DINA LILLIA OLIVEIRA DE AZEVEDO
O PAPEL DAS ROTAS E DA OBTENÇÃO
DE INFORMAÇÕES SOBRE A
EFICIÊNCIA NO FORRAGEIO DE
Dinoponera quadriceps
EM AMBIENTE
NATURAL
DINA LILLIA OLIVEIRA DE AZEVEDO
O PAPEL DAS ROTAS E DA OBTENÇÃO DE
INFORMAÇÕES SOBRE A EFICIÊNCIA NO
FORRAGEIO DE
Dinoponera quadriceps
EM
AMBIENTE NATURAL
NATAL / RN 2009
DINA LILLIA OLIVEIRA DE AZEVEDO
O PAPEL DAS ROTAS E DA OBTENÇÃO DE
INFORMAÇÕES SOBRE A EFICIÊNCIA NO
FORRAGEIO DE
Dinoponera quadriceps
EM
AMBIENTE NATURAL
Orientador: Prof. Arrilton Araújo
NATAL / RN 2009
Título: “O papel das rotas e da obtenção de informações sobre a eficiência no forrageio de Dinoponera quadriceps em ambiente natural”
Autor(a): Dina Lillia Oliveira de Azevedo
Data da defesa: 20 de agosto de 2009.
Banca Examinadora:
________________________________________
Prof. Arrilton Araújo
Universidade Federal do Rio Grande do Norte
________________________________________ Profa. Maria de Fátima Arruda
Universidade Federal do Rio Grande do Norte
________________________________________ Prof. Vincent Fourcassié
Centre de Recherches sur la Cognition Animale Université Paul Sabatier - França
_________________________________________ Prof. Iron Macedo Dantas
Divisão de Serviços Técnicos
Catalogação da Publicação na Fonte. UFRN / Biblioteca Central Zila Mamede
Azevedo, Dina Lillia Oliveira de.
O papel das rotas e da obtenção de informações sobre a eficiência no forrageio de
Dinoponera quadriceps em ambiente natural / Dina Lillia Oliveira de Azevedo. – Natal,
RN, 2009.
84 f. : il.
Orientador: Arrilton Araújo.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências.
1. Formiga (Dinoponera quadriceps) – Dissertação. 2. Forrageio - Dinoponera quadriceps – Dissertação. 3. Comportamento animal – Dissertação. I. Araújo, Arrilton. II. Título.
AGRADECIMENTOS
Agradeço à minha família: Lauro, Dilma, Daisy, Juninho e Denise, entre outros; que me incentivou, deu apoio, acompanhou e acreditou que eu chegaria ao fim desta jornada.
Ao Prof. Arrilton, que com sua sabedoria, paciência e confiança me ensinou muito mais do que comportamento animal, com seus “puxões de orelha” ele me ajudou a superar obstáculos, seu “poder de adivinhação” o fez saber o momento certo para sentar e conversar. Sem falar dos bate-papos e conselhos engraçados desfrutados não só por mim, mas por todos que costumam freqüentar a famosa salinha de Arrilton (risos). Por tudo isso, eu o considero um amigo, e espero continuar usufruindo da amizade, e orientação (não me expulse, por favor!).
Hiélia e Rosiane, que me apresentaram pessoalmente a Dinoponera quadriceps, lá em Nísia, e me deram as primeiras dicas sobre observação do comportamento.
Todos que dividiram comigo a casa em Nísia Floresta: Ana Karinne, Isabelli, Cíntia Higashi, Mary, Raika, Derlan, Audra, Jeniffer, Ana Carolina, Carolina, Danilo, Ingrid, Daniele, Helington, Edilza, Pedro, Alexandre, Anderson entre outros. Também agradeço à FLONA de Nísia Floresta pela parceria que nos permite fazer pesquisa lá e pela “nossa casa”.
À Profa. Fátima Arruda, sempre prestativa, animada, aventureira e com dicas preciosas.
À Jeniffer, como uma formiga cooperou para a realização deste trabalho. Ao Prof. Márcio Zikán, pelo software Oriana.
À Ana Karinne, Marcelo, Melquieges e Anderson, que várias vezes me socorreram nos assuntos tecnológicos.
À Júlia Maria, que muitas vezes me fez, finalmente, sair de casa para levá-la ao cinema, foram muitos filmes infantis durante esta caminhada.
Aos colegas, que várias vezes esbarramos na universidade, pelas conversas aleatórias, engraçadas e sérias: Cirleide, Nalon, Dani, Dani Torres, Jailma, Nilmara, Renan, Sidney e outros mais.
À Lorena e Mary pela amizade, que continua mesmo com a distância.
