Experience with the association of volatiles with food does not affect response in three insect predators
Texto
Imagem
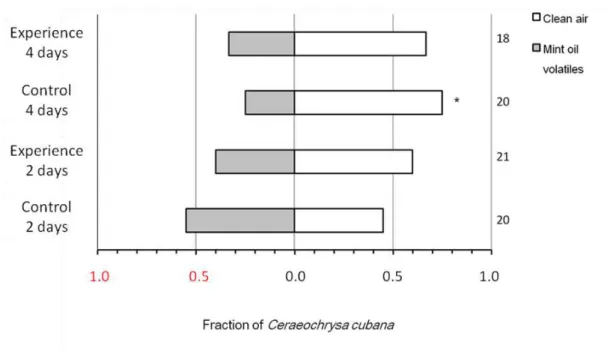
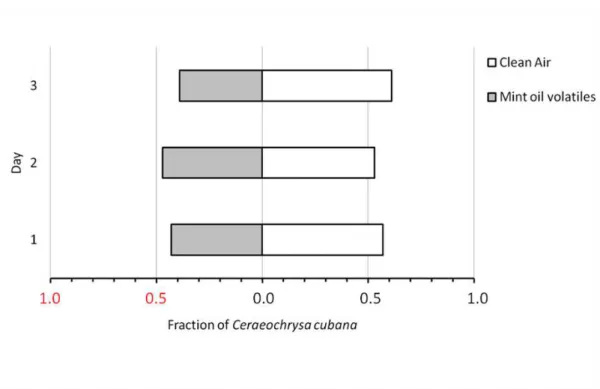
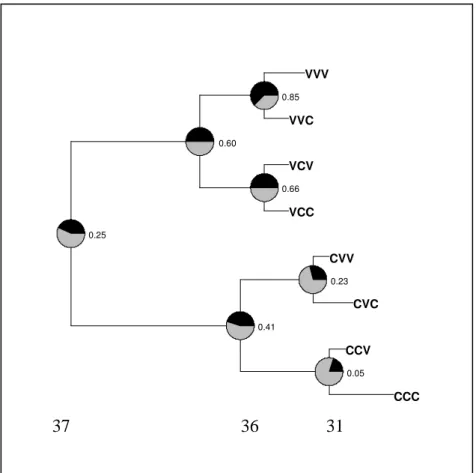
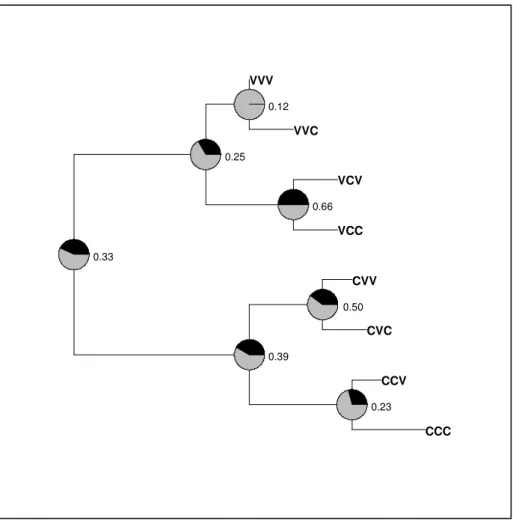
Documentos relacionados
Therefore, both the additive variance and the deviations of dominance contribute to the estimated gains via selection indexes and for the gains expressed by the progenies
están semánticamente vinculados, cada uno de estos forma parte de una unidad sémica bien diferenciada por el plano sintáctico: la estructura he de xynágousa
As doenças mais frequentes com localização na cabeça dos coelhos são a doença dentária adquirida, os abcessos dentários mandibulares ou maxilares, a otite interna e o empiema da
Como se deve tomar o banho f Depois de des- pido, é bom ficar em toilette de banho alguns minu- tos sobre a areia da praia ; durante este tempo, o ar ambiente acha-se em contacto
Para determinar o teor em água, a fonte emite neutrões, quer a partir da superfície do terreno (“transmissão indireta”), quer a partir do interior do mesmo
Neste trabalho objetiva-se apresentar um modelo ideal para gerenciamento de compras e reuso de equipamentos eletrônicos de uma IES, discorrendo sobre a mode- lagem, o desenvolvimento
Taysir Alony foi contactado mais algumas vezes por Bin Laden, para lhe entregar cassetes com declarações, que foram emitidas pela Al Jazeera, quando achou
