Indução de resistência sistêmica por rizobactérias em cultivos hidropônicos
1Fernanda Souza Bernardes
2; Flávia Rodrigues Alves Patrício
3; Amaury Silva Santos
4; Sueli dos Santos Freitas
51Parte da dissertação de mestrado da primeira autora, apresentada ao IAC em abril de 2006. 2Centro de Energia Nuclear na Agricultura (CENA/ USP) / Laboratório de Genética de Microrganismos (ESALQ/USP), Pós-graduanda; 5Instituto Agronômico (IAC), Campinas, SP; 4Instituto Biológico (IB), Caixa Postal 28, Campinas, SP, CEP 13001-970; 5Embrapa Tabuleiros Costeiros, Aracaju, SE;
Autora para correspondência: Sueli dos Santos Freitas (sfreitas@iac.sp.gov.br). Data de chegada: 17/03/2006. Aceito para publicação em: 01/01/2010.
1338
ABSTRACT
Rhizobacteria of the fluorescent group of Pseudomonas spp. were selected for both their capa city in the gr owth promotion of lettu ce seedlings (Lactuca sativa L.) and antagonistic activity against Pythium aphanidermatum from test in vitro. Then, in hydroponic system was assayed the previous application of the Pseudomonas strains in order to reduce damages caused by P. aphanidermatum in cucumber (Cucumis sativus) for induced systemic resistance (ISR), using a technical split-root. The variables shoots and roots dry weight and root length were u sed in evalu ation. T he production of a nta gonistic a nd/or gr owth
Bernardes, F.S.; Patrício, F.R.A.; Santos, A.S.; Freitas, S.S. Induction of systemic resistance by rhizobacteria in hydroponic system. Summa Phytopathologica, v.36, n.2, p.115-121, 2010.
Keywords:Pseudomonas spp., Pythium aphanidermatum, Lactuca sativa L., Cucumis sativus L., hidrolytic enzymes.
Palavras-chave adicionais: Pseudomonas spp., Pythium aphanidermatum, Lactuca sativa L., Cucumis sativus L., enzimas hidrolíticas.
RESUMO
R iz o ba c t ér i a s do gr u p o f lu o r e s ce n t e d e Ps e u d o m o n a s sp p . f o r a m s e le c i o na d a s qu a n t o a s u a c a p a ci d a d e n a p r o m o ç ã o d e crescimento em plântulas de alface (Lac tuca sativa L.) e a tividade a nta gônica contr a Py th iu m a p h a n id e r m a tu m a pa r tir de teste i n vitro. Em seguida, em sistema hidropônico, foi avaliada a aplicação pr év ia dos i sola dos visa ndo r edu zi r os da n os pr ov oca dos p or P. a p h a n i d e r m a t u m e m pe pi no (C u c u m is s a t iv u s) p or r es i st ên ci a s i st ê m i c a i n d u z i d a ( R S I ) , p e l a a d o ç ã o d a té c n i ca “ r a íz e s subdividida s” . As va r iá veis massa da ma téria seca ( pa rte aér ea e r aízes) e comprimento r adicu lar for a m u tiliza da s na a va lia çã o. A
Bernardes, F.S.; Patrício, F.R.A.; Santos, A.S.; Freitas, S.S. Indução de resistência sistêmica por rizobactérias em cultivos hidropônicos. Summa Phytopathologica, v.36, n.2, p.115-121, 2010.
pr omoter s compou nds by r hizobacter ia str a ins wa s obser ved to a ssocia te its activity with the resu lts obta ined in the tests u nder hydroponic system and in vitro conditions. In the in vitro experiment, stra ins Ps 14 0B a nd Ps 1 40 C pr ovided the lar gest development of pla nts ( r oot a nd hypocotyls) in the tr ea tments with a nd withou t pathogen. Some strains tested indicating a possible expression of ISR in in v ivo exper iments. No corr espondence among the compou nds produced by rhizobacteria, plant growth promotion and bioprotection was detected.
Pseud omon as spp. do gru po flu orescen te são facilmente distinguíveis de outras rizobactérias, quando cultivadas em meio B de King (11), devido a sua capacidade em produzir pigmento verde-amarelado fluorescente sob luz u ltravioleta (23 ). Essas r iz o b a c t é r ia s s ã o c a p a z e s d e p r o d u z ir c o m p o s t o s c o m o hormônios, antibióticos, enzimas e sideróforos; são classificadas como “rizobactérias promotoras de crescimen to de plan tas” (RPCPs) seja por modo direto, quando na ausência de patógeno, ou in direto, pela su pressão do mesmo (27). A capacidade de adaptação metabólica e h abilidade em produ zir compostos, permitem qu e esse grupo de rizobactérias possam se adequar ao competitivo ambiente rizosférico, podendo ser empregadas no controle biológico de patógenos (14).
Segundo van Loon et al. (25), quando uma RPCPs coloniza a raiz, moléculas constituintes da célula bacteriana ou por ela s in t e t i z a d a s a g e m c o m o s in a liz a d o r e s d e n o v a s r o t a s meta bólica s. E ss as molécu las ativa m gen e s prom otores de c om p os t os d e d e fe s a, le v an do à e xp r es s ão da re sis tê n cia sistêmica induzida (RSI). Os mecanismos de defesa relacionados à R SI mediados por R P C Ps podem ser estrutu rais, como o aumento da lignificação da parede celular, dificultando a entrada e colonização do patógen o ou bioqu ímicos pela presen ça de enzimas antioxidantes (20).