Numa jornada como essa é essencial a presença, mesmo que de vez em quando, de diversão. Então, agradeço a um grupo que se formou nestes últimos dois anos, por todas as reuniões, sempre com muita comida, e gargalhadas: Kelly (golfinha); Rosaly (abelhuda); Marcelo (passarinho); Melquieges, Priscila e Patrícia (Camarões do Forró), Audra (macaca), Isabelli, Anderson e Linda (família Dó ré mi), Antônio Francisco (o feroz), Matheus (de Audra), Bianca (ex-camarão do forró), Garcia (tubarão) e Marconi (Mopi). Amigos, que venham outras festinhas!
A duas amigas com as quais dividi muitas histórias durante este mestrado: estudos com Habbib’s; choro com música sertaneja; viagens e compras (São Paulo e Pirenópolis); e a mais perigosa delas, nadar na “piiii” do Rio Pium junto com Jorge. Meninas, esses momentos foram marcantes e marcados pelo bom humor, mesmo com a sincronização da TPM. Muito obrigada, Audra e Priscila.
Às minhas formigas que sempre cooperaram comigo. Apesar do susto. À UFRN. Ao Programa de Pós-graduação em Psicobiologia. Ao CNPq.
RESUMO
ABSTRACT
SUMÁRIO
1. INTRODUÇÃO ... 10
2. OBJETIVOS E HIPÓTESES ... 21
2.1. Objetivo ... 22
2.1.1. Objetivos específicos ... 22
2.2. Hipóteses e Predições ... 22
2.2.1. 1ª Hipótese ... 22
2.2.2. 2ª Hipótese ... 22
3. RESULTADOS / ARTIGOS ... 23
3.1. Artigo 1º ... 24
3.2. Artigo 2º ... 49
4. DISCUSSÃO GERAL E CONCLUSÕES ... 71
Todo animal diariamente realiza atividades como forrageio e descanso. As atividades desempenhadas longe do local de descanso ou ninho podem trazer riscos que comprometem a sobrevivência do indivíduo, um risco eminente é o da predação. Todavia, os benefícios adquiridos em tais atividades devem provavelmente superar os custos e fazer valer a pena para o animal arriscar-se. A fim de minimizar os riscos, os animais fazem uso de estratégias que os auxiliam no aproveitamento máximo de tempo e energia; principalmente na atividade de forrageio que é uma das mais importantes para garantir a sobrevivência de todo e qualquer animal.
Na exploração de uma área particular durante o forrageio, a variável de decisão do animal é o tempo que ele deve gastar em tal área, ou melhor, permanecer na área. O tempo é uma variável crítica para os animais (Stephens & Krebs 1986). A decisão de um forrageador poderia ser estruturada com relação ao tempo gasto na área escolhida, porém, esta assunção induz ao erro quando não existe ligação entre tempo de permanência e quantidade de alimento adquirido no local (Stephens & Krebs 1986).
divisão do alimento encontrado pode garantir maior sobrevivência do que cada um por si. Se oportunistas diferentes procuram recursos diferentes, cada indivíduo busca como especialista e se alimenta como generalista, gerando assim o problema da soma de experiência, um indivíduo pode simplesmente se aproveitar do trabalho do outro (Giraldeau & Caraco 2000). Uma forma de sobrepujar este problema é manter relações mútuas com indivíduos que podem ser identificados, como os parentes, por exemplo.
Uma teoria muito utilizada para explicar a otimização do forrageio de vários grupos animais é a teoria do forrageio de ponto central. Ela assume que o tamanho da carga não afeta o tempo de viagem e conseqüentemente a velocidade. A teoria prediz que com o aumento da distância da fonte de recurso ao ninho, um indivíduo terá que selecionar presas de tamanhos maiores e/ou de conteúdo mais rico em energia para compensar os custos energéticos e o investimento de tempo envolvido na viagem a distâncias maiores (Roces 1990; Bonser et al. 1998). Outra teoria comum é o teorema do valor marginal, que prediz que um predador deve deixar a mancha logo que a taxa marginal de captura declina(Charnov 1976). Neste modelo os forrageadores devem buscar presas a grande distância e carregar pesos maiores de alimento de alta qualidade de volta ao ninho, comparado ao de baixa qualidade. Logo, o tempo para capturar e carregar as presas seriam maiores em trechos distantes do que próximo ao ninho (Bonser et al. 1998). Ela é proposta para um predador cujo tempo é gasto no movimento entre manchas (Charnov 1976).
coletam alimento independentemente, mas viajam juntos ao longo de trilhas ou colunas a partir da colônia (Bernstein 1975).
