O gên ero Odontesthes perten cen te à fam ília Ath erin opsidae e con h ecido popularm en te com o peixerei, é relativam en te abun -dan te em rios, lagoas e região costeira da Am érica do Sul, desde o sul do Brasil até a Argen tin a (WHITE 1985).
No su l do Brasil foram registradas dez (10) espécies das q u ais o it o (8) est ão em lago as co st eiras, arro io s e rio s: O. bonariensis (Valen cien n es, 1835), O.aff.perugiae Everm an n & Ken dall, 1906, O. hum ensis de Bu en , 1953, O.retropinnis (d e Bu en , 1 9 5 3 ), O. m irin en sis Bem ven u t i, 1 9 9 5 , O. piqu ava
Malabarba & Dyer, 2002, O.bicudo Malabarba & Dyer, 2002,O. ledae Malabarba & Dyer, 2002. En quan to O.incisa (Jen yn s, 1942) é en con trada som en te n o m ar (BEMVENUTI 2002, MALABARBA & DYER 2002), e O.argentinensis (Valen cien n es, 1835) en con tra-se n a região estu arin a e n a região m arin h a costeira.
Odontesthesargentinensis foi descrito através de caracteres pou co precisos, sen do redescrito por BEMVENUTI (1993), através de n ovos caracteres m orfom étricos e m erísticos, in clu in do u m a revisão taxon ôm ica com sin on ím ias atu alizadas.
(Ather
(Ather
(Ather
(Ather
(Atherinopsidae) da r
inopsidae) da r
inopsidae) da r
inopsidae) da r
inopsidae) da reeeeegião mar
gião mar
gião mar
gião mar
gião marinha costeir
inha costeir
inha costeir
inha costeir
inha costeira do sul do Br
a do sul do Br
a do sul do Br
a do sul do Br
a do sul do Brasil
asil
asil
asil
asil
Alex M oresco & M arlise de A. Bemvenuti
Departamento de Oceanografia, Fundação Universidade Federal do Rio Grande. Caixa Postal 474, Avenida Itália, km 8, Campus Carreiros, 96201-900 Rio Grande, Rio Grande do Sul, Brasil.
ABSTRACT. ReprReproductivReprReprReproductivoductivoductivoductiveeeee biologybiologybiologybiologybiology ofof silvofofof silvsilvsilvsilvererererersidesideside OdontesthessidesideOdontesthesOdontesthesOdontesthesOdontesthes argentinensisargentinensisargentinensisargentinensisargentinensis (V(V(V(V(Valenciennes)alenciennes) (Atheralenciennes)alenciennes)alenciennes)(Ather(Ather(Ather(Atherinopsidae)inopsidae)inopsidae)inopsidae)inopsidae) ofofofofof coastal
coastalcoastal coastal
coastal seaseasea rrrrreeeeegionseasea giongiongiongion ofofofofof thethethe souththethesouthsouthsouth ofsouthofofof BrofBrBrBrazil.Brazil.azil.azil.azil. The silverside Odontesthes argentinensis (Valenciennes, 1835) is distrib-uted in the coastal region of the South Atlantic since the south of Brazil until Argentina. This species presents two populations, a population resident in the Patos Lagoon estuary and another one in the coastal sea region. This work had as objective to determine the reproductive biology of the populations of coastal sea region. The sex ratio indicated predominance of females. The size of first maturation (L50) was estimated in 13.8 cm LT for the males and 16.1 cm LT for the females and the length where all were apt to spawn (L100) was estimated in 16.3 and 19.3cm LT for males and females, respectively. The gonadossomatic relation indicated reproductive period be-tween ends of August and beginning of December, period where the temperature of the water is below 20ºC. The factors of total condition (KT) and somatic (KS) had presented great variations during the cycle. A fall in the values of K coincided with the beginning of the spawning, establishing a relation this fall of K represented the energy that was directed for the development of the gonads. In the analysis of the diameter of the oocytes evidence was found allowing state that the spawning don’t occur at once. The total fecundity (TF) and the lot fecundity (LF) are proportional to the total length. The average of oocytes was of 10,014 for FT and 3,651 for FL. KEY WORDS. Fish; reproduction; spawning.
RESUMO. O peixe-rei Odontesthes argentinensis (Valenciennes, 1835) distribui-se na região costeira do Atlântico Sul desde o sul do Brasil até a Argentina. Esta espécie apresenta duas populações, uma população residente no estuário da Lagoa dos Patos e outra na região marinha costeira. Este trabalho teve como objetivo determinar a biologia reprodutiva da população marinha costeira. A proporção sexual indicou predomínio de fêmeas. O tamanho de primeira maturação (L50) foi estimado em 13,8 cm CT para os machos e 16,1 cm CT para as fêmeas e o comprimento em que todos estão aptos para desovar (L100), foi estimado em 16,3 e 19,3 cm CT para machos e fêmeas, respectivamente. A relação gonadossomática indicou período reprodutivo entre final de agosto e início dezembro, período em que a temperatura da água se manteve abaixo dos 20ºC. Os fatores de condição total (KT) e somático (KS) apresentaram grandes variações ao longo do ciclo. Houve uma queda nos valores de K coincidindo com o início da desova estabelecendo uma relação com este período. Na análise do diâmetro dos ovócitos há evidências de três lotes caracterizando desova parcelada. A fecundidade total (FT) e por lote (FL) são proporcionais ao comprimento total. A média de ovócitos foi de 10.014 para FT e 3.651 para FL.
