
doces é menor durante a gravidez, há um alto consumo desses itens quando disponíveis.
Se durante a gravidez o consumo de doce é aumentado espera-se que o filhote aceite
com facilidade o item doce, tornando essa baixa neofobia a doces uma característica não
inata, mas sim de aprendizado intra-uterino. Dessa forma a preferência por alimentos de
qualidade gustativa doce poderia ser favorecida por essas pistas.
Hipótese 05
Os comportamentos de neofobia se apresentam diferentes para cada sexo já na infância.
Predição
Desde infantes as fêmeas de sagui comum já apresentam menos comportamentos
13 Justificativa
Ao serem isolados do grupo, os filhotes do sexo feminino apresentariam menos
comportamentos neofóbicos aos itens novos apresentados, ingerindo uma maior
quantidade e apresentando uma latência de ingestão menor do que os filhotes do sexo
masculino. Em adultos isso já foi demonstrado e essa característica possivelmente é
4. MATERIAL E MÉTODOS
4.1 Animais e Alojamento
Para a realização deste experimento selecionamos 10 fêmeas adultas reprodutoras
de sagui comum, C. jacchus, estando cinco destas em grupos familiares e cinco apenas em
casais (Tabela I). Dentre as fêmeas, cinco foram analisadas durante a gestação e cinco fora
deste período. Os animais eram de origem cativa, nascidos e mantidos até a idade adulta em
cativeiro, e pertencentes ao Núcleo de Primatologia da UFRN, sob condições naturais de
temperatura, ciclo claro-escuro e umidade.
Tabela I: Caracterização dos animais experimentais de acordo com as condições de alojamentos, presença de gestação, tipo de alimento oferecido às fêmeas e aos filhotes, bem como o sexo destes últimos.
Fêmea
Condição de alojamento/ nº de
animais Condição da fêmea Alimento oferecido à fêmea Filhote testado*
Sexo do filhote
1 Casal/ 2 indivíduos Grávida Queijo Sim Macho e fêmea
2 Família/ 6 indivíduos Grávida Queijo Sim Macho e fêmea
3 Casal/ 2 indivíduos Não grávida Queijo Não -
4 Família/ 4 indivíduos Não grávida Queijo Não
-5 Família/ 6 indivíduos Grávida Pêssego Sim Macho e fêmea
6 Casal/ 2 indivíduos Grávida Pêssego Sim Macho e fêmea
7 Casal/ 2 indivíduos Não grávida Pêssego Não -
8 Família/ 6 indivíduos Não grávida Pêssego Não
-9 Casal/ 2 indivíduos Não grávida Pêssego Não -
10 Família/ 8 indivíduos Grávida Pêssego Sim 2 machos
15 A seleção dos animais foi realizada com base na probabilidade de gestação, sendo
escolhidas fêmeas reprodutoras que se encontravam juntamente com machos reprodutores.
Os filhotes de algumas fêmeas reprodutoras também foram objeto de estudo, ao
completarem dois meses e meio, período no qual inicia a alimentação do infante com itens
sólidos (Yamamoto 1993). No total foram observados 10 filhotes de 5 fêmeas.
Os grupos foram acompanhados nas próprias gaiolas-viveiro, compostas por dois
espaços adjacentes interligados por uma comunicação (janela) que permite a separação dos
animais para a realização do procedimento com os filhotes. As fêmeas que se encontravam
em casais ou em grupos também foram mantidas em suas gaiolas-viveiro originais, sem a
necessidade do uso de um espaço adjacente visto que não foram isoladas.
4.2 Dieta dos animais
Os animais utilizados eram mantidos com uma dieta balanceada incluindo
suplementação vitamínica e protéica diária. A alimentação era oferecida duas vezes por dia,
pela manhã e à tarde. Pela manhã, a alimentação era composta basicamente por um mingau,
feito com banana, pão, ovo, suplemento vitamínico e água. No período da tarde, os animais
receberam frutas e um item protéico (ovo, frango ou tenébrio).
