Análise da expressão das proteínas celulares p53, p16 e ciclina D1 em amostras de tumores penianos
Texto
Imagem
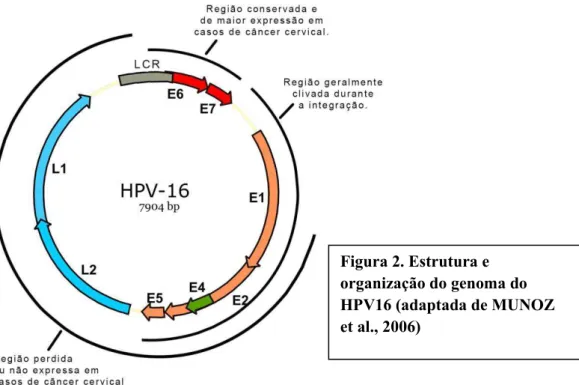
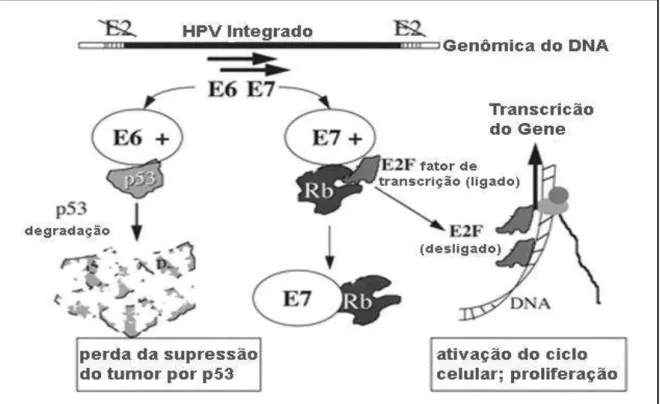


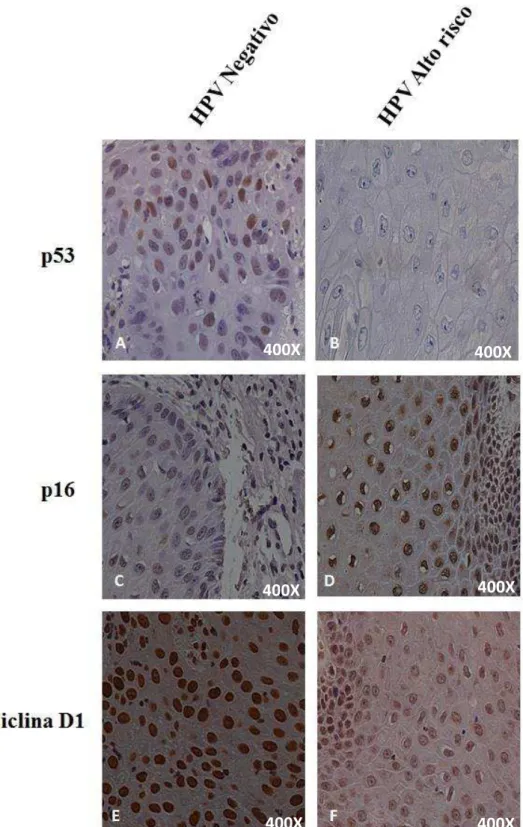
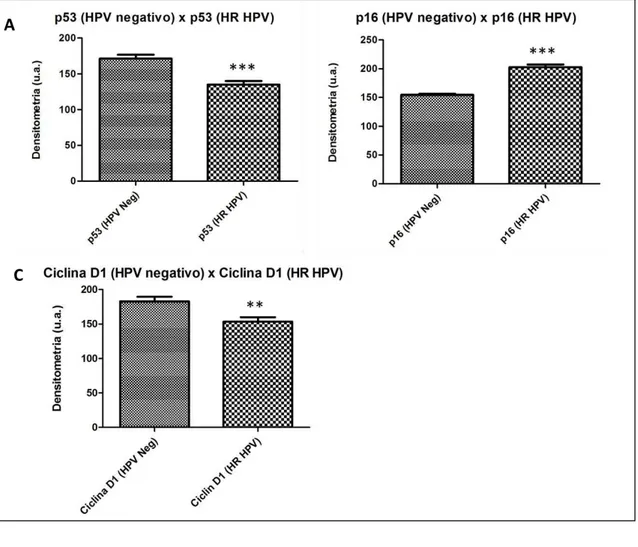
Outline
Documentos relacionados
Pena - reclusão, até cinco anos. Diante disso, o crime previsto no artigo 290 CPM se caracteriza por ser impropriamente militar, podendo ter como sujeitos
15 – Na Lisboa setecentista, são os teatros públicos de ópera (a Academia de Música fundada em 1735 por Alexandre Paghetti, que tinha por principais cantoras as próprias filhas
Permitiria, assim, em tese, o compartilhamento solidário de responsabilidades para a garantia da questão ambiental, impondo-se como uma medida mais eficaz para
5) “Estado da arte” do desenvolvimento local sertaginense. “A Sertã continua a ser pequenina e com pouca população. O setor turístico tem vindo a melhorar e temos
No sentido amplo da significação, pode-se então estabelecer um conceito geral para o constitucionalismo global (ou internacional), tal como propõe Peters (2010, p. 214), no sentido
O objetivo deste trabalho foi analisar a distribuição espaço-temporal de casos de dengue de residentes em São Luís, Maranhão, Brasil, notificados no Sistema de Informação de
We examined the emotional responses (arousal and valence) and perceived effectiveness as a function of smoking status (smokers, non-smokers), gender (male, female), and
Fonte: Instituto Médico Legal de Timon – MA Em 61% das mulheres havia múltiplas lesões Tabela 5, 25% com lesão única e 14% sem lesão As regiões do corpo da mulher onde