A esp écie p ossu i gran d e p lasticid ad e fen otíp ica o q u e perm ite su a adaptação ao estu ário da Lagoa dos Patos e região m arin h a costeira (BAMBER & HENDERSON 1988), form an do du as popu lações distin tas u m a n o in terior da Lagoa dos Patos e ou -tra n a zon a in ter-m areal da p raia do Cassin o (BEHEREGARAY & LEVY 2000).
BEMVENUTI (1987), an alisan do a popu lação estu arin a, ob-servou q u e a desova ocorre em en seadas rasas e protegidas do estu ário, n a presen ça de vegetação su bm ersa, desde o fin al da estação de in vern o até a prim avera. O local e o período da desova da popu lação m arin h a n ão são con h ecidos. Segu n do in -form ações de pescadores ela pode ocorrer n o período de verão n a zon a de arreben tação.
Dian te da au sên cia de in form ações sobre a reprodu ção desta espécie n o am bien te m arin h o costeiro e por tratar se de u m a espécie com im portân cia econ ôm ica para a região, o pro-pósito deste trabalh o foi determ in ar a biologia reprodu tiva de O.argentinensis da região m arin h a costeira adjacen te à praia do Cassin o, Rio Gran de do Su l. A fim de con tribu ir para con h eci-m en to dos eci-m ecan iseci-m os qu e en volveeci-m a perpetu ação e eci-m an ejo da espécie.
MATERIAL E MÉTODOS
Os exem plares foram coletados n a região m arin h a cos-teira ao su l do m olh e oeste da Barra do Rio Gran de (Praia do Cassin o), p róxim o a Estação Marin h a de Aq u acu ltu ra (EMA-FURG), em 32º12’S e 52º10’W, n o su l do Brasil.
As coletas foram efetu adas m en salm en te du ran te o perí-odo de ju n h o de 2004 a ju n h o de 2005, u tilizan do rede de ar-rasto de praia. O m aterial em laboratório foi fixado em form ol 10% e em seguida, arm azen ados em álcool 70%. Para cada exem -plar foram tom adas m edidas do com prim en to total (CT), peso total (PT), peso das gôn adas (Pg).
Para a an álise do ciclo reprodu tivo foram determ in adas: as proporções sexu ais, o tam an h o de prim eira m atu ração sexu -al (L50), a relação gon adossom ática (RGS), o fator de con dição alom étrico, distribu ição de freqü ên cia do diâm etro dos ovócitos para gôn adas m adu ras, fecu n didade total, fecu n didade por lote e estim ativa do n ú m ero de lotes elim in ados por fêm ea a cada ciclo rep ro d u t ivo . O s p arâm et ro s segu em as in d icaçõ es d e VAZZOLER (1996).
As p rop orções sexu ais foram calcu lad as p or classes d e tam an h o e para o período todo, sen do aplicado o teste do Qu i-q u adrado para verificar se ocorre diferen ça sign ificativa.
O tam an h o de prim eira m atu ração sexu al (L50) foi esti-m ad o calcu lan d o-se a p rop orção d e in d ivíd u os sexu alesti-m en te m adu ros por classes de com prim en to m édio.
O s exem p lares foram classificad os em joven s q u an d o ap resen taram gôn ad as im atu ras, e em ad u ltos q u an d o ap re-sen taram q u alq u er dere-sen volvim en to n as gôn adas. Foram se-parados por sexos e agru pados em classes de 20 m m de com prim en to total. Os valores da freq ü ên cia relativa foraprim represen -tados por classes de com prim en to total. A cu rva obtida foi aju
s-t ad a segu n d o a exp ressão m as-t em ás-t ica:
Fr
= -
1
(
e
-aCTb)
on de: Fr = freq ü ên cia relativa de in divídu os adu ltos; e = base dos logaritm os n atu rais; CT = com prim en to total em cm ; a e b = con stan tes estim adas relacion adas ao aju ste de cu rvas. Para esta an álise foi u tilizado m aterial biológico com plem en tar ce-dido pelo PELD – estu ário da Lagoa dos Patos e região costeira adjacen te (Laboratório de Ictiologia).
O período reprodu tivo foi determ in ado através da rela-ção gon ad ossom ática (RGS), u tilizan d o-se ap en as as fêm eas, con form e VAZZOLER (1996). Os pesos das gôn adas foram obtidos com balan ça de precisão de q u atro casas decim ais.
O fator de con dição alom étrico total (Kt) e som ático (Ks) foram calcu lados para cada exem plar fêm ea segu n do LE CREN (1951). O delta K foi obtido pela su btração Kt – Ks.
A distribu ição de freq ü ên cia do diâm etro dos ovócitos foi feito em u m a su b-am ostra de cada gôn ada com au xílio de lu pa com ocu lar m icrom étrica e posteriorm en te con vertido em m ilím etros para obten ção da freq ü ên cia de ocorrên cia dos diâ-m etros dos ovócitos por diâ-m ês.
