Acta bot. bras. 1(2):27-40 (1988) sup ••
ASPECTOS DA EVOLUC;AO E DA GEOGRAFIA DO GENERO PHILODENDRON SCHOTT (ARACEAE)
Simon J. Mayo (1)
RESUMO - , 0 genero Philodendron se divide em tres subgeneros, cada urn com distintos padr6es de morfologia, anatomia e distribui<;lio. Em analise filogen6tica, 0 subgenero Meconostigma, 0 uni-co uni-com centro de especia<;lio no sudeste do Brasil, mostra-se bastante apom6rfiuni-co, ainda que cladis-ticamente prirnitivo no genero. AnAlises fen6ticas mostram que 0 genero se constitue urn taxon dis-tinto, apesar de nlio ter urn s6 carater defmitivo. Mostram tarnbtSm que feneticamente, Philodendron esta mais perto de certos generos da Africa ocide(ltal (Cuicasia, Cercestis, Rhektophyllum) do que dos generos principalmente asiaticos com os quais esta ligado pela classifica<;lio tradicional.A morfologia do gineceu varia muito no subgenero MeconostigmLl, com as formas mais simples ocorrendo no sudeste do Brasil e as mais elaboradas na Amazonia. Compara<;lio com 0 "grupo de fora" indica que as formas mais simples sao primitivas no subgenero, apontanto as esp6cies princi-palmente rupfcolas, P. adamantinum Schott e P. leal-costae Mayo & G.M.Barroso, como as de gi-neceus mais primitivos. Com base na morfologia do gineceu, poderia ser sugerido que 0 subgenero evoluiu em princfpio na parte oriental do Brasil, como urn grupo adaptado aos habitats abertos, e que na bacia amazonica ele chegou somente mais tarde. Os dois outros subgeneros, Philodendron e ·Pteromischum, surgiram posteriormente, principalmente como epffitas de florestas umidas, e se mostram hoje mais diversos no nordeste da Am6rica do SuI.
ABSTRACT - The genus Philodendron can be divided into three subgenera which are very dis-tinct in vegetative and floral morphology, floral anatomy and distribution. Phylogenetic and phe-netic analysis of floral characters indicate that subgenus Meconostigma, which unlike subgenera Philodendron and Pteromischum has a predominantly southeastern range in South America, is hi-ghly apomorphic but cladistically primitive in the genus. Phenetic analyses show that the genus can be viewed as distinct, although no definitive diagnostic character is known to exist. These analyses also show that Philodendron is phenetically closer to certain west African genera (Cuicasia, Cerces-tis, Rhektophyllum) than to the mainly Asiatic genera with it has been traditionally associated. Gy-noecial morphology in sub gen. Meconostigma varies considerably, with simpler forms predomina-ting in southeastern Brazil, and the most elaborate in Amazonia. Outgroup comparison suggests that the simpler types are more primitive and indicates that the rupicolous species P. adamantinum Schott and P.leal-costae Mayo & G.M.Barroso possess the most primitive gynoecial type. Based on gynoecial morphology it is suggested that the subgenus evolved initially in eastern Brazil as a group adapted to open habitats, arriving only later in the Amazon basin. The other two subgenera, Philo-dendron add Pteromischum, arose later, as epiphytes of humid forestes and today are most diverse in the northeast of South America.
Key-words: Philodendron, Araceae, biogeography, phylogenetic systematics
Introdu~ao
o
genero Philodendron Schott, bem conhecido dos paisagistas pelo grande numerode especies ornamentais, e 0 segundo maior da famRia Araceae, com cerca de 350
espe-cies hoje reconhecidas (Bunting 1986, Mayo 1986). Sua area de distribuic;:ao e essencial-mente neotropical, atingindo seu limite meridional na regiao do Rio de la Plata. As especies sao particularmente bem diversificadas nas matas Omidas tropicais, mas tambem ocorrem em brejos, sobre afloramentos rochosos e ate em regioes semiaridas.
Um conhecimento detalhado da geografia e da taxonomia do genero e interessante aos fitoge6grafos devido ao grande numero de especies e a sua diversidade (veja por exemplo, os resultados de French & Tomlinson 1984). Porem, os conhecimentos taxon6-micos ao nivel especifico de que dispomos atualmente permanecem ainda superficiais. Novas especies tern sido descobertas continuamente a tax as que nao tem decres cido (Bunting 1975, 1986). Baseado Elm dados eXistentes, 0 genero pode sel'dividido em tres subgeneros, Philodendron, Pter;'mischum
e
MeconostigfTUl, que sao bem distintos em mor-fologia floral e vegetativa, anatomia floral e distribuic;:ao.Um estudo taxonomico recente de Philodendron subgen. MeconostigfTUl, 0 menor dos tres subgeneros (Mayo 1986), forneceu novos dados com certos pontos de interesse ao fitoge6grafo, ainda que especulativos. 0 presente trabalho consta de um esboc;:o dos resultados relevantes e do contexto taxonomico.