O gênero Pythium, pela nova classificação e caracterização de su as fases reprodu tivas, difere dos fu n gos verdadeiros e pe rt en ce a o R e in o C h ro mist a (2 8 ) . O P e ro n o sp or om ic et o
p r od u ç ã o d e c o mp o s t os a n t a g ô n i c os e / ou p r o mo t o r e s d e crescimento pelos isola dos de Pse u d om o n as spp. flu orescente foi ve r i fic a da d e m od o a a s so cia r s u a a t iv ida de co m o s r es u lt a d os obtidos nos testes in vitro e em hidroponia. No experimento in vitro
P y t h i u m a p h a n i d e r m a t u m ( E d s o n ) Fi t z p . e s t á e n t r e o s pr in cipais pat ógen os c au sa dore s da pod ridã o ra dicu lar em c u lt iv o s h id r o p ô n ic o s , s e u s z o ó s p o r o s s ã o a d a p t a d o s a o ambiente aquáticos e estimulados por exsudados radiculares. O processo de infecção e colonização dos tecidos ocorre em menos de 24 horas. A principal forma de controle desse patógeno, que se caracteriza pela agressividade de infestação, é a preventiva (10, 15).
Sistemas hidropônicos apresentam alta umidade, densidade de cultivo e disponibilidade de nutrientes, fatores que favorecem o estabelecimen to de patógenos. E n tretan to, essas mesmas condições também são benéficas à introdução e permanência de microrganismos que atuam no controle biológico (15).
Os objetivos deste trabalh o foram: selecion ar isolados de
Pseud omona s spp. fluorescentes que promovam crescimen to vegetal e/ou atividade antagônica a P. aphanid ermatum a partir de testes in vitro; avaliar a aplicação prévia de isolados de
Pseu d o mo n as spp. em sistema h idropônico, n a redução dos danos provocados por P. aphanidermatum por RSI a partir da técnica “raízes-subdivididas” e avaliar a produção de compostos potencialmente antagônicos como ácido hidrociânico (HCN),
-1,3-glicanase, pectinase, celulase e o promotor de crescimento ácido in dol-acético (AIA).
MATERIAL E MÉTODOS
O experimento in vitro foi realizado no Laboratório de Micr obiologia do Solo do In s titu to Agron ômico (IAC ) . Os experimentos in vivo foram desenvolvidos em casa de vegetação localizada no Centro Experimental do IAC.
Experimento in vitro
Sementes de alface crespa, cv. Verônica foram desinfectadas em solu ção de h ipoclorito de sódio a 1 %. Após su cessivas lavagens em água destilada esterilizada, as semen tes foram t r a n s f e r id a s p a r a p la c a s d e P e t r i c o n t e n d o p a p e l f ilt r o , p r e v ia m e n t e u m e d e c id o e e s t e r ili z a d o . As s e m e n t e s permaneceram nas placas por 24 horas em temperatura ambiente para germinação.
S e s s e n t a is o la d os d e P s e u d o m o n a s s p p . f lu o r e s c e n t e perten cen tes à coleção de RP C P s do In stitu to Agron ômico (IAC ), em Campin as (SP ), foram transferidos para tubos de ensaio contendo meio B de King sólido e mantidos a 28ºC por 24 horas. Uma “alçada” da colônia bacteriana foi transferida para tubos contendo 5 mL de solução de MgSO4 a 0,01 mol.L-1 e h o m og e n eiz a do s em a git a do r me c â n ic o . S e me n t es p ré -ge rm in ada s fo ram a dicio n a das à s su s pe n s ões b ac ter ia n a s, permanecendo por 45 minutos.
O inóculo de P. aphanid ermatum, cedido pelo Laboratório de Fitopatologia do Instituto Biológico, em Campinas (SP), foi cu ltivado em placas de P etri contendo meio V8 e mantido a 28ºC por 96 horas. Placas de Petri contendo ágar-água receberam em seu cen tro u m disco de 1 0 mm con ten do o micélio do patógeno. As sementes pré-germinadas de alface que receberam as rizobactérias foram distribuídas (10 sementes/placa) em torno do patógeno. As placas permaneceram em temperatura ambiente por 96 h oras. Os comprimentos das raízes e dos hipocótilos foram avaliados. O delineamento foi o inteiramente casualizado, com 5 repetições, sen do cada parcela representada por uma
placa. Para cada isolado, prepararam-se 10 placas, cinco com o patógeno e cinco sem ele.
Experimento in vivo
Sementes de pepino japonês h íbrido (cv. Hokush in), após assepsia em solução de hipoclorito a 2% e sucessivas lavagens com água destilada esterilizada, foram colocadas para germinar em placas con ten do papel filtro previamen te esterilizado e umedecido. A solução nutritiva foi formulada conforme exigências nu tricion ais da cultura. No oitavo dia após a germinação, as plân tu las foram tran sferidas para o sistema h idropôn ico. A d iv isã o d o s ist e m a r a d ic u la r e a a p lic a ç ão d a s u s pe n s ã o bacteriana foram realizadas no vigésimo e vigésimo terceiro dia após a germinação, respectivamente.