A fecun didade (F) foi estim ada pelo m étodo gravim étrico. Foram selecion adas 12 gôn adas e colocadas em solução de Gilson m odificada (VAZZOLER 1996) para dissociar os ovócitos. Feita a dissociação foi registrado o peso total dos ovócitos e extraída sub-am ostras (três réplicas) que foram pesadas, sen do con tado o n úm ero de ovócitos gran des e pequen os de cada réplica.
A fecu n didade total (FT) foi estim ada por regra de três sim ples a partir da m édia do n ú m ero de ovócitos das três répli-cas e m édia do peso das m esm as pelo peso total dos ovócitos n a gôn ada. A fecu n didade por lote (FL) tam bém foi estim ada por regra de três sim ples, u tilizan do o lote de ovócitos de m ai-or diâm etro, bem diferen tes dos dem ais, elim in an do o proble-m a de deliproble-m itar a proble-m oda a ser con siderada. Taproble-m béproble-m foi estiproble-m a-do o n ú m ero de lotes elim in aa-dos para o períoa-do reprodu tivo através da relação FT/ FL (MORSE 1980).
Obten ção de dados abióticos, tem peratu ra e salin idade d a águ a, fo ram o bt id o s p elo Pro gram a Eco ló gico d e Lo n ga Du ração (PELD – estu ário da Lagoa dos Patos e região costeira adjacen te). Para cada m ês foi calcu lado o valor m édio da tem -peratu ra e salin idade e com parado com valores m édios da rela-ção gon adossom ática.
RESULTADOS
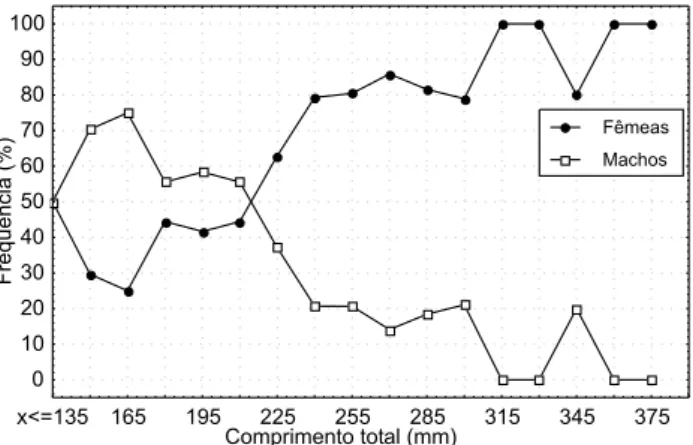
Na an álise da proporção sexu al por classe de com prim en -to -total, foram observados q u e dos 391 exem plares coletados 76,5% foram fêm eas e 23,5% foram m ach os com valor de 2 =
Tam an h o de prim eira m aturação (L50) foi estim ado em 161 m m de CT para as fêm eas e 138 m m CT para m ach os. O com pri-m en to copri-m o qual todos os in divíduos estiverapri-m aptos a partici-par ativam en te do processo reprodutivo (L100) foi estim ado em 193 m m para fêm eas e 168 m m para m ach os (Figs 2 e 3).
As retas de aju stes do L50 ficaram assim defin idas:
(
)
Fr
= -
1
e
-3 22 10, ´ -14´CT11 04, para as fêm eas e(
)
Fr
= -
1
e
-2 9 10, ´ -12´CT10 05, para os m ach os.As m aiores m édias de RGS foram registradas en tre agosto e d ezem bro, corresp on d en d o aos m eses d e rep rod u ção, com um pico m áxim o em n ovem bro, on de o peso m édio das gôn adas eq u ivaleu a 7,3% do peso total da fêm ea (Fig. 4). Nos m eses de m arço a ju n h o a RGS foi in ferior a u m (1), pois as gôn adas se en con travam em repou so.
Na figura 5 estão represen tadas as variações de tem peratu-ra, salin idade e RGS. O período de baixa tem peratura da água correspon de à estação de in vern o, com a m en or m édia registrada de 13,5 °C n o m ês de agosto de 2004. Nos m eses seguin tes h ouve progressivo aum en to de tem peratura até atin gir 26,1°C n o m ês de m arço de 2005, correspon den te à estação de verão. A salin idade apresen tou um a queda brusca n o m ês de outubro de 2004 atin -gin do 22,8‰ . Nos dem ais m eses os valores m édios variaram en tre um m ín im o de 27‰ (julh o de 2005) e um m áxim o de 33‰ (agosto de 2004). O aum en to n os valores de RGS correspon deu ao in ício do aum en to da tem peratura. Os m aiores registros de RGS, que in dicam o período de m aior desova, ocorreu quan do a tem peratura estava abaixo de 20ºC, período que correspon de ao fin al do in vern o e aos m eses da prim avera. Os m aiores valores de RGS tam bém correspon deram ao período de queda brusca de salin idade, n o en tan to esta queda é ocasion al, devido ao gran de volum e de ch uvas que ocorreu n a época. En tre os m eses de jan ei-ro e fevereiei-ro n ão h ouve captura de exem plares.