Material e Metodos
Na preparac;:ao da revisao de Philodendron subgen. MeconostigfTUl, material herbori-zado dos seguintes herbflrios foi utiliherbori-zado; A, B, C, CAY, CTES, E, F, G, HB, HBG, INPA, K, L, LE, M, MBM, MICH, MO, NY, P, RB, S, SEL, SPF, U, UB, UC, UEC, US. Para 0 es-tudo da morfologia e da anatomia floral do genero, foi utilizado material vivo e preservado em ~rcool, colhido ou no campo no Brasil ou das colec;:6es cultivadas do Roberto Burle Marx
e
do Royal Botanic Gardens, Kew . .A anatomia floral foi estudada ou atraves da dissecc;:ao de flores inteiras fixadas, uti-lizando um microsc6pio Olympus (modelo SZ-III, binocular), ou atraves de sec;:6es em se-rie de flores impregnadas com a cera "Paraplast", e cortadas corn um microtomio do tipo Jung Model 1130. T odas as sec;:6es foram coloridas utilizando safranina '0' e "alcian blue", e montadas em "euparal"; sao todas depositadas nas colec;:6es do Royal Botanic Gardens, Kew.
Resultados e Discussao
1. Definic;:ao de Philodendron subgenero MeconostigfTUl
. Este taxon sempre foi admitido pelos botanicos que estudaram 0 genero, des de 0 reconhecimento do pr6prio genero (Schott 1829). Os carAteres diagn6sticos, de acordo com a tradic;:ao, incluem os. seguintes: 1) hAbito arborescente, 2) caule duro e fibroso co-berto das cicatrizes de folhas velhas dispostas em espirais, e, 3) estames finos e
compri-dos.
o
subgenero MeconostigfTUl e tambem distinto geograficamente, tendo sua maior di-versidade nas regi6es sui e leste da regiao tropical da America do Sui. Na bacia amaz6ni-ca 0 numero de especies e bem menor e, nas vertentes pacificas dos Andes , na parte se-tentrional da Colombia e nas Anti/has, 0 subgenero e desconhecido (Figs . 1-16). Ern con-traste marcante, os subgeneros Philodendr on e Pteromischum atingem sua maior diversi-dade na regiao noroeste da America do Sui, com especies que alcanc;:am 0 Mexico e as' Antilhas.Meu estudo(Mayo 1986) confirmou que 0 subgenero tem limites bem defi nidos e ampliou 0 nurnero de caracteres que 0 diferenciam dos dois outros subgeneros, Philoden-dron e Pteromischum. De acordo com osnovos dados, 0 su bgenero M econostigfTUl pode ser definido pelos seguintes carAteres:
I) Zona esteril do espAdice (composta de estamin6dios) longa, entre zonas pistiladas e estaminiferas.
Evolu~ao em PIriIoduttInM 29
III) Caule adulto composto morfologicamente de unidades simpodiais de uma folha s6, e de um padrao caracterizado pela supressao do intern6 hipopodial.
IV) Vascularizagao do gineceu mostrando um sistema axilar de feixes 0 qual e
indepen-dente do sistema que fornece os 6vulos atraves dos funiculos. V) Feixe vascular do estame nao bifurcado.
VI) Dutos de resina na zona adaxial da espata tendo a forma de "J" em segao longitudinal.
2. Posigao cladistica de Philodendron subgen. Meconostigma
A base das ideias filogeneticas apresentadas neste trabalho vem de um estudo
anatomico dos carateres da inflorescencia em 15 especies do genero Philodendron,
repre-sentando a maioria das segoes atuais -(Tabela 1). Das especies estudadas, quatro
perten-cem ao subgenero Meconostigma e onze ao antigo subgenero "Euphilodendron"
reconhe-cido por Krause (1913).
TABELA 1
Especies do genero PhiLodendron estudadas ao nfvel anatomico (para resultados
comple-tos veja Mayo 1986).