As rizobactérias foram cultivadas em frascos de Erlenmeyer contendo 125 mL de meio B de King, a 28ºC por 48 horas sob agitação. Em seguida, os meios de cultura foram centrifugados durante 15 minutos a 3000 x g. O sobrenadante foi descartado e o pellet foi ressuspenso em 50 mL de solução de MgSO4 a 0,01 mol.L-1. Aplicaram-se 10 mL da suspensão em 2 L de água, volume do recipiente h idropônico. A concen tração bacteriana (in vitro e in vivo) foi estimada a partir da contagem do número d e u n ida de s f or m ad or a s de co lôn ia (U FC ) p or dilu içã o e plaqueamento em meio B de King.
No p rime ir o ex pe rim en to f or am u tiliz ad os o s is olad os Ps140B, Ps140C, Ps141A, Ps142A, Ps851D, Ps852B, Ps852C, P s 8 5 7 C , P s 8 6 4 C , P s 8 6 5 A, P s 8 6 6 A, P s 8 6 6 B e P s 8 7 1 B , escolhidos por promover ou não o crescimento vegetal em alface. No segundo experimento foram selecionados alguns isolados t e s t a d o s n o p r im e ir o e x p e r im e n t o p a r a c o n f ir m a ç ã o d o s re su lt ad os o bt id os q u a n t o à pr om oç ão d e cr es cime n t o ou controle da doença. Isolados que apresentaram caráter benéfico somen te in vitro também foram escolh idos. Desse modo, no s eg u n d o e xp er ime n t o , o s is o la do s f o ra m: P s 2 1 B , P s3 2 A, P s4 4 A, P s 4 5 A, P s 6 3 A, P s 8 9 , P s 1 4 0 B , P s 1 4 0 C , P s 1 4 1 A, Ps142A, Ps142B, Ps143C e Ps852C.
O patógeno P. aphanid ermatum com 96 horas de crescimento sobre meio V8 a 28ºC foi transferido na forma de discos de 10 mm (2 5 discos/frasco) para frascos E rlenmeyer conten do 50 mL de água destilada esterilizada. Estes permaneceram imersos por mais 96 horas a 28°C para a formação dos zoósporos. A su spen são obtida foi aplicada n os vasos hidropônicos aos 30 dias após a germinação. O número de zoósporos foi determinado em hemacitômetro após adição de 1 mL de Tween 40 em 4 mL da suspen são.
Dez dias após a aplicação do patógeno, procedeu-se à coleta das plantas, as quais tiveram seus conjuntos de raízes avaliados: massa da matéria seca (parte aérea e raízes) e o comprimento radicular.
O delineamento dos experimentos in vivo foi o inteiramente casu alizado, com 4 repetições por tratamen to. C ada parcela represen tava uma plan ta com raízes subdivididas, formando do is c on ju n to s d e ra íze s, n o q u a l u m co n ju n t o rec eb eu a ap lica ção da su sp en sã o b acte rian a e o o u tro o in óc u lo do patógeno. Os controles foram representados por plantas com e sem patógeno, ambos sem aplicação da rizobactéria.
Produção de compostos
metabólitos:ácido indol-acético (AIA), segundo Bric et al. (2); produção de -1,3-glucanase de acordo com Renwick et al. (19); e a produção de ácido hidrociânico (HCN), pectinase e celulase, pela metodologia descrita por Cattelan et al. (3).
Análise Estatística
Todos os resultados obtidos nos experimentos in vitro e in vivo
foram submetidos à análise de variância e comparação das médias pelo teste de Tukey a 5 %. Os valores de porcentagem foram previamente transformados pela aplicação do arcsen x/100.
RESULTADOS E DISCUSSÃO
Experimento in vitro
As rizobactérias u tilizadas n este trabalho foram isoladas da rizosfera de várias espécies vegetais como alface, algodão, café, tomate, soja. Entretanto, estudos anteriores de inoculação e reisolamento mostraram que não existe especificidade entre estes isolados e u ma determin ada plan ta h ospedeira, sen do capazes de colonizar a rizosfera de diferentes espécies vegetais (22; 6). Desse modo, visando otimizar os testes in vitro, optou-se por trabalh ar com alface. J á em casa de vegetação foram u t i liz a d a s p l a n t a s d e p e p in o , s e n d o e s s a e s p é c ie m u it o empregada em estudos com P. aphanid ermatum.