O fator de con dição in dica o bem estar da espécie para o período reprodu tivo. Na figu ra 6 estão represen tados os valo-res m édios do fator de con dição total (Kt) e os valovalo-res m édios do fator de con dição som ático (Ks). Os valores apresen taram gran des variações m en sais, sen do de ju n h o a dezem bro de 2004 co n d izen t es co m as variaçõ es d o RGS, in d ican d o a en ergia despen dida do corpo para as gôn adas e tam bém a in flu ên cia do peso das gôn adas m adu ras. De m arço a ju n h o de 2005 as gôn adas n ão estão m ais in terferin do n o peso total e as varia-ções de k pode ser resu ltado de con divaria-ções alim en tares recen tes. O delta K defin ido pela diferen ça en tre os dois fatores de con dição Kt e Ks (Fig. 7), represen ta o fator gon adal on de os valores m ais altos correspon deram ao período de desova. Os m aiores valores foram registrad os en tre agosto e n ovem bro, p eríod o q u e coin cid iu com a d esova observad a p ela RGS. A qu eda n o valor de delta K, do m ês de n ovem bro para dezem -bro, m ostrou q u e a m aioria das fêm eas en con trava-se desova-das, con firm an do o fin al do período de desova.
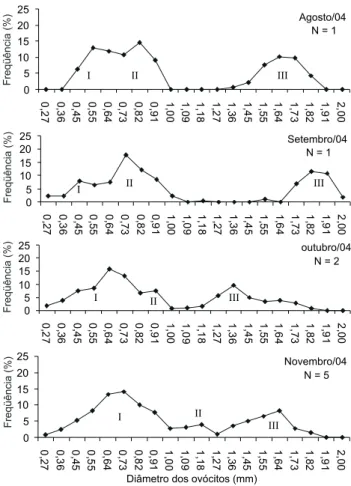
A distribu ição de freq ü ên cia do diâm etro dos ovócitos m ostrou três grupos m odais an alisados em n ove exem plares dos quatro m eses de m aior RGS (Fig. 8). A m oda III form ada por um grupo de ovócitos bem distin tos e m aduros, m ediu en tre 1,36 e 1,91 m m de diâm etro em agosto e 1,64 a 2,0 m m em setem bro, quan do ocorreu a desova deste lote. Em outubro um n ovo lote se destacou com 1,18 a 1,82 m m de diâm etro, passan do a m edir 1,27 a 1,91 m m n o in ício de n ovem bro. As outras duas m odas I e II são form adas por ovócitos de diâm etros m en ores e, em bora n ão apresen tem gran des diferen ças em freqüên cia, são distin tos n a estrutura. No grupo da m oda I en con tram -se os ovócitos de reserva com form a e superfície irregulares e cor esbran quiçada. Os ovócitos da m oda II apresen tam superfície arredon dada, m ais lisa e cor am arelada. A desova é parcelada em lotes, eviden ciada pela presen ça das três m odas e pelo aum en to n o diâm etro dos ovócitos da últim a m oda.
Os doze exem plares utilizados para estim ar a fecun didade m ediram en tre 212 a 342 m m CT, com fecun didade total varian -do de 6082 e 26724 ovócitos, respectivam en te. A fecun didade por lote foi estim ada para o grupo de ovócitos de m aior diâm e-tro. O n úm ero de ovócitos por lote variou de 2282 a 8694, respec-tivam en te, para os m esm os exem plares citados acim a. O n úm ero de lotes variou de 2,05 a 3,63 com m édia arredon dada para 3 lotes elim in ados por fêm ea duran te um ciclo reprodutivo (Tab. I). A relação en tre a fecu n didade total e o com prim en to to-tal resu ltou em u m a cu rva de potên cia (Fig. 9). A relação en tre a fecu n didade por lote e o com prim en to total apresen tou u m a relação lin ear (Fig. 10). As expressões ficaram assim defin idas: Fecu n didade total FT = 0,05 x CT3,685 e Fecu n didade por lote:
FL = 57,381 x CT – 11799.
DISCUSSÃO
Ho u ve u m gran d e p red o m ín io d e fêm eas n o p erío d o com o u m todo e n as classes de m aior tam an h o. A m aioria dos exem plares m ach os predom in ou n as m en ores classes de com
-Comprimento total (mm)
Freqüência
(%)
0 10 20 30 40 50 60 70 80 90 100
x<=135 165 195 225 255 285 315 345 375
Fêmeas
Machos
Figura 1. Proporção sexual da população m arinha costeira de O.
argent inensis, relação entre com prim ento total e freqüência de
Mean+SD Mean-SD
Mean+SE Mean-SE
Mean
Meses 0
1 2 3 4 5 6 7 8 9 10 11 12
J/04 J A S O N D J F M A M J/05
RGS
RGS
TEM
SAL
Meses 0
3 6 9 12 15 18 21 24 27 30 33 36
J/04 J A S O N D J F M A M J/05
V
ariáveis
Figuras 2-7. (2-3) Distribuição das freqüências relativas de fêm eas (2) e m achos (3) adultos de O.argent inensis da população marinha costeira, por classes de com prim ento total (cm ). (4) M édias m ensais da relação gonadossom ática para exem plares fêm eas de O.
argent inensis, coletadas na região m arinha costeira no período de junho/ 2004 a junho/ 2005. (5) Relação entre as variações m ensais dos
fatores am bientais e biológicos na região m arinha costeira no período de junho/ 2004 a junho/ 2005; (TEM ) tem peratura em graus Celsius, (SAL) salinidade, (RGS) m édia da relação gonadossom ática. (6) Variação m ensal dos valores m édios do fator de condição alom étrico total (Kt) e som ático (Ks) da população m arinha costeira de O.argent inensis, no período de junho/ 2004 a junho/ 2005. (7) Variação m ensal de delta k da população m arinha costeira de O.argent inensis, no período de junho/ 2004 a junho/ 2005.