Subgenero Meconostigma
P. bipinnatifidum Schott ex Endlicher P. goeLdii G.M.Barroso
P. saxicoLum K.Krause P. williamsii J.D. Hooker
Subgenero Pteromischum
P. sonderianum Schott
Subgenero Philodendron
Segao Baursia
P. insigne Schott P. LongiLaminatum Schott P.frits-wentii Bunting
Segao Calostigma
P. smithii Engler
P. burle-marxii G.M.Barroso
Segao Tritomophyllum
P. tripartitum (Jacquin) Schott
Segao Polytomium
P.fendleri Engler
Segao Schiztplzyllum
P. pedalum (W.J.Hooker) G.Don
Segao Macrolonchium '
P. melinonii Brongniart Segao desconhecida
P. "sp. nov. do Equador"
As Segoes Philodendron e Camptogynium, ambas do subgenero Philodendron, nao foram
Para a analise cladistica das quinze especies, utilizaram-se 41 caracteres anatomi-cos da inflorescencia e, como "grupo de fora", a maio ria dos generos tradicionalmente
as-sociados com Philodendron da subfamOia Philodendroideae (Engler 1912), junto com
Cer-cestis, Rhektophyllum, Nephthytis e Montrichardia da subfamOiaLasioideae, e Culcasia da
sul;JfamOia Pothoideae (Mayo 1986). 0 cladograma resultante forneceu dados que refort;:am
o reconhecimento dos tres subgeneras Meconostigma, Pteromischum e Philodendron.
o
subgenero Meconostigma tem um numera relativamente grande de caracteresau-tapom6rficos e trata-se, portanto, de um taxon bem distinto. Po rem, cladisticamente, 0
sub-genera e 0 mais primitiv~ dos tres, de acordo com esta analise. Os subgeneras
Pteromis-chum e Philodendron sao grupos-irmaos, e juntos formam 0 grupo-irmao do subgenera Me-conostigma (Fig. 17).
o
cladograma e baseado exclusivamente em caracteres da anatomia floral, e assimpode ser testado par comparat;:ao com outras carateres provavelmente derivados (apo-m6rficos) destes subgeneras. A distribuit;:ao destes carateres se mostra congruente com
o cladogratna. Assim, 0 subgenera Meconostigma, delimitado no cladograma par carateres
anatomicos, pode tambem ser caracterizado pela longa zona esteril do espadice. 0
sub-genera Pteromischum e tambem caracterizado pela morfologia do caule (composto de
uni-dades simpodiais de muitas folhas ou 'plei6fi1as'), com os carateres associados: - lamina
foliar simples e bainhas foliares compridas. 0 subgeneras Philodendron pode sertambem
caracterizado pela morfologia do caule, que
e
composto de unidades simpodiais de umaunica folha, mas nas quais 0 intern6 hipopodial e sempre bem desenvolvido enquanto que
o 'primeira intern6 da unidade e suprimido.
Esses carateres, quando associados aos do cladograma original, forneceram os
elementos para uma classificat;:ao (Fig. 17) que permite supor a seguinte hip6tese para a
hist6ria do genera. Do Philodendron ancestral,o subgenera MecOlwstigma foi 0 primeira a
surgir.O subgenera Pteromischum resultou de uma adaptat;:ao estreita ao habito epifitico na
mata umida, ligada
a
evolut;:ao de uma morfologia do caule muito espcializada. Muitosca-racteres florais, porem, permaneceram plesiom6rficos (generalizados) e parecidos aos dos generas do "grupo de fora".
Subgenera Philode1lllron desenvolveu uma morfologia caulinar caracteristica e um
sistema excepcional de dutos resiniferas no espadice. A secret;:ao de resina na
inflores-cencia durante antese ocorre em quase todas as espl!cies estudadas, mas
e
excretada doespadice apenas em subgenera Philodendron (Mayo 1986). Subgenera Philodendron
e
0mais diversificado em numera de especies, abrangendo a maioria das especies do pr6prio genero.
3. Morfologia do gineceu no subgenero Mec01wstigma
o gineceu do subgenera
Meconostigma mostra ampla variat;:ao na estrutura doesti-II:,e e no numera de 16culos do ovario. Esta variat;:ao estrutural do estilete parece ser
resultante de crescimento diferencial em tres zonas diferentes durante ontogenese:
-I) extensao dos 16bulos estilares.
II) extensao da regiao do estilete na qual correm os canais estilares.
III) estensao, ou rebratat;:ao, do apice primario floral e desenvolvimento do mesmo em cupula central estilar.
Estes tres tipos de crescimento, ocorrendo quase simultanearnente, resultam em di-versas formas maduras (Figs. 18-29). A comparat;:ao com aqueles generas do "grupo de
Evolu~iio em P,.,."",_ 31
que a ocorrencia de crescimento-extensao em cada uma das regioes trata-se de um
ca-rater apom6rfico (derivado). Duas especies, P. adamantinum (Fig. 18) e P.leal-costae (Fig.
19), se assemelham muito morfologicamente dos generos do "grupo de fora".
o
numero de 16culos do ovario, de 3 ate 4716culos em cada ovario, e tambem muitovariavel. A comparac;ao com os generos estudados no "grupo de fora" indica que ovarios
3-loculados e 4-loculados sao pr'ovavelmente plesiom6rficos no genero Philodendron, e 7
especies mostram estas condic;oes em pelo menos alguns dos gineceus num s6 espadice (P. dardanianum (Fig. 20), P. brasiliense (nao ilustrada), P. tweedianum (Fig. 21), P.
uligino-sum (Fig. 22), P. adamantinum (Fig. 18), P. corcovadense (Fig. 24), P.leal-costae (Fig. 19) ).