4.3 Escolha dos Alimentos
Para a realização do experimento foram selecionados dois itens alimentares não
conhecidos pelos animais. O primeiro era um item alimentar doce, sendo esse pêssego em
alimento fora previamente observada em animais, machos e fêmeas adultos, isolados com a
finalidade de verificar a palatabilidade e aceitação dos itens, e assim a possibilidade de
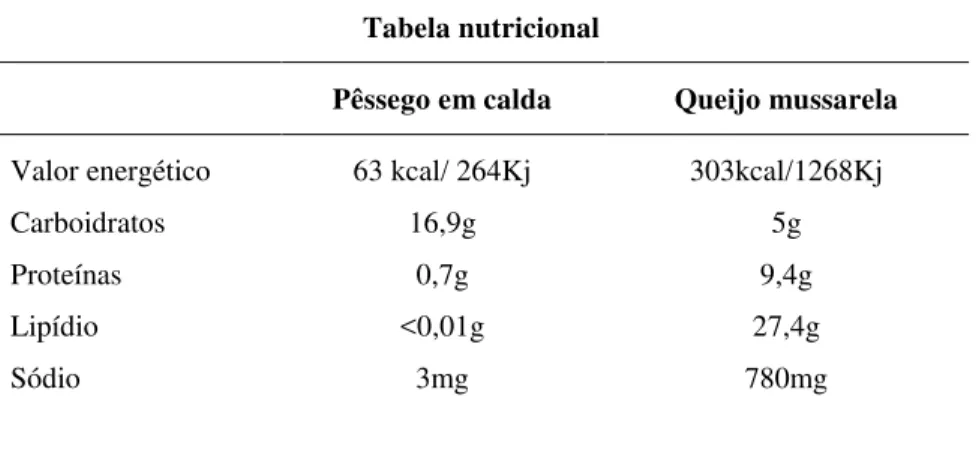
execução do experimento. Consideramos a tabela nutricional dos alimentos escolhidos
(Tabela II), a fim de verificar a quantidade de calorias, carboidratos, proteínas e sódio
presente em cada um deles. Cada alimento oferecido foi cortado em formato de cubo de
aproximadamente 1cm3, e apresentado aos animais em pratos de coloração prata, próprios
para a alimentação. Foram oferecidos três cubos por animal, sendo que para animais que se
encontravam juntos com a família, contabilizamos três cubos para cada indivíduo do grupo
com o objetivo de reduzir a competição pelo alimento nos grupos.
Tabela II – Tabela nutricional dos alimentos selecionados para o experimento, para 100g, conforme a Tabela Brasileira de Composição de Alimentos- TACO, UNICAMP (Lima et al. 2006).
4.4 Procedimento
Iniciamos o experimento com a habituação dos animais ao observador, realizada por
meio de visitas diárias ao cativeiro, no período de um mês, onde se encontravam os animais
previamente selecionados. As visitas de adaptação foram realizadas pela manhã, entre 7h e
10h. Os animais permaneceram em suas próprias gaiolas-viveiro ao longo do período de
Tabela nutricional
Pêssego em calda Queijo mussarela
Valor energético Carboidratos Proteínas Lipídio Sódio
17 estudo. As observações para coleta de dados foram realizadas no período da manhã, entre
7h e 9h30min, horário correspondente ao período em que normalmente recebem
alimentação no cativeiro. As observações foram realizadas em dias consecutivos, cinco
vezes durante a semana durante duas semanas consecutivas.
4.4.1 Fêmea gestante
Cada fêmea foi mantida em sua gaiola-viveiro de origem, sem haver necessidade de
isolamento no período da execução do experimento. No final da gestação, mais
especificamente em torno de duas semanas antes do nascimento dos filhotes, oferecemos
um item alimentar dentre os previamente selecionados, e acompanhamos o comportamento
da fêmea, frente a este alimento. As observações foram baseadas período gestacional da
fêmea, sendo realizadas 15 sessões para cada, terminando juntamente ou próximo da data
do nascimento dos filhotes(+/- 2 dias). O tempo para cada observação foi estabelecido em
15 minutos, ou até o término do alimento oferecido. Cada uma das fêmeas se encontrava
em uma gaiola-viveiro juntamente com o seu respectivo grupo familiar ou parceiro (casais).
O procedimento para as fêmeas fora do período gestacional foi idêntico ao realizado com as
fêmeas grávidas.
4.4.2 Filhotes
Iniciamos as observações com os filhotes assim que estes atingiram a idade de 10
semanas, período referente ao início da ingestão de sólidos e também a parcial
do grupo, para que não houvesse influência destes na aceitação dos alimentos. Com a
finalidade de diminuir o estresse, esta separação foi realizada sem que houvesse o manejo
dos filhotes, utilizando a divisão existente entre os dois compartimentos da própria
gaiola-viveiro. Seguido da separação, aguardávamos um período de aproximadamente cinco
minutos antes da apresentação do alimento aos filhotes, também numa quantidade de três
cubos por indivíduo, e registramos os comportamentos realizados. O período estabelecido
para a observação foi de 15 minutos por experimento, sendo realizadas 10 sessões por par
de filhotes. Apresentamos para os filhotes exclusivamente o item doce. Os filhotes foram
subdivididos em dois grupos, o primeiro cuja mãe teve contato com o alimento durante a
gestação e o segundo cujas mães não tiveram esse contato.
