Performance of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) in fruits: comparison of two laboratory populations.
Texto
Imagem

Documentos relacionados
Resumo da análise de variância valores de F para porcentagem de germinação G, índice de velocidade de germinação IVG, comprimento da parte aérea CPA, comprimento radicular CRA e matéria
A PMEA/RJ dispôs sobre o Sistema Municipal de Educação Ambiental voltado para execução dessa política pública, por isso este estudo apresentou-a como um processo em disputa em
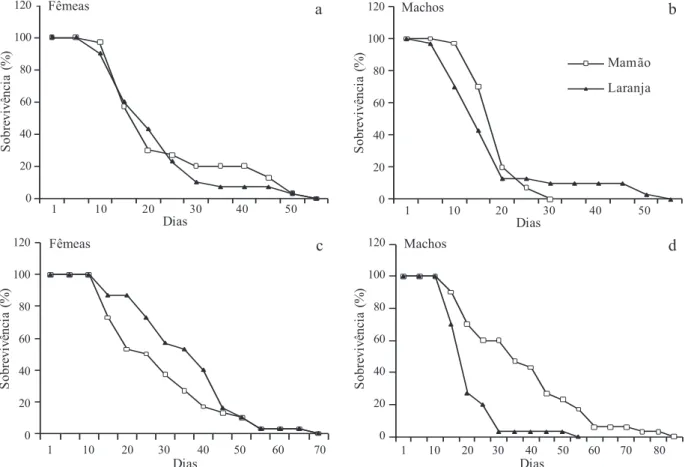
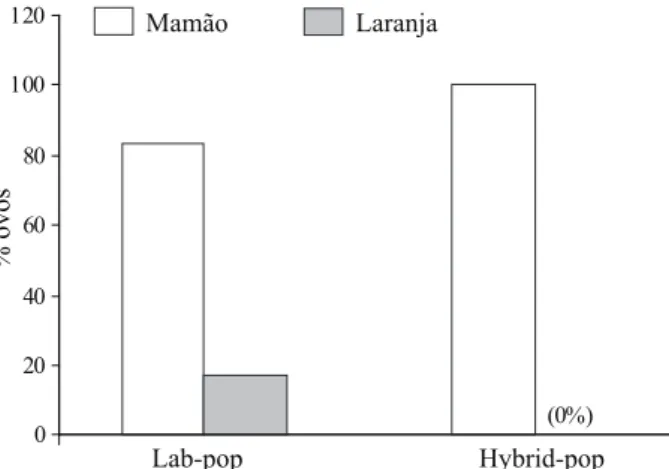
Influence of different tropical fruits on biological and behavioral aspects of the Mediterranean fruit fly Ceratitis capitata (Wiedemann) (Diptera, Tephritidae).. Studies on
A comissão de vaquejada que será composta pelos organizadores do evento, pelos organizadores das etapas e pela comissão de vaquejada da Prefeitura Municipal,
Os objetivos específicos da pesquisa são os seguintes: demonstrar que a atividade investigatória do Estado, especialmente a investigação criminal, tem fundamentos constitucionais,
Nota: Todos os programas e seus desdobramentos estão sujeitos à aprovação do CIF, no âmbito do Acordo e, quando aplicável, das autoridades competentes....
As propriedades físicas e mecânicas dos CMMs dependem, de modo geral, de inúmeros fatores como: propriedades e fração volumétrica dos componentes individuais, distribuição
• Na bebida, além do 5-CQA, foram encontradas concentrações ainda maiores dos isômeros 3 e 4-CQA. • A falta de estudos relativos à quantificação de ácidos