2
3
4
5
7
0 10 20 30 40 50 60 70 80 90 100
113 123 133 143 153 163 173 183 193 203
Comprimento total (mm)
Freqüência
(%)
L50
L100
Freqüência
(%)
0 10 20 30 40 50 60 70 80 90 100
108 118 128 138 148 158 168 178
Comprimento total (mm)
L100
L50
Delta K
Meses 0,00
0,02 0,04 0,06 0,08 0,10 0,12 0,14
J/04 J A S O N D J F M A M J/05
V
alores
de
Kt
e
K
s
Kt
Ks
Fator de condição
V
alores
de
Kt
e
Ks
Meses 1,40
1,45 1,50 1,55 1,60 1,65 1,70 1,75 1,80
J/04 J A S O N D J F M A M J/05
prim en to total. Segun do NIKOLSKY (1963), GROSS & SARGENT (1985) e LOWE-MCCONNELL (1999), o predom ín io de fêm eas n as classes de m aiores tam an h o é de ocorrên cia gen eralizada en tre os pei-xes, sen do relacion ado com o u m au m en to da fecu n didade. O in verso ocorre, geralm en te, em espécies on de o m ach o defen -de o n in h o con tra predadores.
A proporção sexual de O.argentinensis foi em m édia de três (3) fêm eas para cada m ach o. Segun do VAZZOLER (1996) e WU etal. (2001) esta proporção pode diferir de 1:1 devido às diferen -ças n a taxa de crescim en to, taxa de m ortalidade, lon gevidade, in versão d e sexos e m igrações p ara local d e d esova. Em O. argentinensis as fêm eas foram predom in an tes n as m aiores clas-ses de tam an h o, supon do que a taxa de crescim en to das fêm eas é m aior que a dos m ach os, e assim , estar ocorren do um a seleção predativa qu e pode alterar a proporção sexu al dim in u in do o n ú m ero de m ach os. Ou tros au tores com o CIECHOMSKI (1972), BURBIDGEet al. (1974) e MORENO etal. (1977) tam bém en con tra-ram predom ín io de fêm eas ao estudarem outras espécies de pei-xe-rei, sen do a proporção sexual variável para cada espécie.
O tam an h o de prim eira m atu ração estim ado foi bem in -ferior ao en con trado p or BEMVENUTI (1987) para esta m esm a es-p écie. A au tora estim ou o valor de L50 em 214 m m CT p ara m ach os e 232 m m CT para fêm eas, en qu an to qu e n este traba-lh o o L50 foi estim ado em 138 e 161 m m CT para m ach os e fêm eas, respectivam en te. PAIVA & SCHEFFER (1982) estim aram o L50 em 250 m m para am bos os sexos n a espécie de peixe-rei de águ a doce O.bonariensis n a Bacia do rio Uru gu ai.
Um a diferen ça de tam an h o tam bém é observada em re-lação ao com prim en to em qu e todos os exem plares estão m a-d u ros (L100). Segu n d o BARBIERI etal. (2004), essa p recocid ad e reprodu tiva é u m a característica adaptativa con siderada com o u m a tática reprodu tiva adotada pelas espécies em geral, n o sen -tido de recu perar o eq u ilíbrio popu lacion al. As m an ifestações
Figura 8. Distribuição da freqüência dos ovócitos da população m arinha costeira de O.argentinensis, em quatro meses de maior valor da relação gonadossom ática; I a III são as m odas de ovócitos de cada gônada.
Tabela I. Núm ero estim ado de ovócitos em doze fêm eas de O. argentinensis da população marinha costeira durante o período de desova. Pequeno = núm ero de ovócitos pequenos referentes ao diâmetro da m oda I e II; Grande = número de ovócitos grandes referentes ao diâmetro da m oda III da figura 8.