4.5Comportamentos registrados
a. Exploração do alimento: caracterizada pela frequência que o animal tocou,
cheirou, mordeu ou lambeu o alimento.
b. Latência para a primeira mordida no alimento: tempo transcorrido desde o
início da observação à primeira mordida do animal no alimento oferecido.
c. Consumo do alimento: ao final da observação era contabilizado o número
total de itens consumidos completamente pelos animais.
d. Aproximações ao comedouro: quando os filhotes se dirigiam ao comedouro,
sendo consideradas efetivas quando o animal atingia uma distância de
menos de 15 cm do comedouro.
e. Afastamento do comedouro: quando os filhotes saíram do comedouro a uma
4.6 Análise dos dados coletados
Utilizamos o teste não-paramétrico de Mann-Whitney para todas as comparações com
um nível de significância de 5%. As comparações realizadas foram:
(1) Para as fêmeas adultas:
a. Comparamos a quantidade de itens consumidos bem como a latência para o
consumo do primeiro item em função da gestação;
b. Comparamos a quantidade de itens consumidos bem como a latência para o
consumo do primeiro item de acordo com a qualidade gustativa dos
alimentos (doce ou não doce);
c. Comparamos a quantidade de itens consumidos bem como a latência para o
consumo do primeiro item de acordo com a gestação e a qualidade gustativa
dos alimentos (doce ou não doce).
(2) Para os filhotes:
a. Comparamos a quantidade de itens consumidos, a latência para o consumo
do primeiro item, bem como a quantidade de aproximações considerando os
filhotes cujas mães receberam o alimento e cujas mães não receberam o
alimento;
b. Comparamos a quantidade de itens consumidos, a latência para o consumo
do primeiro item, bem como a quantidade de aproximações em função do
21
5. RESULTADOS
5.1 Fêmeas
A influência da gestação no comportamento neofóbico das fêmeas
De uma forma geral, pudemos observar que as fêmeas no período gestacional se
mostraram mais receptivas aos itens alimentares não familiares, independente da qualidade
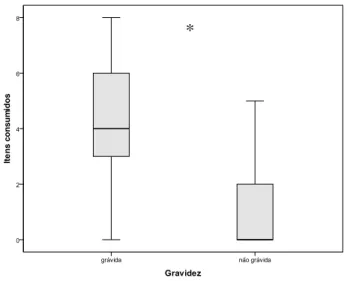
gustativa dos mesmos. Houve diferença significativa tanto no consumo de itens (U = 468,5;
Z = -5,473; P < 0,001) (Figura 1), quanto na latência para o consumo do primeiro item (U =
376,5; Z = -6,145; P < 0,001) (Figura 2).
Figura 1: Mediana e desvio interquartílico da quantidade de itens consumidos em relação à gestação.
Figura 2: Mediana e desvio interquartílico da latência para o consumo do primeiro item, independente da qualidade gustativa, em relação à gestação.
A qualidade gustativa e o comportamento de neofobia em fêmeas de C. jacchus
As fêmeas, independentemente de estarem ou não em período gestacional
responderam melhor aos alimentos doces, apresentando uma latência menor para o
consumo do primeiro item (U = 549,000; Z = -4,653; P < 0,001) (Figura 3), e consumindo
uma maior quantidade de itens doces do que de itens salgados (U = 689,500; Z = -3,665; P
< 0,001) (Figura 4).
23
Figura 3: Mediana e desvio interquartílico da latência para o consumo do primeiro item apresentado em relação à qualidade gustativa deste item (doce ou não doce).
Figura 4: Mediana e desvio interquartílico da quantidade de itens consumidos por fêmeas em relação à qualidade gustativa do alimento.
Gestação x qualidade gustativa
Com o objetivo de verificar se a qualidade gustativa poderia influenciar
diferencialmente fêmeas grávidas e não grávidas, analisamos mais detalhadamente o
comportamento das fêmeas, e foi possível observar que para cada alimento especificamente
(doce ou não doce) a média dos itens consumidos foi maior para as fêmeas no período de
gestação para ambos respectivamente (alimento doce: U = 102,00; Z = -5,246; P < 0,001;
alimento não doce: U = 60,00; Z = -3,939; P < 0,001) (Figura 5), bem como a latência para
o consumo do primeiro item foi menor do que a apresentada pelas fêmeas não grávidas
(alimento doce: U = 108,00; Z = 5,109; P < 0,001; alimento não doce: U = 74,500; Z =
-3,509; P<0,001) (Figura 6).
Figura 5: Média e intervalo de confiança (95%) para os itens consumidos doces e não doces em relação à gestação.
*
25
Figura 6: Média e intervalo de confiança (95%) para a latência em segundos para o consumo do primeiro item.