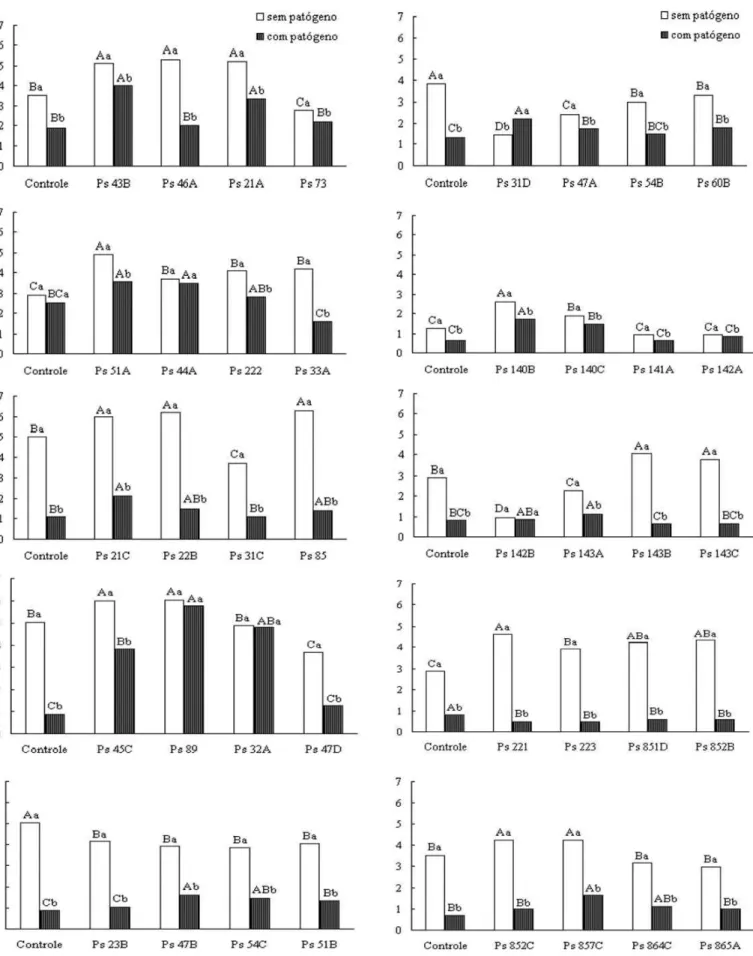
D a s 6 0 r iz o b a c t é r ia s i n o c u la d a s e m s e m e n t e s p r é -germinadas de alface, constatou-se que os isolados Ps 21C, Ps 45C , Ps 51A, Ps 44A, Ps 89, Ps 140B , Ps 140 C e Ps 8 57C promoveram crescimento radicu lar na ausência e presença do patógeno (Figura 1). Destes, apenas os isolados Ps 140B e Ps 140C – proporcionaram maior desenvolvimento radicular e do h ipocótilo (dados n ão mostrados) em plantas de alface nos tratamentos sem e com patógeno.
O contato direto en tre rizobactéria e patógen o permitiria q ue o antagon ismo se man ifestasse pela produção de algu m composto antimicrobiano ou pela competição por n utrien tes. As condições de teste incluem a planta, o que o diferencia de um teste de confron tação direto, patógeno e potencial agente de controle biológico. Essa pode ser uma forma de facilitar os testes de seleção de agentes an tagônicos a patógenos, desde q ue os resultados obtidos correspon dam aos verificados em condições de casa de vegetação.
O crescimento radicu lar dos controles de algu n s grupos avaliados n ão apresentou diferen ças entre os tratamen tos na presença e ausência do patógeno, como se a doença não houvesse ocorrido. Supõe-se que o patógeno não tenha entrado em contato com a raiz, ou o tenha tardiamente, a ponto de não comprometer o crescimento vegetal até a avaliação. Vários parâmetros como nutrientes, o pH do meio e a temperatura de incubação podem influenciar no comportamento bacteriano prejudicando a síntese de compostos que possam atuar durante a atividade antagônica, inviabilizando sua ocorrência em condições in vitro (24).
No experimento in vitro , o nú mero de UFC presentes na suspensão, onde as sementes pré-germinadas foram imersas foi de 0 ,84 x 108/mL. Nessa concentração as células bacterianas reduzem o seu crescimento, dan do in ício à fase estacionária. S e g u n d o Vin in g ( 2 6 ) , a p r o d u ç ão d e alg u n s me t a b ó lit o s secundários com caráter antimicrobiano, sob condições in vitro, in icia -s e n o f in al da fa se lo ga r ít mic a e p rin cípio d a f as e estacionária. Embora não se tenha verificado o número de células
presentes no tecido da planta, após a fase de incubação, acredita-se que a sínteacredita-se de metabólitos com atividade antagônica comece tão logo o processo de colonização do tecido vegetal se inicia, facilitando o estabelecimento das rizobactérias no ambiente.
Experimentos in vivo
Quando o tratamento controle que recebeu a inoculação do patógeno (CC P ) foi comparado com os demais tratamentos expostos ao P. aphanid ermatum e que, também, receberam o inóculo das rizobactérias em uma das partes do sistema radicular su bdividido, observou-se qu e n ão h ou ve diferen ça en tre as m as sa s da ma té ria s ec a da s r aíze s (Figu ra 2 ). P o ré m, a o confrontar com o tratamento controle na ausência do patógeno (CSP) nota-se que houve uma redução significativa na massa da matéria seca das raízes in fectadas por P. aph anid ermatu m e tratadas com os isolados Ps 21B, Ps 44A, Ps 45A, Ps 63A, Ps 140C e Ps 142B.
A observação dos resultados da massa da matéria seca das raízes den tro de cada tratamen to revelou qu e os isolados Ps 44A, Ps 89, Ps 140B, Ps 141A, e Ps 852C não apresentaram diferen ças (Figu ra 2). Essa resposta obtida pela comparação dos efeitos da ausência e presença do patógeno nas raízes de u ma mesma plan ta (“ raízes subdivididas”) pode in dicar u m modo de controle via resistência sistêmica induzida. A técnica d e r a íz e s -s u b d iv i d id a s p e r m it e q u e o s m i c r o r g a n is m o s rizobactéria e patógeno se mantenham separados espacialmente. Desse modo, não há o controle direto do patógeno pela ação de alguma substância antagônica produzida pela bactéria.
Sideróforos, assim como o ácido salicílico, podem ativar o s is t e m a d e d e f e s a d a s p la n t a s ( 1 3 ) . P o r é m , p a r a o desen volvimen to da cultu ra em sistema hidropôn ico, h á n a s o lu ç ã o d e c u lt iv o a p r e s e n ç a d e f e r r o . Sid e r ó f o r o s s ã o produzidos pelas rizobactérias em baixa concentração ou total ausên cia de ferro n o ambien te. Desse modo, é possível qu e nesse ambiente a ocorrência de RSI em plantas tratadas com
Pseud omonas sp. e cultivadas em solução nutritiva se deva a ou tro fator.