M eses Com prim ento total (mm ) Número de ovócitos Total ovócito Lotes
Pequeno Grande
Agosto/ 2004 254 4764 2501 7265 2,90
Setem bro/ 2004 278 9590 3637 13227 3,64
Outubro/ 2004 255 5430 3498 8928 2,55
255 5321 2696 8017 2,97
Novembro/ 2004 342 18030 8694 26724 3,07
285 4887 3932 8819 2,24
281 6634 4043 10677 2,64
226 3800 2282 6082 2,67
265 5368 2375 7743 3,26
262 4658 3006 7664 2,55
271 4679 4132 8811 2,13
257 3193 3014 6207 2,06
M édia 269 6363 3651 10014 2,72
Agosto/04 N = 1
0 5 10 15 20 25 0,
27 0,3
6 0, 4 5 0, 5 5 0, 6 4 0, 7 3 0, 8 2 0, 9 1 1, 0 0 1, 0 9 1, 1 8 1, 2 7 1, 3 6 1, 4 5 1, 5 5 1, 6 4 1, 7 3 1, 8 2 1,
91 2,00
I II III
Setembro/04 N = 1
0 5 10 15 20 25 0, 2 7 0, 3 6 0, 4 5 0, 5 5 0, 6 4 0, 7 3 0, 8 2 0, 9 1 1, 0 0 1, 0 9 1, 1 8 1, 2 7 1, 3 6 1, 4 5 1, 5 5 1, 6 4 1, 7 3 1, 8 2 1, 9 1 2, 0 0
I II III
outubro/04 N = 2
0 5 10 15 20 25 0, 2 7 0, 3 6 0, 4 5 0, 5 5 0,
64 0,7
3
0,
8
2
0,
91 1,00 1,09 1,1
8
1,
27 1,36 1,4
5
1,
5
5
1,
64 1,73 1,8
2 1, 9 1 2, 0 0
I II III
Novembro/04 N = 5
0 5 10 15 20 25 0, 2 7 0, 3 6 0, 4 5 0,
55 0,64 0,7
3
0,
8
2
0,
91 1,00 1,0
9 1, 1 8 1, 2 7 1, 3 6 1, 4 5 1, 5 5 1, 6 4 1, 7 3 1, 8 2 1,
91 2,00
Diâmetro dos ovócitos (mm)
d e táticas rep rod u tivas em p eixes são p rovocad as p or vários fat o res co m o as m u d an ças n as co n d içõ es ab ió t icas e p o r sobrepesca (LOWE-MCCONNELL 1999). Estes fatores tam bém atu -am n esta popu lação em estu do.
O ciclo an u al de desen volvim en to gon adal dos exem pla-res fêm eas de O.argentinensis, foi caracterizado com os valores m édios m en sais da relação gon adossom ática (RGS). Segu n do ISAAC-NAHUM & VAZZOLER (1987), este in dicador é bastan te efici-en te para determ in ar o grau de desefici-en volvim efici-en to e o período rep rod u tivo.
O período reprodu tivo desta popu lação foi en tre agosto e dezem bro de 2004, en q u an to para ou tras espécies de ateri-n ídeos, com o Atherinellabrasiliensis (Qu oy & Gaim ard, 1825), a desova ocorreu en tre ou tu bro e fevereiro, tan to n o estu ário do rio Tram an daí (BERVÍAN & FONTOURA 1997), q u an to n o estu á-rio da Lagoa dos Patos (BEMVENUTI 1987). Esta au tora tam bém determ in ou o período de desova en tre agosto a fin al de setem -bro p ara O.argentinensis n o in terior do estu ário da Lagoa dos Patos. O período reprodu tivo dos peixes-rei é bastan te variável, sen do in flu en ciado pelos fatores am bien tais, prin cipalm en -te, pela tem peratu ra de cada região.
Quan to à in fluên cia dos fatores abióticos n o desen volvim en to das gôn adas, h á uvolvim a relação en tre o in ício do crescivolvim en -to dos ovários com o in ício da elevação da tem peratura da água (fin al da estação de in vern o). Quan to à salin idade n ão h á evi-d ên cia evi-d e in flu ên cia n a RGS. A q u eevi-d a evi-d o valo r m éevi-d io evi-d a salin idade, registrada n os m eses de m aior valor da RGS, é um fato ocasion al, decorren te da quan tidade de ch uva n o m ês, que aum en tou a vazão de água doce da Lagoa dos Patos para a região m arin h a costeira. Quan to à ausên cia de exem plares, n os m eses de jan eiro e fevereiro, os pescadores artesan ais argum en tam que a espécie se aproxim a da praia em busca de alim en to tran sporta-do pelos córregos de água sporta-doce que deságuam n o m ar. Quan sporta-do ocorre estiagem esses córregos secam e com o con seqüên cia a espécie n ão se aproxim a destes locais im possibilitan do a captu-ra dos exem plares.
O fator de con dição alom étrico (K) é u m a m edida q u e forn ece in form ações con fiáveis do bem -estar fisiológico da espé-cie (BRAGA 1986) e seu valor reflete con dições alim en tares re-cen tes e gastos de reservas em atividades cíclicas (VAZZOLER 1996). A diferen ça en tre o fator de con dição alom étrico total e
so m át ico , d elt a K, reflet e a ép o ca e a d u ração d o p erío d o reprodu tivo. O delta K tam bém represen ta a en ergia can aliza-da para o desen volvim en to aliza-das gôn aaliza-das, ou gasta em m igrações reprodu tivas, q u e estava acu m u lada n as vísceras, m ú scu -los e/ ou fígado. O delta K apresen tou m aiores valores en tre os m eses de ju lh o e dezem bro, estan do coeren te com os valores apresen tados pela relação gon adossom ática, con firm an do u m lon go período reprodu tivo e in dican do q u e n estes m eses a es-pécie teve m aior desova. Há u m a boa relação en tre os valores do delta K com a RGS, sen do possível con clu ir q u e a RGS é u m a boa in dicadora do período de desova para a espécie.