5.2 Filhotes
A experiência intra-uterina e a neofobia alimentar de filhotes após o nascimento
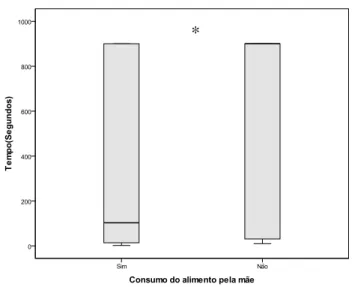
Houve diferença significativa entre os comportamentos referentes à latência para o
consumo do primeiro item (U = 876,000; Z = -2,427; P = 0,015) (Figura 7), quantidade de
itens consumidos (U = 895,500; Z = -2,302; P = 0,021) (Figura 8) e também nas
aproximações em relação ao comedouro (U = 862,000; Z = -2,530; P = 0,011) (Figura 9),
para filhotes cuja mãe tivera contato com o item apresentado em relação aos cuja mãe não
teve este contato. Os comportamentos exploratórios não apresentaram diferenças
significativas.
*
Figura 7: Mediana e desvio interquartílico da latência para o consumo do primeiro item em relação à experiência intra-uterina (consumo do item pela mãe durante o período gestacional).
Figura 8: Mediana e desvio interquartílico da quantidade de itens consumidos por filhotes em relação ao consumo prévio da mãe destes mesmos itens durante o período gestacional.
*
27
Figura 9: Mediana e desvio interquartílico da quantidade de aproximações do filhote em relação ao consumo prévio do item pela mãe durante o período gestacional.
Neofobia alimentar e o sexo dos filhotes
Verificamos que os filhotes do sexo feminino expressam menos comportamentos
neofóbicos do que os do sexo masculino, apresentando uma latência menor para o consumo
do primeiro item apresentado (U = 1346,500; Z = -2,230; P = 0,02) (Figura 10),
consumindo mais itens (U = 895,500; Z = -3,127; P = 0,002) (Figura 11) e se aproximando
Figura 10: Mediana e desvio interquartílico da latência para o consumo do primeiro item relacionado ao sexo do filhote.
Figura 11: Mediana e desvio interquartílico da quantidade de itens consumidos em relação ao sexo do filhote.
*
29
Figura 12: Mediana e desvio interquartílico da quantidade de aproximações do alimento apresentado em relação ao sexo do filhote.
6. DISCUSSÃO
Vários estudos sugerem que animais onívoros, em especial primatas, apresentam
preferência para alimentos doces, não demonstrando neofobia àqueles que contenham alto
teor de açúcar (Johnson 2007, Laska 2000, 2001). Em macaco aranha, por exemplo, Laska
(2000) verificou que quando oferecidos itens alimentares variados, houve preferência aos
de alto valor energético, em especial aos que contenham maior quantidade de carboidratos.
Além disso, outros primatas também parecem utilizar a qualidade gustativa doce como
critério para a seleção de sua dieta (Laska 1999). Nossos resultados parecem apoiar essa
hipótese. A partir das categorias comportamentais observadas, o alimento doce, parece
realmente não despertar tantos comportamentos neofóbicos nas fêmeas quanto o alimento
salgado, sendo mais rapidamente aceito. As fêmeas apresentadas aos itens doces tiveram
um tempo de latência menor para o consumo do primeiro item, e também ingeriram uma
maior quantidade de itens do que as expostas ao item salgado. Esse fato parece estar de
acordo com a teoria de que a qualidade gustativa doce seria uma pista confiável de
palatabilidade e de menores riscos, já que grande parte dos itens que possuem essa
característica podem ser considerados alimentos em potencial. Além disso, a aceitação de
itens não familiares mais energéticos pode ser benéfica ao diminuir a chance do risco de
inanição (Krebs & Davies 1996).
Nossa pesquisa sugere que, além disso, o período de gestação exerce influência no
comportamento de neofobia alimentar, tanto para os itens doces quanto para os itens
salgados. Fêmeas grávidas se mostraram mais receptivas para as duas categorias
alimentares quando comparadas às fora deste período, porém o alimento doce foi mais
31 comportamentos neofóbicos a estes itens, aceitando-os desde a primeira exposição, não
realizando, inclusive, comportamentos exploratórios antes da ingestão e consumindo uma
maior quantidade de itens do que a pré-estabelecida para cada indivíduo. Qual seria o
motivo para esses alimentos despertarem respostas menos neofóbicas em gestantes?