4. A relac;ao entre geografia, hAbitos e morfologia do gineceu no subgenero Meconostif?ma
A ecologia das especies sugere que 0 subgenero Meconostigma e bem adaptado a
ambientes abertos com alto grau de insolac;ao. Porem, no ambiente fiorestal este subgene-ro tem side relativamente menos bem sucedido.
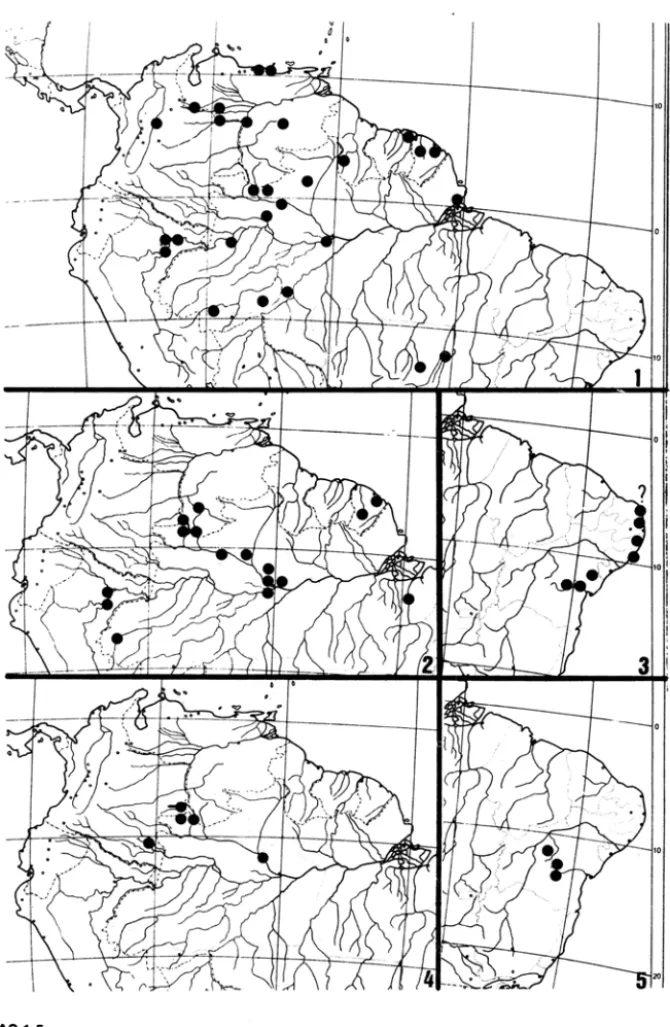
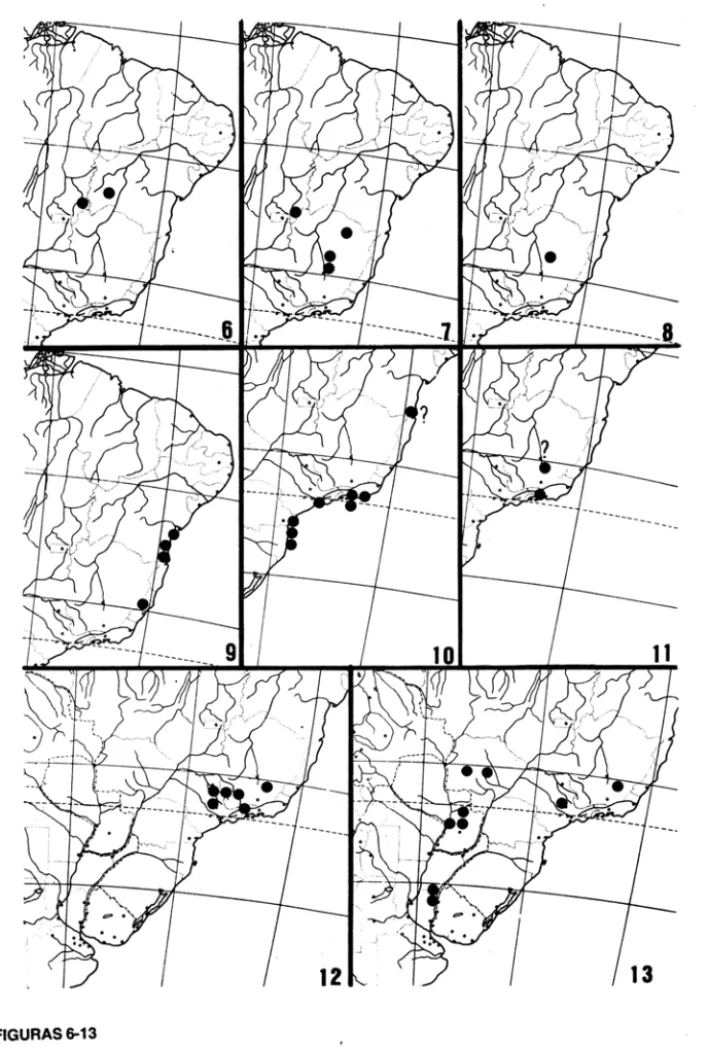
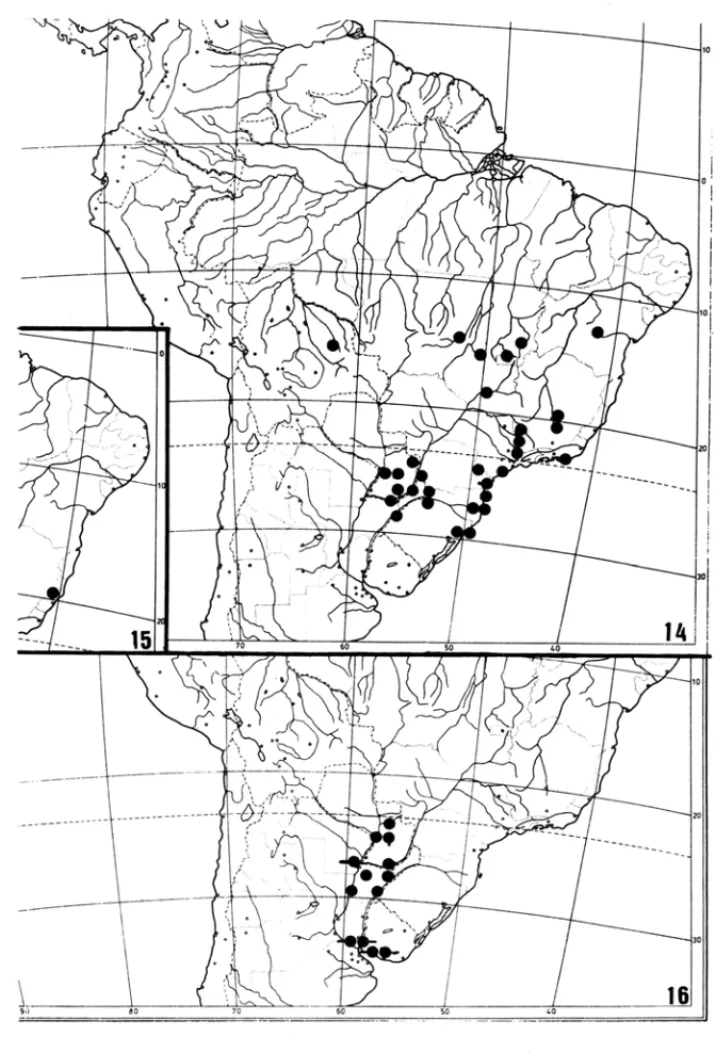
Todas as especies com gineceu exibindo numero elevado de caracteres plesiom6rfi-cos ocorrem no leste e no sui da area de distribuic;ao do subgenero (Grupo A, Tabela 2, Figs. 3, 6-8, 10-11, 14, 16) e mostram mais variac;ao ecol6gica ainda que todas sejam de ambientes abertos. Sao, portanto, plantas aquaticas, saxicolas, epiffticas ou terrestres so-bre substrato arenoso, como, por exemplo, a restinga. As duas especies com a morfologia
mais primitiva, P. leal-costae (Fig. 3) e P. adamantinum (Fig. 6), sao saxfcolas, ainda que P.
leal-costae ocorra tamMm em restingas no estado de Alagoas (Fonseca-Vaz et al. 1984).
Em contraste, as tres especies que ocorrem na regiao amazonica, P. goeldii. P.
ve-nezuelense e P. solimoesense (Grupo C, Tabela 2, Figs. 1-2,4) tem gineceus mais deriva-dos, com estiletes muito elaborados e geralmente mais de 20 16culos em cada ova.rio. Sao todas especies epifiticas em mata fechada e plantas terrestres em ambientes mais aber-tos.
Outras especies com morfologia estilar apom6rfica ocorrem no leste e no sui da area de distribuic;ao (Grupo B, Tabela 2, Figs. 5,9,12-13,14) e mostram variaC;ao de ha.bito
pa-recida com a das especies do Grupo A, com plantas aquaticas (P. brasiliense. Fig. 12; P.
undulatum. Fig. 13), saxfcolas (P. saxicolum. Fig. 5), e epifiticas (P. williamsii. Fig. 9). As
diversas formas de P. bipinnatifidum (Fig. 14), que tem gineceus de carater apom6rfico, se
enquadram neste grupo de especies.