A distribu ição da freq ü ên cia do diâm etro dos ovócitos foi feita em n ove gôn adas m adu ras, m ostran do três lotes de o vó cit o s, sen d o igu al ao n ú m ero d e lo t es en co n t rad o em Leuresthestenuis (Ayres, 1860) e Basilichthysaustralis Eigen m an n , 1928 (CLARK 1925, MORENOetal. 1977). BEMVENUTI (1987) descre-veu três diferen tes tam an h os de ovócitos para O.argentinensis e X enom elaniris(=Atherinella)brasiliensis n o estu ário da Lagoa dos Patos, caracterizan do u m a desova do tipo parcelada. A de-so va p arcelad a é caract eríst ica em esp écies t ro p icais e su b-tropicais possibilitan do m aior probabilidade de sobrevivên cia (NIKOLSKI 1963). É tam bém u m a estratégia adotada pela espécie q u e m in im iza a com petição in tra-específica e dim in u i o risco de predação ou , ain da, de ocorrer em época on de as con dições am bien tais n ão são favoráveis para sobrevivên cia da prole. Além disso, a desova parcelada é u m m ecan ism o qu e au m en ta o es-forço rep rodu tivo (BURTetal. 1988).
A fecu n didade de O.argentinensis variou de 6082 a 26724 ovócitos, sen d o d iretam en te p rop orcion al ao com p rim en to total. Qu an to m aior o tam an h o da fêm ea, m aior foi o n ú m ero de ovócitos, u m a característica q u e é observada n a m aioria das espécies de teleósteos. BEMVENUTI (1987) estim ou u m a variação da fecu n didade de 1182 a 10783 para O.argentinensis e de 148 a 794 para Atherinellabrasiliensis, peixe-rei de peq u en o tam a-n h o, a-n o estu ário da Lagoa dos Patos.
No en tan to, a estim ativa da fecun didade em peixes com desova parcelada, feita com base em an álises m acroscópicas, tor-n a-se com plexa devido à dificuldade de se distitor-n guir ovócitos de reserva daqueles ovócitos em desen volvim en to (BAGENAL 1978). Por isso, além da fecun didade total foi estim ada a fecun didade p or lote e o n ú m ero d e lotes d e cad a fêm ea p ara u m ciclo 0
5000 10000 15000 20000 25000 30000
200 220 240 260 280 300 320 340 360
Comprimento total (mm)
Número
ovócitos
Fec = 0,0499CT3,685
R = 0,81992
Número
ovócitos
Fec = 573,81CT - 11799
R = 0,8605
20 2000 4000 6000 8000 10000
200 220 240 260 280 300 320 340 360
Comprimento total (mm)
Figuras 9-10. Fecundidade total (9) e por lote (10) da população m arinha costeira de O.argent inensis pelo com prim ento total.
reprodutivo. A fecun didade por lote em relação ao com prim en -to -total apresen -tou um a reta lin ear crescen te, com variação de 2282 a 8694 ovócitos por lote. Em m édia, O. argentinensis da região m arin h a costeira apresen tou três lotes de ovócitos por período reprodutivo, que varia de agosto a dezem bro.
AGRADECIMENTOS
A João P. Vieira e Clarice B. Fialh o, à eq u ipe do Laborató-rio de Ictiologia, aos ocean ólogos Mateu s T. Ku tter e Fábio F. Roselet; à Bióloga Th aís M. Lopes; à Capes.
REFERÊNCIAS BIBLIOGRÁFICAS
BAGENAL, T.B. 1978. Aspects of fish fecu n dity, p. 75-101. In: S.D. GERKIN G (Ed .). Ec o l o g y o f fre sh w ate rfish p ro d u c ti o n .
Oxford, Blackwell, Scien tific Pu blication s, 520p .
BAMBER R.N. & P.A. HENDERSON. 1988. Pré- adaptative plasticity in ath erin id s an d th e estu arin e seat of teleost evolu tion .
Jo urn alFish Bio lo gy, Lon don , 3 3 (Su p p l. A): 17-23.
BARBIERI, G.; F.A. SALLES.; M.A. CESTAROLLI & A.R. TEIXEIRA FILHO. 2004. Estratégias reprodutivas do dourado, Salm inus m axillosus e do curim batá, Prochiloduslineatus, n o Rio Mogi Guaçu, São Paulo, com ên fase n os parâm etros m atem áticos da din âm ica populacion al. ActaScien tiarum, Marin gá, 26 (2): 169-174. BERHEGARAY, L. & J.A. LEVY. 2000. Population gen etics of th e
silver-side Odon testh es argen tin en sis (Ath erin iform es): eviden ce for speciation in a estuary of south ern Brazil. Co peia, Austin ,
2: 441-447.
BEMVENUTI, M.A. 1987. Abu n dân cia, distribu ição e reprodu ção de peixe-rei (Ath erin idae) n a zon a estu arin a da Lagoa dos Patos, RS, Brasil. Atlân tica, Rio Gran de, 9 (1): 5-32. BEMVENUTI, M.A. 1993. Red escrição d o p eixe-rei Odon testh es
argentinensis (Valen cien n es) Pisces: Ath erin idae, n a costa do Rio Gran de do Su l. Atlân tica, Rio Gran de, 1 5: 17-35.
BEMVENUTI, M.A. 2002. Diferen ciação m orfológica das espécies d e p e ix e s-r e i, O d on t es t h es Ev e r m a n n & Ke n d a ll (Osteich th yes, Ath erin op sid ae) n o extrem o su l d o Brasil: m orfom etria m u ltivariada. Rev istaBrasileiradeZo o lo gia, Cu ritiba, 1 9 (1): 251-287.