É preciso observar que para fêmeas de C. jacchus, um aporte alimentar adequado é
um fator de grande importância, especialmente na gestação (Box 1997, Lacerda 2004,
Power 1999), pois, assim como a maioria dos primatas do Novo Mundo, o custo energético
envolvido nesse período é extremamente alto (Nievergelt & Martin 1999). Ainda segundo
estes autores, as mudanças que ocorrem no corpo da fêmea grávida (aumento do peso
corporal em aproximadamente 20%) certamente acarretam um aumento no consumo
energético, que deve ser suprido com enriquecimento da dieta. Além disso, em
calitriquídeos, fêmeas normalmente dão luz a gêmeos dizigotos que pesam em torno de
15% do seu próprio peso, e que o tamanho do filhote aumenta ainda mais os gastos
energéticos e parece estar relacionado com o melhoramento da alimentação da fêmea no
período gestacional (Sanches et al. 1999). Devido a todos esses custos, há de se esperar que
fêmeas grávidas sejam pouco neofóbicas a itens que possam representar alimentos em
potencial, especialmente aos mais energéticos.
Em relação aos filhotes, verificamos a maior receptividade dos mesmos aos
alimentos quando estes foram previamente apresentados às suas mães durante o período de
gestação. Esse resultado corrobora com a nossa hipótese de que há influência intra-uterina
no comportamento alimentar pós-natal, ou seja, o consumo de determinados alimentos pela
fêmea grávida influencia diretamente o comportamento alimentar de sua prole. Esse fato
era esperado, pois, durante o seu desenvolvimento o feto é exposto a nutrientes, sabores e
absorvidos pelo feto, entrando em contato com o seu aparato olfato-gustatório. Alguns
estudos mostraram que, em humanos e em animais de laboratório (ratos, camundongos e
suínos), mudanças no comportamento alimentar da mãe durante o período gestacional
podem gerar um “imprinting” pré-natal afetando o apetite durante a vida futura da prole, ou
seja, preferências para sabores específicos podem ter origem na exposição ao sabor da dieta
materna (Stylianos 2008). Outros trabalhos demonstraram o aumento da palatabilidade na
infância e na vida adulta de roedores, após a experiência pré-natal, ao álcool (Chotro et al.
1990, Youngentob et al. 2007, Eade et al. 2009) e ao sal (Leshen 1999).
Poderíamos, dessa forma, considerar que a baixa neofobia aos alimentos doces na
infância e ao longo da vida, podem ter origem intra-uterina, visto que observamos que a
fêmea se mostra altamente neofílica a estes itens quando grávida. Esse fato pode ter
evoluído por ser adaptativamente vantajoso. Se considerarmos que fêmeas que aceitam
facilmente alimentos doces e altamente energéticos influenciam sua prole para a aceitação
de itens alimentares da mesma categoria, esta fêmea estará ampliando a chance de
sobrevivência de seus filhotes e consequentemente atuando no aumento de sua própria
aptidão.
Observamos também que ocorre diferença na resposta aos alimentos quando
comparamos o sexo dos filhotes. Em nossos dados, os filhotes do sexo masculino se
mostraram mais prudentes, apresentando maior latência para o consumo dos itens
oferecidos e também ingerindo uma menor quantidade, demonstrando o que poderíamos
considerar como um comportamento mais neofóbico. Esses dados estão de acordo com a
nossa hipótese, uma vez que diferenças semelhantes já foram observadas em saguis, C.
jacchus, adultos. Michels (1998) ao realizar estudos sobre fêmeas de C. jacchus, observou
33 acordo com Petto e Devin (1988), observações realizadas imediatamente após a introdução
de um alimento na condição de cativeiro mostraram forte diferença entre os sexos tanto na
aceitação quanto no tempo gasto com o consumo. Fêmeas se mostraram mais receptivas e
também permaneceram mais tempo se alimentando, além de ingerirem uma maior
variedade de alimentos, quando estes foram oferecidos. Além disso, estudos realizados por
Yamamoto et al. (2004) também verificaram essa diferença comportamental entre os sexos.
Segundo esses autores, ao apresentar uma tarefa de desempenho para a aquisição de um
alimento, os saguis do sexo feminino (reprodutoras ou não) obtiveram um melhor resultado
que os do sexo masculino, apresentando uma latência menor para a obtenção do alimento,
utilizando mais estratégias para tal, e com isso consumindo mais itens.
Testes análogos foram realizados em calitriquídeos do gênero Saguinus (S. labiatus,
S. fuscicollis e S. oedipus). Apesar dos autores terem encontrado algumas variações entre as
três espécies, os resultados se mantiveram semelhantes na diferença do desempenho e
aquisição de alimento entre os sexos, com as fêmeas apresentando melhores resultados,
menor neofobia ao aparato experimental, obtendo assim uma maior quantidade de
alimentos (Box et al. 1995). É importante ressaltar que os primatas utilizados nesses testes
não apresentam diferenças morfológicas que poderiam justificar a diferença de
desempenho, e sendo assim, esses resultados poderiam também ser explicados pela maior
necessidade energética da fêmea devido aos custos envolvidos posteriormente na
reprodução.