5. Padr6es hist6ricos e geograficos do subgenero Mecollostigma
Na bacia amazOnica, a ocorrencia de poucas especies de habito mais unifornie e
com morfologiado gineceu muito derivada sugere que 0 subgenero Mecollostigma poderia
ter colonizado essa regiio numa ~poca relativamente r~cente em contraste com a parte
sudeste da area de disbibu~ao, ollde as espeCies sao mais numerosas, mais
diversifica-das em Mbito e mostram, pelo menos em alguns casos, rnorfoiogia do gineceu primitiva. Por outro lado, podeser suposto que nas condic;:6es climaticas atuais, a bacia ama-zonica oferece urn conjunto de ambientes ecol6gicos muito mais reslrito a esse grupo de
plantas heli6filas, e que apenas umas Poucas especies especializadas sobreviveram
a
ampliac;:ao da area de fIoresta umida desde 0 Pleistoceno, quando 0 clima daquela regiao
era mais srido. e talvez mais favoravel ao florescimento de urn numero maior de especies.
Porem, e evidente que as matas da regiao atlantica oriental do Brasil tem especies
que sao epitrticas mas tambOO1 mostram gineceus generalizados e que, dentro da regiao
amaz6nica, M um grande conjunto de areas de habitat mais aberto, como por exemplo as
serras guianenses e imensas areas brejosas, onde especies analog as As do Brasil oriental
TABELA2
Philodendron subgenero Meconostigma - Habito em relac;:ao a morfologia do gineceu
Especie Habitb
A. Es~cies do sudeste .com gineceu relativamente primitivo
P. adamantinum P. leal-costae P. dardanianum P. tweedumum
saxfcola saxfcola, terrestre
aquAtica aquAtica
P . uliginosum aquAtica
P . speciosum eprtita,saxfcola P. corcovadense epffita, terrestre P. bipinnatifidum
*
aquAtica, saxfcola, epffitaB. Esp~cieS do sudeste com gineceu mais especializado
P. brasiliense P. undulatum P. saxicolum _
P. williamsii
aquAtica aquAtica
saxfcola
eprtita, terrestre P. bipinnatifidum *. aquAtica, saxfcola, eprtita
C. Esp~cies amazonicas com gineceu muito derivado
P , goeldii epffita, terrestre P. venezuelense 'eprtita, terrestre P. solinwesense eprtita, terrestre
*
P. bipinnatifidume
muito variAvel em morfologia floral e vegetativa (veja Mayo 1986).Alem dis'so, a distribuic;:ao das especies amazonicase muito mais extensado que a da maio ria das especies orientais, entre as quais ha exemplps de especies estre,itamente
endemicas (P. adamantinum, P. saxicolum, P. speciosum). Estes fatos sugerem que tais
especies orientais sejam muito antigas, enquanto que asespecies amazonicas saode ori~
gem mais recente. . ' / .
6. Evoluc;ao e geografia dO genero'PhilodeiUiron
. As dua's hip6teses tefe'rentes aci subgenero Meconostigma, ' um1i posi<;M dadfstica
primitiva e .lIma area de 'djstribuic;:ao relictual nOffrasil oriental, d~9ap oioa poSsibilidade de
que 0 grupo ~ irrriao do . generb consta dos tres _. ge~er'osafricairo:ocidentaisCeri:estis,
Rhektophjillum e Clllcasia.' Os estLidos de French (com. pess.f
e'
French"& Tomlinson(1984)
ja.
indicaram queessetcjU;3.trogeneros sao bem' mais pr6xlmos do que antes'peri-s,avam ostaxbnomislas. Aevoluc;:Mdo conjunto de gener-os deveter ocorridcina
superfl-cie. da
Gondwana-
ori~
nta i ;eO
$ut:igene
r 6
Mec(J/lostigma poderia ter 'aparecidb no prinCipioda hist6rici do continel'lte da America do Sui briental, talvez entao se desenvolvendocomo
urn gruPb de especies adaptadas a ambientes abertos. . - . . .
_ Entre as especies dos' subgenero $ Pteromischum e Philodendron; porem, a maioria
sao eprtitase poucas sM' adaptadas asituac;:6esexpostas: Epffitas destessubgeneros sao tfpicas de ambientes sombreados abaixo da Copada floresta, sobre troncos ou galhos maiores. Estes dois subgeneros devem ter evoluido .essencialmente como plantas de fler
Evolu~ao em p~
33
As areas de distribuic;:ao dos subg€meros Pteromischum e Philodendron sugererr:
uma hist6ria condicionada principalmente pela alterac;:ao continua do mosaico de areas de fiorestas umida inferidas pelo modelo dos "refugios" (Prance 1982). Os centros principais de diversidade especifica correspondem, pelo menos parcialmente, a alguns dos refugios indicados para a America do SuI. Possivelmente como reac;:ao ao desenvolvimento de
re-fugios fiorestais, bem como
a
diversificac;:ao ecol6gica provocad~pela orogenia dos Andes,o sLibgenero Philodendron passou por um processo intense de especiac;:ao, especialmente
no Peru, Equador, Colombia, Panama e Venezuela.