Há diversos relatos que evidenciam o efeito benéfico das rizobactérias via indução do sistema de defesa em cucurbitáceas. Plantas de pepino tratadas com Pseud omonas sp. apresentaram redução nos sintomas provocados por P. aphanid ermatum pelo at ras o e /ou dim in u içã o n a co lon iza ção do pa tóge n o. E s se fenômeno foi atribuído, embora não comprovado, à ativação de mecanismos de defesa na planta de ordem física e/ou bioquímica (5). Zhou & Paulitz (29) verificaram a ocorrência de RSI por
Pseud omonas spp. em plantas de pepino uma semana antes da aplicação do inóculo de P. apha nid ermatum, pela adoção da técnica raízes-subdivididas em cultivo hidropônico.
1). Contudo, esse comportamento distinto in vitro e in vivo é confirmado por dados da literatura (7). Nenh um dos isolados se mostrou promotor de crescimento vegetal em relação ao tratamento controle sem patógeno (CSP). Segundo Sharma & Nowak (2 1), a resistência sistêmica induzida é u m processo c o n s e gu in t e à p ro m o ç ã o d e c r e s c im e n t o, p o s s iv e lm e n t e , dependente da colonização e sensibilização do hospedeiro.
As concentrações dos zoósporos inoculados no primeiro e segundo experimento foram de 7,5 x 104 zoósporos/mL e 3,4 x 104 zoósporos/mL respectivamente. Essas concentrações estão dentro do padrão utilizado em outros estudos, qu e variam de 103 a 1 06 zoósporos/mL (4; 29). O inóculo das rizobactérias la n ç a d as s o b r e a á g u a d e ir r ig a ç ã o d o v a s o h id ro p ô n ic o apresentava uma concentração de 0,53 x 108 UFC/mL. Embora, a concentração de rizobactérias não tenha sido determinada ao término do experimento, segun do R aaijmakers et al. (1 8), a população mínima necessária de RPCPs para que ocorra a RSI é de 105 UFCs, ou seja, é preciso que ocorra uma sinalização “quorum sensing” para que a produção de compostos indutores c o m e c e m a s e r s i n t e t iz a d o s p e la s r iz o b a c t é r ia s e m u m a concentração que possa ser reconhecida pela planta. De acordo com Paulitz et al. (16), o tempo de inoculação e estabelecimento bacteriano no hospedeiro, o intervalo de aplicação do patógeno, a presente con centração dos microrganismos, assim como, a determinação do período de amostragem vegetal para análise, são fatores que influenciam na resposta a ser obtida durante o estudo do processo de defesa vegetal.
Produção de compostos
Ps eu d o mo n a s sp p. f lu or es c en te s po de m p ro du zir u m a
grande variedade de metabólitos secu n dários an tagôn icos a fungos fitopatôgenicos como antibióticos, sideróforos, enzimas h id rolíticas, c ompos tos vo láteis q u e in ibem o cr escime n to micelial etc.
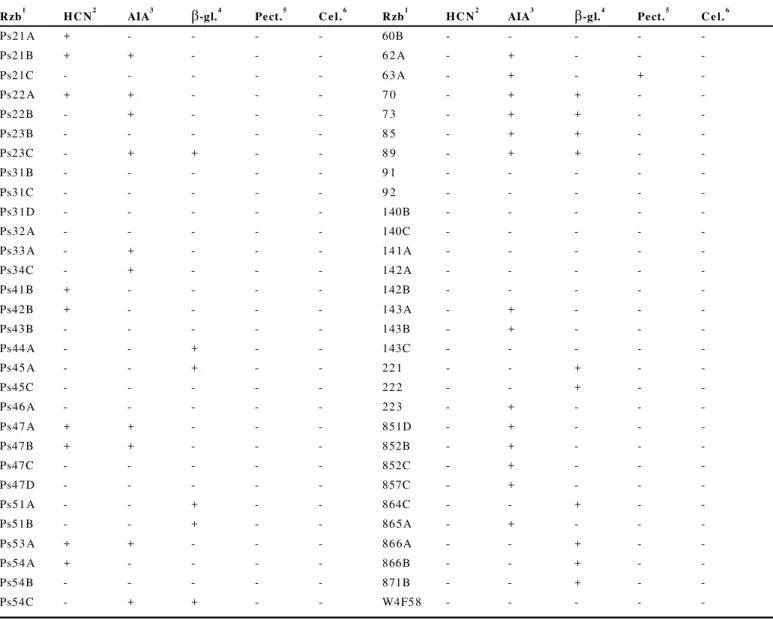
No presente trabalh o determin ou -se a produ ção de cin co compostos com caráter antagônico ou promotor de crescimento vegetal. A substância mais produzida foi o ácido indol-acético (AIA) observado em 40% dos isolados; a produ ção de ácido hidrociânico (HCN) foi verificada em nove isolados, a pectinase p or a pe n a s u m is olad o. A c elu la se n ão f oi pr od u z id a po r nenhuma das rizobactérias. Observou-se que 20% dos isolados produzem mais de uma das substâncias analisadas (Tabela 1).