Insetos sociais, como abelhas e formigas, costumam ser utilizados como modelo para estudos sobre estratégias de forrageio. Os insetos eusociais, que incluem todas as formigas e cupins e algumas abelhas e vespas, podem ser distinguidos como um grupo por possuírem três características em comum: cooperação no cuidado dos jovens; divisão reprodutiva do trabalho; e, sobreposição de pelo menos duas gerações capazes de contribuir para o trabalho da colônia (Wilson 1971).
As formigas são separadas das abelhas e vespas por uma diferença estrutural: a presença de uma cintura longa entre o tórax e o abdômen conhecida como pecíolo (North 1996). Elas pertencem a um táxon amplo de aproximadamente 10.000 espécies, vivendo através de grandes extensões de condições ecológicas que refletem no seu comportamento de forrageio (Schafer et al. 2006). Dos 296 gêneros de formigas descritos no mundo inteiro, 118 encontram-se na região neotropical e destes, 60 são endêmicos desta região; os demais ou têm distribuição mundial ou são compartilhados com o continente africano (Caetano et al. 2002).
indivíduos que pode ocorrer através da morfologia ou da idade. Desta maneira, as operárias individuais executam apenas um número pequeno de tarefas em qualquer tempo, minimizando os erros que possam ser cometidos e organizando o trabalho da colônia (North 1996).
Os ninhos das formigas variam amplamente entre as espécies. Existem nidificações arbóreas, como ninhos construídos nos troncos das árvores, que aos olhos dos leigos parecem cupinzeiros, ou mesmo vivendo em bromélias e, terrestres, que podem ser dentro de cavidades de troncos caídos, sob folhas ou subterrâneos com arquiteturas surpreendentes. As formigas podem ocupar um ninho único ou múltiplo. Esta última condição é conhecida como polidomia (Holway & Case 2000). Devido à estrutura descentralizada, as colônias polidômicas podem se livrar de certas restrições associadas ao forrageio de ponto central, pois a dispersão do ponto central provavelmente melhora a eficiência do forrageio. A polidomia foi registrada nas espécies Camponotus gigas (Pfeiffer & Linsenmair 1998), Linepithema humile (Holway & Case 2000), Cataglyphis iberica (Cerdá et al. 2002), Tapinoma sessile (Buczkowski & Bennett 2006), Pachycondyla goeldii (Denis et al. 2007) e Iridomyrmex purpureus (Wilgenburg & Elgar 2007).
somente na orientação para o ninho, como também para os recursos alimentares encontrados (Vilela et al. 1987). As formigas podem reagir sensivelmente a vários odores, e é provável que elas possam perceber heterogeneidades olfativas no ambiente e usá-las para orientação (Ehmer 1999).
Além da orientação por pistas químicas, as formigas podem fazer uso de pistas celestes, como uma bússola, e também sinais visuais como pontos de referência. Esta última é provavelmente comum à maioria das espécies de formiga. Elas podem assim, se orientar na presença de uma trilha com o auxílio adicional de pistas visuais diretas como demonstrado pela Atta, que mesmo na ausência de uma trilha química completa a orientação para o ninho, apesar de gastar um tempo de retorno significativamente mais longo (Vilela et al. 1987). Em Paraponera clavata, as operárias usam, preferencialmente, pistas de ponto de referência local durante a orientação e apresentam uma hierarquia das pistas que mudam com a experiência da operária (Harrison et al. 1989). Possivelmente, a orientação baseada em pistas visuais aumentará a velocidade de forrageio e, pode também ser mais estável ao longo do tempo e menos susceptível a interrupções ambientais (Harrison et al. 1989). Um exemplo do uso de bússola solar é encontrado na formiga do deserto Cataglyphis fortis, que conta não só com uma bússola, mas também com algum tipo de velocímetro, visto que esta espécie tem como meio predominante de navegação a integração da trilha, atualizando continuamente um vetor de integração do limite da área que informa sua posição relativa ao ninho (Knaden & Wehner 2005).
comportamental, anatômica e fisiológica da espécie. Temperatura do solo, umidade, radiação e vento são os principais fatores abióticos que os afetam (Pol & Casenave 2004).
Todas as formigas pertencem a uma única família, Formicidae. Dentre as várias subfamílias de formicídeos está a Ponerinae, subdividida em tribos. As Ponerinae distribuem-se por todo o mundo, porém são mais abundantes nas regiões tropicais. As formigas desta subfamília são, geralmente, predadoras (Caetano et al. 2002), mas, podem também se alimentar de néctar extrafloral, substância doce secretada por homópteros, secreções de larvas de lepidópteros, bem como sementes ricas em lipídios. Os modos de forrageio se estendem de solitário a grupo de caçadoras, e de predadores especialistas a generalistas (Fourcassié & Oliveira 2002). Elas possuem um ferrão funcional, com o qual submetem a sua presa. Juntamente com a subfamília Cerapachyinae, seus representantes são os mais primitivos da região neotropical, tanto por sua morfologia como pelo seu comportamento (Caetano et al. 2002). Todas as tribos pertencentes à subfamília Ponerinae demonstram níveis de organização social que são comparáveis àqueles das subfamílias de formigas consideradas, filogeneticamente, mais evoluídas (sensu Morgan et al. 2003), unidos a uma comunicação química elaborada (Morgan et al. 2003).