Os comportamentos que atuam na melhoria do desempenho para a obtenção do
alimento, assim como as características menos neofóbicas, podem ter sido evolutivamente
importantes para a fêmea, pois a aptidão da fêmea é altamente influenciada pelo acesso aos
fêmeas receptivas (Silk 2002). Em várias condições o sucesso reprodutivo da fêmea tem
sido ligado a sua condição nutricional, pois fêmeas melhor alimentadas iniciam a
reprodução mais cedo, produzem proles mais saudáveis e se reproduzem por mais tempo
35
7. CONCLUSÕES
Os resultados obtidos em nosso trabalho nos permitem chegar às seguintes
conclusões:
1. Tanto fêmeas em período gestacional quanto as fora desse período são menos
neofóbicas a alimentos com alto teor de açúcar do que a alimentos salgados;
2. Fêmeas em período gestacional são menos neofóbicas do que fêmeas fora deste
período tanto para itens doces quanto para salgados;
3. Filhotes que tiveram contato intra-uterino com um item alimentar respondem
mais prontamente a esse alimento do que os filhotes que não passaram por essa
experiência;
4. Desde a infância as fêmeas já demonstram menos comportamentos neofóbicos
do que os machos de sagui (Callithrix jacchus).
Apesar da consistência dos nossos resultados, mais dados necessitam ser coletados a
fim de esclarecer outros mecanismos envolvidos na neofobia alimentar bem como na
8. REFERÊNCIAS BIBLIOGRÁFICAS
Abbott, D. H, Barrett, J., & George, L. M. (1993). Comparative aspects os the social
suppression of reproduction in female marmosets and tamarins. In: A. B. Rylands (Org.),
Marmosets and Tamarins: Systematics, Behaviour and Ecology (pp. 152-163). Oxford: Oxford Science Publications.
Ahlborn, S., & Rothe, H. (1999). Food selection of Semi free Common Marmosets
(Callithrix jacchus): Indications for Optimal Foraging. Primates, 40(3), 479-486. Box, H. O. (1997). Forranging strategies among male and female marmosets and tamarins
(Callitrichidae): New perpectives in an underexplored area. Folia Primatology, 68, 296-306.
Box, H. O, Röhrhuber, B., & Smith, P. (1995). Female tamarins (Saguinus-Callitrichidae)
feed more successfully than males in unfamiliar foraging tasks. Behavioral Processes, 34, 3-12.
Caldwell, C. A., & Whithen A. (2004). Testing for social learning and imitation in common
marmosets, Callitrhix jacchus, using an artificial fruit. Animal Cognition, 7, 77-85. Carlson, N. R. (2001). Comportamento Alimentar. In: P. J. C. Nogueira (Org.), Fisiologia
do comportamento (pp. 392-423). São Paulo: Manole
Castro, C. S. S. (2003). Tamanho da área de vida e padrão de uso do espaço em grupos de
37 Chambers, P. L., & Hearn, J. P. (1985). Embryonic, foetal and placental development in the
Common marmoset monkey (Callithrix jacchus). Journal of Zoology, 207(4), 545-561. Chotro, M. G., & Molina, J. C. (1990). Acute ethanol contamination of the amniotic fluid
during gestational day 21: postnatal changes in alcohol responsiveness in rats.
Developmental Psychobiology, 6, 535-47.
Day, R. L., Coe, R. L., Kendal, J. R., & Laland, K. N. (2003). Neophilia, innovation and
learning: a study of intergeneric differences in callitrhichid monkeys. Animal Behavior, 65, 559-571.
Dominy, N. J., Lucas, P. W., Osorio, D.,Yamashita, N. (2001). The Sensory Ecology of
Primates Food Perception. Evolutionary Anthropology,10, 171-186.
Eade, A. M., Scheehe, P. R., Molina, J. C., Spear, N. E., Youngentob, L. M.,
&Youngentob, S. L. (2009). The consequence of fetal ethanol exposure and adolescent
odor re-exposure on the response to ethanol odor in adolescent and adult rats.
Behavioural and Brain Function, 5 (3).doi:10.1186/1744-9081-5-3.
Ferrari, S. F. (1987). Food transfer in a wild marmoset group. Folia Primatologica, 48, 203-206.
Ferrari, S. F. (1996). A vida secreta dos saguis. Ciência Hoje, 20(119), 18-26.
Ferrari, S. F., & Lopes Ferrari, M. A. (1989). A re-evaluation of the social organization of
the Callitrichidae, with reference to the ecological differences between genera. Folia Primatologica, 52, 132-147.