No experimento in vitro 1 3% dos tratamen tos que foram e x po s t os a o p a t óg e n o e ap r e s en t a ra m ma io r c r e sc im en t o radicular, estavam inoculados com rizobactérias produtoras de
-1 ,3 glucanase. As glucanas são o principal componente da parede celular de P. aphanid ermatum e a enzima -1,3 glucanase é responsável pela degradação dessa molécula. Nesse caso, pode ser que o controle do patógeno deva-se à presença dessa enzima. A -1,3 glucanase foi produzida por 16 isolados dos isolados avaliados. Fridlender et al. (9) verificaram que Pseud omo nas cepacia, produtora de -1,3 glucanase, inibiu o crescimento de
Rhizoctonia solani, Sclerotium roffsii e Pythium ultimum. B agn asco et al. (1 ) con clu íram q u e o prin cipal fator de controle de fu ngos patogênicos em leguminosas foi devido à produção de HCN, pois isolados de P. fluorescens produ tores de sideróforos e de HCN não tiveram sua atividade antagônica afetada quando na presença de ferro. Apesar de o HCN ser um ácido volátil importante na inibição de agentes patogên icos, somen te os isolados P s 2 1 A, P s 47 A e P s 4 7B , produ tores
Figura 2. Massa da matéria seca (g) das raízes de plantas de pepino cultivadas em sistema hidropônico pela técnica raízes subdivididas, onde parte das raízes recebeu o inóculo de Pythium aphanidermatum e a outra o inóculo de isolados de Pseudomonas spp. fluorescentes.Médias de 4 repetições. Experimento 2.
* CSP: controle sem patógeno; CCP: controle com patógeno.
dessa molécu la, foram capazes de au men tar o crescimen to radicular, possivelmente, pela inibição do patógeno. Kremer & Souissi (12) observaram que rizobactérias produtoras de HCN in fluenciaram o crescimento de plântu las de alface, fato q ue pode ter se repetido neste experimento.
No experimento in vivo algumas das rizobactérias inoculadas s ã o pr o d u t o r as d e u m ou d o is d os c o m p o st o s a v a lia d o s . E n t r et a n t o, n e n h u m is ola do u t iliza d o n es t e e xp er ime n to , pro du to r de HC N e p ectin ase, su b stân cias pot en cialme n te antagônicas e que também podem ser responsáveis pela resistência sistêmica induzida (RSI), resultou em efeito benéfico às plantas. Os antibióticos produzidos por bactérias podem atuar sobre os fungos comprometendo algum processo fisiológico da célula cujo efeito pode ser fungistático ou fungicida, inibindo a germinação de esporos ou pela lise celular (24).
Embora os resultados benéficos encontrados em ambos os experimentos, in vitro e in vivo, não pudessem ser associados a
pr es en ça de co mp os tos p rod u z id os pe las r iz oba ct érias , as a n á lis e s b io q u ím i c a s p e r m it e m s e le c i o n a r is o l a d o s q u e apresen tem maior probabilidade de serem eficientes em u m p ro c es s o de co lon iza ç ão e co n tr o le bio ló g ic o . Há q u e s e c o n s id er a r q u e a p e n as a lg u m a s p ou c a s m o lé c u las f o r a m pesq uisadas: é possível qu e ou tras substân cias possam estar envolvidas e explicar os resultados obtidos.
REFERÊNCIAS BIBLIOGRÁFICAS
01. Bagna sco, P.; D e La Fuente, L.; Gua ltier i, G.; Noya, F.; Arias, A. Fluorescent Ps eu do mo na s spp. as biocontr ol a gents a ga inst for a ge legu me r oot pa thogenic fu ngi. Soil Biology and
Bio-chemis try, O x for d, v. 3 0 , n. 1 0 , p. 1 3 1 7 -1 3 2 2 , 1 9 9 8 .
02 . B r ic, J . M.; B ostock , R . M.; Silver ston, S. E. R a pid in s itu
a ssa y for indolea cetic a cid pr odu ction by ba cter ia immobili-z ed o n a ni t r oc e ll u l os e me m b r a n e . Appl i ed and E nv i
ron-Tabela 1
. Análise dos compostos produzidos pelas rizobactérias de
Pseudomonas spp. fluorescentes,
em meios de cultura
sólidos.
*A presença (+) e ausência (-) dos compostos produzidos pelas 1rizobactérias como 2ácido hidrociânico, 3ácido indol-acético, 4 β-1,3-glucanase, 5pectinase e 6celulase foram avaliadas qualitativamente pela formação do halo em torno das colônias (média de duas repetições). Rzb1 HCN2 AIA3
β
-gl.4 Pect.5 Cel.6 Rzb1 HCN2 AIA3β
-gl.4 Pect.5 Cel.6Ps21 A + - - - - 60B - - - -
-Ps21B + + - - - 6 2 A - + - -
-Ps21C - - - 6 3 A - + - +
-Ps22 A + + - - - 7 0 - + + -
-Ps22B - + - - - 7 3 - + + -
-Ps23B - - - 8 5 - + + -
-Ps23C - + + - - 8 9 - + + -
-Ps31B - - - 9 1 - - - -
-Ps31C - - - 9 2 - - - -
-Ps3 1D - - - 140B - - - -
-Ps32 A - - - 140C - - - -
-Ps33 A - + - - - 14 1A - - - -
-Ps34C - + - - - 14 2A - - - -
-Ps41B + - - - - 142B - - - -
-Ps42B + - - - - 14 3A - + - -
-Ps43B - - - 143B - + - -
-Ps44 A - - + - - 143C - - - -
-Ps45 A - - + - - 22 1 - - + -
-Ps45C - - - 22 2 - - + -
-Ps46 A - - - 22 3 - + - -
-Ps47 A + + - - - 851D - + - -
-Ps47B + + - - - 852B - + - -
-Ps47C - - - 852C - + - -
-Ps4 7D - - - 857C - + - -
-Ps51 A - - + - - 864C - - + -
-Ps51B - - + - - 86 5A - + - -
-Ps53 A + + - - - 86 6A - - + -
-Ps54 A + - - - - 866B - - + -
-Ps54B - - - 871B - - + -
-mental M ic ro bio lo gy, Wa sh i ng t o n, v. 5 7 , n. 2 , p .5 3 5 - 5 3 8 , 1 9 9 1 .