BERVIAN, G. & N.F. FONTOURA. 1997. Reprodu ção de Atherinella brasiliensis n o estu ário do Rio Tram an daí, Im bé, Rio Gran de do Su l, Brasil (Teleostei, Ath erin opsidae). Bio ciên cias, Por-to Alegre, 5 (2):19-32.
BRAGA, F.M.S. 1986. Estu do en tre fator de con dição e relação p eso/ com p rim en to p ara algu n s p eixes m arin h os. Rev ista Brasileirade Bio lo gia, Rio de Jan eiro, 4 6 (2): 339-346. BURBIDGE, R.G.; M.C. CARRASCO & P.A. BROW N. 1974. Age, growth ,
len gh t-weigh t relation sh ip, sex ratio an d food h abits of th e Argen tin e pejerrey, Basilichthysbonariensis (Cu v. an d Val.), from Lake Peñ uelas, Valparaíso, Ch ile. Jo urn alFishBio lo gy,
Lon don , 6: 299-305.
BURT, A.; D.L. KRAMMER.; K. NAKATSURU & C. SPRY. 1988. Th e tem -p o of re-p rodu ction in Hyphessobryconpulchripinnis (Ch ara-cidae) with a discu ssion on th e biology of ‘m u ltiple spawin g’
in fish es. En v iro n m en tal Bio lo gyo f Fish es, Neth erlan ds.
2 2 (1):15-27.
CIECHOMSKI, J.D. 1972. Reprodu ccion y fecu n didad del corn alito Austroatherinaincisa (Jen yn s, 1942) Marrero, 1950. Pisces, Ath erin idae. An ales de la So cied ad e Cien tifica Argen ti-n a, Bu en os Aires. 1 9 3 (5-6): 263-272.
CLARK, F.N. 1925. Th e life h istory of Leuresthestenuis, an ath erin e fish with tide con trolled spawn in g h abits. Califo rn ia Fish an d Gam eFish eryBulletin, Californ ia, 1 0: 1-51.
GROSS, M.R. & R.C. SARGENT.1985. Th e evolu tion of m ale an d fem ale p aren tal care in fish es. Am erican Zo o lo gist, Wa-sh in gton , 2 5: 807-822.
ISAAC-NAHUM, V.J. & A.E.A.M. VAZZOLER. 1987. Biologia reprodutiva de Micropogoniasfurnieri (Desm arest, 1823) (Teleostei, Sciaen i-dae), 2. Relação gon adossom ática, com prim en to e peso dos ovários com o in dicadores do período de desova. Bo letim do In stitutoOcean o gráfico, São Paulo, 35 (2): 123-134. LE CREN, E.D. 1951. Th e len gth -weigh t relation sh ip an d season al
cycle in gon ad weigh t an d con dition in th e pearch (Perca fluviais). Jo urn alo fAn im alEco lo gy, Lon don , 2 0: 201-219.
LOWE-MCCONNELL, R.H. 1999. Estudo seco ló gico s de co m un i-dadesdepeix estro p icais. São Pau lo, Edu sp, 534p. MALABARBA, L.R. & B. DYER. 2002. Description of th ree n ew species
of th e gen u s Odontesthes from th e rio Tram an daí drain age, Brazil (At h erin ifo rm es: At h erin o p sid ae). Ic h th y o lo g ic al Ex plo ratio n o fFresh w aters, Mü n ch en , 1 3 (3): 257-272. MORENO, C.A.; R. URZÚA & N. BAHAMONDE.1977. Breedin g season ,
sexu al rate an d fecu n dity of Basilichthysaustralis Eigen m an n 1927, from Maipo River, Ch ile. (Ath erin idae, Pisces). Studies o nNeo tro picalFaun aEn viro n m en t, Valdivia, 1 2: 217-223.
MORSE, W.W. 1980. Maturity, spawn in g, an d fecun dity of Atlan tic croaker, Micropogon ias un dulatus, occurrin g n orth of Cape Hatteras, North Carolin a. U.S. Fish ery Bulletin, Seattle, 78
(1): 190-95.
NIKOLSKY, G.V. 1963. Th eEco lo gyo fFish es. Lon don , Academ ic
Press, 352p.
PAIVA, M.P. & A.C. SCHEFFER. 1982. Matu ridade e reprodu ção do p eixe-rei Odontesthesbonariensis (Valen cien n es), n a Bacia do rio Jacuí (Brasil). Ciên ciaeCultura, São Paulo, 34 (12):
1649-1653.
VAZZOLER, A.E.A.M. 1996. Bio lo gia d a rep ro d u ção d e p eix es teleó steo s: teo ria e p rática. Marin gá, EDUEM, SBI, 169p. WHITE, B.N. 1985. Evolution ary relation sh ips of th e Ath erin opsin ae
(Pisces: Ath erin id ae). Co n tribu tio n s in Scien ce, Natu ral Histo ry Museum o f Lo s An geles Co un ty, Los An geles, (368):
1-20.
WO O TTO N, R. J. 1990. Ec o lo g y o f te le o st fish e s. New York, Ch apm an an d Hall, 404p.
WU, CC.; WC. SU & T. KAWASAKI. 2001. Reprodu ctive biology of th e dolph in fish Coriphaena hippurus on th e east coast of Taiwan . Fish eriesScien ce, Japan , 6 7: 784-793.