Fragaszy, D., Mason, W. A. (1978). Response to novelty in Saimiri and Callicebus:
Fragaszy, D., Visalberghi, E., & Galloway, A. (1997). Infant tuffed capuchin monkeys´
behavior with novel foods: opportunism, not selecitivity. Animal Behavior, 53, 1337-1343.
Galef Jr., B. G. (1996). Food selection: Problems in understanding how choose foods to eat.
Neuroscience Biobehavioral Review, 20(1), 67-73.
Goehl, J. R., & Tardif, S. (2009). Mother´s little helper? The placenta and its role in
intrauterine maternal investment in the common marmoset (Callithrix jacchus). In: S. M.
Ford, L. M. Porter & L. C. Davis (Orgs.), The smallest anthropoids the marmoset/callimico radiation (pp.301-330). New York: Springer
Hauser, M. D. (1988). How infant vervet monkeys learn to recognize starling alarm calls
-The role of experience. Behaviour, 105, 187-201.
Haydon, J. L. (1984). Infant-caregiver Interactions in Common Marmosets (Callithrix
jacchus): Consequences of Rejections and Rub-offs of Infants by Caregivers. Primates, 25(4), 450-461.
Hearn, J. P. (1983). The common marmoset (Callithrix jacchus). In: J. H. Hearn (Org.)
Reproduction in New World Primates: New Models in Medical Science (pp. 183-216). Boston (USA): MTP Press Limited.
Hearn, J. P., Abbott, D. H., Chambers, P. D., Hodges, J. K., & Lunn, S. F. (1978). Use of
39 Johnson, E. C. (2007). Rhesus Macaques (Macaca mulatta) Are Not Neophobic Toward
Novel Food With a High Sugar Content. American Journal of Primatology, 69, 591-596.
Koenig, A., & Rothe, H. (1991). Social Relationships and Individual Contribution to
Cooperative Behaviour in Captive Common Marmosets (Callithrix jacchus). Primates, 32(2), 183-195.
Krebs, J. R., & Davies, N.B (1996). Introdução à Ecologia Comportamental. São Paulo:
Atheneu Editora.
Lacerda, D. F. (2004). Fatores que influenciam a escolha alimentar em sagüis comuns (Callithrix jacchus) cativos. Dissertação de Mestrado. Universidade Federal do Rio Grande do Norte, Natal.
Laska, M. (2001). A comparison of food preferences and nutrient composition in captive
squirrel monkeys, Saimiri sciureus, and pigtail macaques, Macaca nemestrina.
Physiology & Behavior,73, 111-120.
Laska, M., Schull, E., & Scheuber, H. P. (1999). Taste Preference Thresholds for
Food-Associated Sugars in Baboons (Papio hamadryas anubis). International Journal of Primatology, 20, 25-34.
Laska, M., Salazar, L. T. H., & Luna, E. R. (2000). Food Preferences and Nutrient
Composition in Captive Spider Monkeys, Ateles geoffroyi. International Journal of Primatology, 21, 671-683.
Laska, M., Scheuber, H. P., Salazar, L. T. H., & Luna, E. R. (2003). Sour-taste tolerance in
Leshem M., (1998). Salt preference in adolescence is predicted by common prenatal and
infantile mineralofluid loss. Physiology Behaviour, 63, 699-704.
Lima, D. M., Colugnati, F. A. B.,Padovani, R. M., Rodriguez-Amaya, D. B., Salay, E., &
Galeazzi, M. A. M. (2006). Tabela Brasileira de Composição de Alimentos-TACO. Campinas-SP: Núcleo de Estudos e Pesquisas em Alimentação-NEPA.
Luckett, W. P. (1974). Comparative Development and Evolution of the Placenta in
Primates. American Journal of Primatology, 3, 142-234.
Michels, A. M. (1998). Sex Differences in Food Acquisition and Aggression in Captive
Common Marmosets (Callithrix jacchus). Primates, 39(4), 549-556.
Nievergelt, A. M., & Martin, R. D. (1999). Energy intake during the reproduction in
Captive Common Marmosets (Callitrhix jacchus). Physiology & Behavior, 65, 849-854.
Petto, A. J., & Devin, M. (1988). Food Choice in Captive Common Marmosets (Callithrix
jacchus).Laboratory Primates Newsletter, 27, 7-9.
Power, M. L. (1999). Aspects of energy expenditure of callitrichid primate: physiology and
behaviour. In: P. Dolhinow & A. Fuentes (Orgs.) The Nonhuman Primates (pp. 225-230). California: Mayfield Publishing.