03 . C a ttela n, A. J .; H a r tel, P. G.; Fu hr ma nn, J . J . Scr eening for pl a nt gr o wth -pr o mot ing r hi zob a ct er ia to pr omot e e a r l y so y-b ea n g r ow t h. So i l Sci e nce So c ie t y o f Ame r ic an J o ur nal, Ma dison, v.6 3 , n. 6 , p.1 6 7 0 -1 6 8 0 , 1 9 9 9 .
04. Chen, C.; Bélanger, R. R.; Benhamou, N.; Paulitz, T. C . Indu-ced s ystemic r esista n ce ( I R S) by Ps e u d o m o n a s spp . impa ir s pr e- a nd p ost- infe ctio n de velo pmen t of Py th iu m a p h a n id e r -m a tu -m on cu cu mbe r r oots. European Journal of Plant Pa-thology, H ola nda , v.1 0 4 , n. 9 , p.8 7 7 - 8 8 6 , 1 9 9 8 .
05 . C hen, C .; Béla nger, R . R .; Benha mou , N .; Pau litz, T. C. De-fens e enzy mes in du ced in cu cu mber r oots b y tr ea tment with pla nt gr owt h-pr omoti ng r hizo ba cte r ia ( PG PR ) a n d Py th i u m a p h a n id e r m a tu m. Phys io lo gic al and M o le cular P lant P a-thology, Lon don, v.5 6 , n. 1 , p.1 3 -2 3 , 2 0 0 0 .
06 . Fr ei ta s, S. S.; Melo , A. M. T.; D onz eli, V. P. P r omo çã o do cr e scime nto d e a lf a ce po r r iz oba ct ér ia s . Re vist a B ras ilei ra
de Ciência do Solo, Viçosa , v. 2 7 , n. 1 , p. 6 1 -7 0, 2 0 0 3 .
07. Freita s, S. S.; Pizzinatto, M. A. Ação de r izobactér ia s sobre a incidência de C o lle to tr ic h u m g o sy ip ii e pr omoçã o de cr esci-m e nt o e esci-m p l â n t u l a s de a l g od o e i r o (G o s s y p i u m h i r s u t u m) .
Summa P hytopat ho log ic a, J a bo ti ca ba l , v. 2 3 , n. 1 , p. 3 6 -4 1 , 1 9 9 7 .
08 . Fr eitas, S. S.; Vildoso, C . I . A. R izoba ctér ia s e promoçã o do cr es ci me nt o d e pl a n ta s c ít r i ca s. Re vi st a Br as il ei ra de Ci
-ência do Solo, Viçosa , v. 2 8, n. 6 , p. 9 8 7 -9 9 4 , 2 0 0 4 .
09 . Fr idlender , M.; I nba r , J .; C het, I . B iologica l contr ol of soil-bor ne pla nt pa thogens by a -1 ,3 -glu ca na se pr odu cing Ps e u -d o m o n a s c e p a c ia. Soil Bio logy and Biochemistry, O xf or d, v.2 5 , n. 9 , p.1 2 11 -1 2 2 1 , 1 9 9 3 .
10. Glick, B .R., Penr ose, D.M.; Li, J. A model for the lowering of pla nt ethylene concentr ations by plant gr owth-pr omoting bac-te r ia . Jo urnal T heoreme Bi olog y, v. 1 9 0 , p . 6 3 -6 8 , 1 9 9 8 . 11. King, E. O.; Ward, M. K. e Raney, D. E. Two simple media for th e de mon st r a t io n of py oc ya n in a nd fl u or es ci n. Jo ur nal of
Laboratory and Clinical M edicine, St. Louis, v.4 4 , n. 2 , p. 3 0 1 - 3 0 7 , 1 9 5 4 .
12 . Kr emer, R . J.; Souissi, T. Cyanide production by rhizoba cter ia a nd potentia l for su ppr ession of weed seedling growth. Cur-rent M icrobiology, N ew Yor k, v. 43 , n. 3 , p.18 2-1 86 , 2 0 01 . 1 3. Leema n, M.; Den Ou den, F. M.; Va n Pelt, J . A.; D ir k x, F. P. M.; Steijl, H .; Ba kker, P. A. H. M.; Schipper s, B. Ir on availa-b il it y a f fe ct s i n du ct i on of s y st em i c r es is t a n c e to Fu s a r iu m wilt of r a dish by Pse ud om o na s fluo res ce ns. Phytopathology, St. Pa u l, v. 86 , n. 2 , p. 1 4 9-1 55 , 1 9 96 .
14 . O ’Su llivan, D . J .; O’G ara , F. Tra its of fluor escent Ps eu do mo -n a s s pp . i nv o l v e d i n su p p r es s i o n o f p la n t r o o t p a t h o g en s .