7. Rela<;:6es taxonomicas do genero Philodendron
Engler (1912) ligou Philodendron com dois generos principalmente aSiaticos, HOIlu/
-/OmRno e S c hismllto~lottis , e esse conceito foi mantido por Borger e Nicolson (1987). 0 estudo de Grayum (1984) sobre as afinidades genericas resultou em concius6es bastante
diferentes. Ele sugere que Philodendron e mais pr6ximo a Dieffenbllchia, um genero
neo-tropical.
Uma analise fenetica de 14 generos (Figs. 30-31), utilizando 40 caracteres principal-mente da infiorescencia (veja Mayo 1986 para uma apresentac;:ao mais detalhada), mostrou
que:-I) os tres subgeneros de Philodendron agrupam-se mais entre si do que a qualquer
outro genero (Fig. 31) ;
II) 0 genero feneticamente mais proximo a Philodendron e Cercestis, da Africa
oriental.
A analise mostrou tambem que Philodendron e sistematicamente bastante isolado e
que nao tem relac;:ao estreita com nenhum genero analisado (Fig. 30-31). Esta conclusao, ainda que inconclusiva, indica que nao ha argumento firme para manter a ligac;: ao
tradicio-nal do genero com HOIrUl!ome na e S c hisl/l (/ IO ~ ! o tti s , mas ao mesmo tempo, apoia a
hip6te-!3e, ja discutida, de 0 grupo Cercestis·Rhektopln'III11Il-Cu!casill e seu "qrupo-irmao".
8. Geografia dos generos da subtamfila f' 11/ /odcilllroidclIc
A pro pos ta relac;:ao afri cana de Philodcl/{!ron tem poucas semelhanc;:as na famnia
das Ar aceae de acordo com ideias atu ais da taxonomia. Mayo (1987) e Grayum (1984) discutem as IigaC;:6es transatlEmticas da fam nia, das quais as mais bem defin idas parecem
se r uma en tre Callopsis (Africa orien tal) e Ulell/'ll/ll (regiao a m a z o n~c a do Peru ), e outra
entre Typhonodorlllll (Madagascar) e Pelt(//ulra (regiao oriental dos EUA).
Uma class ificac;:ao melho r fundamen tada e necessari a antes de que uma analise ri -go ro sa da biogeografia dos generos da subfamnia seja posslvel. Uma analise cladlstica
abrangendo todos os generos e 0 primeiro requisito . Apesa r dis so, dos dados disponlveis
pode ser lanc;:ada a hip6tese que 0 gru po-irmao de Philodendro n e provavelmente 0
con-junto de Cercestis-Rhektophyllllll1-Cll/msill, e que 0 taxon ancestral imediato deles ocorri a
na area continental hoje represe ntada pe la Africa ocidental e a parte oriental da Ameri ca do
Sui. Esta ultima regiao seria a area de distribui c;:ao inicial do genero Philodcnd ron , e a
Agradecimentos
Agradego a Diretoria do XXXVIiI Congresso Nacional de Bot~nica, realizado na Uni-versidade de Sao Paulo, em janeiro 1987, e especialmente a Presidente, Nanuza Lufza de Menezes e a Primeira Secretaria, Ana Maria Giulietti, pela oportunidade de participar do Congresso, por muita amizade e colaboragao. Agradego tamb{)m ao Professor Ver-non H. Heywood, a quem devo apoio essencial durante a realizagao do estudo em que foi baseado 0 presente trabalho; ao Roberto Burle Marx por ter me permitido estudar a sua colegao maravilhosa de Araceas; aos assessores pela revisao do portugues; e aos cura-dores dos seguintes herbarios por ter me fornecido material: A, B, C, CAY, CTES, E, F, G,
HB, HBG, INPA, L, LE, M, MBM, MICH, MO, NY, P, RB, S, SEL, SPF, U, UB, UC, UEC, US.
Referencias Bibliograficas
BOGNER, J. & NICOLSON D.H. 1987. A revised classification of Araceae with dichotomous keys. In J.Arditti & L.Nyman, The Biology of the Araceae (no prelo).
BUNTING, G.S. 1975. Nuevas especies para la revision de las Araceas venezolanas. Acta Bot. Venezuel. 10: 263-335.
BUNTING, G.S. 1986. New taxa of Venezuelan Araceae. Phytologia 60(5): 293-344.
ENGLER, A. 1912. Araceae-Philodendroideae-Philodendreae. Allgemeiner Teil, Homolameninae und Schjsmatoglottidinae. Em A. Engler (ed.), Das Pflanzenreich 55 (IV. 23Da): 1-134.