41 Rothe, H. (1978) Sub-grouping behaviour in captive Callitrhix jacchus families: A
preliminary investigation. In: H. Rothe, H. J. Wolters & J. P. Hearn (Orgs.), Biology and Behaviour of Marmosets (pp. 233-257). Gottingen (Germany): Eigenverlag. Rutherford, J. N., & Tardif, S. (2009). Developmental Plasticity of the Microscopic
Placental Architecture in Relation to Litter Size Variation in the Common Marmoset
Monkey (Callithrix jacchus). Placenta, 30, 105-110.
Rylands, A. B. (1996). Habitat and evolution of social and reproductive behavior in
Callitrichidae. American Journal of Primatology, 38, 5-18.
Rylands, A. B., & Faria, D. S. (1993). Habitats, feeding ecology, and home range size in
the genus Callithrix. In: A. B. Rylands (Org.) Marmosets and Tamarins: Systematics, Behaviour, and Ecology (pp. 262-272). London: Oxford University Press.
Sanchez, S., Pelaez, F., Burmann, C. G., & Kaumanns, W. (1999). Costs of Infant-Carrying
in the Cotton-Top Tamarin (Saguinus oedipus). American Journal of Primatology, 48,
99–111.
Silk, J. B. (2002). Female, Food, Family and Friendship. Evolutionary Antropology, 11,
85-87.
Stevenson, M. F. (1977). The common marmoset (Callitrhix jacchus) as a model for
ethological research. American Association for Laboratory Animal Science, 27, 895-900.
Stevenson, M. F., & Rylands, A.B. (1988). The marmosets, genus Callithrix. Ecology and Behavior of Neotropical Primates, 2, 131- 222.
Stylianos, N. (2008). Prenatal imprinting of postnatal specific appetites and feeding
behavior.Metabolism Clinical and Experimental, 57(2), 22-26.
Torri, R., Abbott, D. H., & Nigi, H. (1996). Morphological Changes of the Ovary and
Hormonal Changes Through the Ovarian Cycle of the Common Marmoset (Callithrix
jacchus). Primates, 37(1), 49-56.
Visalberghi, E. (1994). Learning processes and feeding behaviour in monkeys. In B. G.
Galef, M. Marnardi & P. Valsecchi (Orgs.), Behavioral Aspects of feeding: basic and applied research on mammals (pp. 257-270). Chur (Switzerland): Harwood Academic Publ.
Visalberghi, E., Sabbatini, G., Stammati, M., & Adessi, E. (2003). Preferences toward
novel food in Cebus apella: the role of nutrients and social influences. Physiology & Behavior, 80, 341-349.
Visalberghi, E., & Fragaszy, D. (1995). The behaviour of capuchin monkeys, Cebus apella,
with novel food: the role of social context. Animal Behavior, 49, 1089-1095.
Visalberghi, E., & Adessi, E.(2006). Comer com Inteligência. Viver Mente e Cérebro,
158, 73-78.
Vitale. A., & Queyras, A. (1997). The response to novel foods in common marmoset
(Callithrix jacchus): the effects of different social contexts. Ethology, 103, 395-403. Voelk B., Scrauf, C., & Huber, L. (2006). Social contact infuences the response of infant
marmosets towards novel food. Animal Behavior, 72, 356-372.
Wiens, F., & Zitzmann (2003). Social Dependence of Infant Slow Lorise to Learn Diet.
43 Wise, R. A. (2009). Role of brain dopamine in food reward and reinforcement.
Philosophical Transactions of the Royal Society B, 361, 1149-1158.
Yamamoto, M. E. (1993). From dependence to sexual maturity: the behavioural ontogeny
of Callitrichidae. In: A. B. Rylands (Org.) Marmosets and Tamarins: Systematics, Behaviour, and Ecology (pp.235-254). London: Oxford University Press.
Yamamoto, M. E., Domeniconi, C., & Box, H. O. (2004). Sex differences in common
Marmosets (Callitrhix jacchus) in response to an unfamiliar food task. Primates, 45, 249-254.
Yamamoto, M. E., & Lopes, F. A. (2004). Effect of Removal from the Family Group on
Feeding Behavior by Captive Callithrix jacchus. International Journal of Primatology, 25(2), 489-500.
Yamamoto, M. E., Arruda, M. F., Alencar, A. I., Sousa, M. B. C., & Araújo, A. (2009).
Matting System and Female-Female Competition in the Common Marmoset, Callithrix
jacchus In: S. M. Ford, L. M. Porter & L. C. Davis (Orgs.) The smallest anthropoids the marmoset/callimico radiation (119-134). New York: Springer
Youngentob, S. L., Molina, J. C., Spear, N. E., & Youngentob, L. M. (2007) The effect of
gestational ethanol exposure on voluntary ethanol intake in early postnatal and adult rats.