M i crobio lo gi cal Re vi ew, Wa s hi ng to n, v. 5 6 , n. 6 , p. 6 6 2 -6 7 -6 , 1 9 9 2 .
15. Pau litz, T. C.; Belanger, R . R. Biological contr ol in gr eenhou-se sy ste ms. Annual R evi ew of P hyt opatho log y, Pa l o Alt o, v. 39 , n. 1, p. 1 0 3-1 3 3 , 20 0 1 .
1 6. Pau litz, T. C .; Zhou , T.; R a nkin, L. Selection of r hizospher e
ba cter ia f or b iolo gica l co ntr o l of Py t h iu m a p h a n id e r m a tu m
o n h y d r o po n i c a ll y g r o w n c u c u m b e r. B i ol o g i cal C o ntro l, Amster da m, v. 02 , n. 3 , p. 2 2 6 -2 3 7 , 1 9 9 2 .
17. Pieterse, C. M. J.; Van Pelt, J. A.; Van Wees, S. C. M.; Ton, J.; Ver ha gen, B. W. M.; Léon-Klooster ziel, K.; Ha se, S.; De Vos, M.; Oosten, V. V.; Pozo, M.; Spoel, S.; Van D er Ent, S.; Koor-nneef, A.; Chalfun-Junior, A.; Resende, M. L.V.; Va n Loon, L. C . I ndu çã o de resistência sistêmica por r izobactér ia s e comu -nica ção na rota de sina lizaçã o para uma defesa r efinada.
Revi-são Anual de Patolo gia de Plantas, Pa s so Fu ndo , v.1 3 , n. 1 , p.2 7 7 -2 9 5 , 2 0 0 5 .
18. Raa ijma kers, J. M.; Leeman, M.; Van Oorschot, M. P. M.; Van der Sluis, I; Schippers, B .; Bakkers, P. A. H. M. Dose-response r ela tionships in biologica l contr ol of fu sar iu m wilt of r a dish by Pse u d om o n as spp. Phytopathology, St. Pa u l, v. 8 5 , n.10 , p. 1 0 7 5 - 1 0 8 1 , 1 9 9 5 .
1 9 . R en wi ck , A .; C a mp be ll, R .; C o e, S. A sse ss me nt of in v iv o
s cr ee ni ng s ys te ms f or p ot en ti a l b io co nt r o l a g en ts o f G a e u -m a n n o -m y c e s g r a -m in is. Plant Pathology, St. Pa u l, v. 4 0 , n. 4 , p.5 2 4 -5 3 2 , 1 9 9 1 .
20 . Romeiro, R . S.; G ar cia, F. A. O . Contr ole B iológico de enfer-mida des de pla ntas incitadas por bactérias. Revisão Anual de Patol ogi a de Plantas, Pa s so Fu ndo , v.11 , n. 1 , p.1 9 5 -2 2 8 , 2 0 0 3 .
21 . Sha rma , V.; N owa k, J. Ve rticilliu m wilt suppr ession in toma to with pseu domona d ba cter iu m. Canadian Journal of M
icro-biology, Otta wa , v. 4 4 , n. 5, 1 99 8 .
2 2 . S otte r o, A. N .; Fr eit a s, S. S.; Mel o, A. M. T.; Tr a ni, P. E. R izoba ctér ia s e a lfa ce: coloniza çã o r izosfér ica , pr omoçã o de c r es c i m e nt o e co n t r ol e b i ol ó g i c o. R e vi s t a B r as il e i r a de Ciência do Solo, Viçosa , v. 3 0 , n. 2 , p. 2 2 5 -2 3 4 , 2 0 0 6 . 2 3 . St a n ie r , R . Y; P a ll er o ni , N . J ; D o u d o r o ff , M . T he a e r o bi c
ps eu d om ona s: a t a x ono mic s tu d y. Jo ur nal Gener al of M
i-crobiology, Lo ndon, v. 4 3 , n. 2 , p.1 5 9 -2 7 1 , 1 9 6 6 .
24. Trivedi, P.; Pandey, A.; Palni, L. M. S. In vitro evaluation of antago-nistic properties of Ps eud omo nas co rru gate. Microbiological
Research, Alemanha, v. 163, n. 3, p. 329-336, 2008.
2 5 . Va n L oo n , L . C . ; B a k k e r , P. A . H . M. ; Pi e t er s e, C . M . J . Systemic r esistance indu ced by r hizospher e ba cter ia . Annual
R ev ie w of P hyt opat ho lo gy, P a l o Al to , v. 3 6 , n. 1 , p. 4 5 3 -4 8 3 , 1 9 9 8 .
2 6 . Vin i ng L . C . F u nc t io n s o f se c on d a r y me t a bo l it e s . Annual Review of M icrobiology, Pa lo Alto, v. 44 , n. 1 , p. 3 9 5-4 27 , 1 9 9 0 .
2 7 . Weller, D . M. Biologica l contr ol of soilbor n pla nt pa thogens in the r h izosph er e w ith ba cter ia . Annual Re view of
Phyto-pathology, Pa lo Alto , v.2 6 , n. 1 , p.3 7 9 - 4 0 7 , 1 9 8 8 .
28. West, P. V.; Appiah, A. A.; Neil, A. R. G. Advances in research o n o o m y c et e r oo t p a th o g e n s. P hys i o l o gi c al and M o le c