FONSECA-VAZ, A.M.S ., Andreata R.H.P. & Guedes P.R. 1984. Philodendron leal-costae Mayo & G.M.Barroso (Araceae) - Observa<;6es s6bre sua distribui<;ao geografica, morfologia e ecologia. Anais XXXIV Congr .Nac.Bot., Porto Alegre, Brasil 2: 229-233.
FRENCH, J.C. & TOMLINSON P.B. 1984. Patterns ofstem vasculature in Philodendron. Amer. J .Bot. 71 : 1432-1443.
GRAYUM, M.H . 1984. Palynology and Phylogeny of the Araceae. Ph. D dissertation, Univ. Massachusetts (Amherst), EUA.
KRAUSE, K. 1913. Araceae-Philodendroideae-Philodendreae-Philodendrinae. ln A. Engler (ed.), Das Pflanzeweich60 (lV.23Db): 1-143.
MA YO, SJ. 1986. Systematics of PItiIodendroIt Achott (Araceae) with special reference to inflorescence choracters, Ph.D. thesis, University of Reading, UK.
MA YO, S.l. 1987. Aspects of Aroid Geography (manuscrito).
PRANCE, G.T. 1982 (ed.). Biological Diversification in the Tropics. Columbia Univ. Press, New York . SCHOTT, H.W. 1829. Fur Liebhaber der Botanik. Wiener Zietschr. Kunst, Literatur, Theater WId Mode
FIGURAS 1-5
Areas de distr ib ui~ao em Philodendron subgen. Meconostigma.
Fig. 1. P. solinwesense A.C.Smith; Fig. 2. P. goeldii G.M.Barroso; Fig. 3. P. leal-costae Mayo & G.M.Barroso; Fig. 4. P. venezuelense Bunting; Fig. 5. P. saxicolwn K.Krause;
FIGURAS 6-13
Areas de distribui<;§o em Philodendron subgen. Meconostigma.
Evolu~ao em PhiIothtttlrrm 37
FIGURAS 14·16
Areas de distribuigao em Philodendron subgen. Meconostigma.
MECONOSTIGMA
PTEROMISCHUM
PHILODENDRON
1
10
13
2
11
11.
3
r12
1.)
.
I.
16
5
r
17
6
7
1B
.
B
··19
9
LU
21
22
23
17
FIGURA. 17
Classificac;:lio dos tr~s subgeneros de Philodendron (asigla' indica um carater apom6rfico)
Caracteres
1. Centro de diversificac;:ao especllica no sudeste do Brasil 2. *Canais estilares independentes no gineceu
3. oZona estaril do espadice Iongo.
4. *Sistema vascular do gineceu com feixes axilares independentes 5. 'Estames finos e compridos
6. *Feixe vascular do estame nao bifurcado
7. 'Dutos de resina da zona adaxial da espata com fonna de "J" 8. *Espata muito espessa
9. *lntem6dio hipopodial da unidade simpodial do caule suprimido 10. *Tanino"faltando no estame e no gineceu
11. 'Rafides perifaricas no estilete
12. *Unidades simpediais do caule com muitas folhas ("plei6filas") 13. 'Sulcos est6miais do estame ausentes
14. 'Tecas do estame contfguas 15. *Resina excretada no espadice 16. *Plexo vascular do estilete faltando
17. 'Primeiro intern6dio da unidade simpodial do caule suprimido 18. Centro de diversificac;:ao especffica no noroeste da America do Sui 19. *L6bulos estilares faltan do
20. *Tanino faltando na epidenne do estame 21. 'Constric;:ao central da espata bem evidente 22. 'Canais estilares independentes no gineceu
-~
Evolu~io em P".;j . . .
39
='-=~29
FIGURAS 18-29
Morfologia do gineceu em especies de Philodendron subgenero Meconostigma , em se<;:ao longitudinal. Linhas obllquas indicam tecido do estigma; Meas pontilhadas indicam a presenc;;a de rAfides; linhas espes-sas indicam a posiC;ao dos canais estilares independentes.
I
.J'
~10
"Vi~
1 1
1
'00
PH
RH CE CU
NE
AIJ
ZA
PE
TV MO
OJ
AN
HO
SC
30
1
so
I
1
, '
-1 -1
I
.-L
.--L
,--L
'"
'00ME
PT
PH
RH CE
CU
NE AG ZA
PE
TV MO
or
AN
HO
SC
31
FIGURAS 30-31
Fenogramas dos generos da subfamnia Philodendroideae, resultante s de uma analise tipo "Average Linka-ge Cluster Analysis", utilizando 0 programa chamado "GENSTAT".
30. Matriz 1 - Philodendron tratqdo como um s6 genero.
31. Matriz 2 - Cada subgenero de Philodendron tratado independentemente.