As espécies de Ponerinae do gênero Neotropical Dinoponera Roger (1861) estão entre as maiores formigas conhecidas, atingindo ou mesmo ultrapassando o comprimento de 3 cm. A distribuição do gênero está restrita à América do Sul e, o território inclui o sudeste da Colômbia, leste do Peru, todo o Brasil, leste da Bolívia e do Paraguai e nordeste da Argentina. A espécie Dinoponera quadriceps Santschi (1921) é endêmica da área brasileira conhecida como “polígono das secas” no nordeste do Brasil. O tamanho e as medidas das operárias de Dinoponera quadriceps têm proporção pouco menor do que da
abdômen menos brilhante e na forma peculiar do pecíolo, que tem sua extremidade anterior marcante e estreitamente arredondada e, sua extremidade posterior amplamente arredondada (Kempf 1971).
Em aproximadamente 100 espécies de formigas pertencentes a 10 gêneros, todos na subfamília Ponerinae, a casta de rainha morfologicamente especializada está ausente (Peeters 1997). Dinoponera quadriceps é uma espécie de formiga sem esta casta, conseqüentemente todas as operárias são capazes de reproduzir. Todavia, sua reprodução está sob controle hierárquico, operárias com postos de dominância diferentes têm perfis comportamentais distintos e o posto se correlaciona com a atividade de postura de ovos (Monnin & Peeters 1999). Ao contrário de outras espécies, a alfa virgem de Dinoponera
quadriceps não exibe qualquer postura de chamado sexual, visto que ela é a única operária
atrativa para os machos; entretanto, ela provavelmente também libera feromônios sexuais. Após a cópula, a genitália do macho permanece ligada ao abdômen da operária e bloqueia o acesso por rivais potenciais. A presença de uma operária sexualmente atrativa em colônias
de Dinoponera quadriceps não é regulada sazonalmente, pois a operária beta alcança o
posto alfa depois da morte da gamergate (denominação da operária alfa após a reprodução), antes ou depois da fissão da colônia e, somente assim pode acasalar (Monnin & Peeters 1998). A gamergate provavelmente vive de 1-2 anos. Em uma colônia estabelecida ela é a mãe de todas as operárias e, estas por sua vez são irmãs completas (Monnin & Peeters 1999).
Dinoponera quadriceps nidifica no solo e suas operárias forrageiam solitariamente e
Rodrigues 2006). Este mesmo padrão foi observado para as forrageadoras da Dinoponera
gigantea (Fourcassié et al. 1999). Na Dinoponera gigantea as trilhas químicas não são
indispensáveis à orientação na vegetação. As pistas de orientação principais parecem ser dadas pelos pontos de referência do ambiente (Fourcassié et al. 1999). O ambiente tropical em que ambas Dinoponera quadriceps e Dinoponera gigantea vivem, provavelmente, oferece uma abundância de pontos de referência visuais. Assim, as formigas podem ser capazes de se orientar pelo uso somente de um subconjunto destes pontos de referência permitindo flexibilidade maior na sua orientação.
Pontos de referência familiares não são desligados do contexto dentro do qual eles tenham sido adquiridos e não são unidos de forma generalizada ou como um mapa. Eles instruem as formigas, preferivelmente, do que provêm informações iguais a um mapa (Wehner et al. 2006). Procurando solitariamente, as formigas podem ater-se, para se orientar em seu ambiente, exclusivamente na integração da trilha, mecanismo pelo qual o animal em movimento atualiza a localização de um objetivo memorizado usando informação baseada na rota, que é a informação de seus próprios movimentos (Benhamou & Poucet 1996); ou na memória espacial de pontos de referência.
A duração da viagem da Dinoponera quadriceps indiretamente demonstra o nível de investimento de uma operária na atividade extra-ninho, especialmente naquelas relacionadas à busca e coleta de alimento (Araújo & Rodrigues 2006). Um dos efeitos imediatos do transporte de alimento nas formigas é a redução da velocidade de locomoção. Depois de encontrar e capturar a presa, as operárias retornam ao ninho com uma trajetória mais linear e uma velocidade maior.
A velocidade de locomoção das operárias pode ser afetada pela distância da área de forrageio e o transporte de alimento, conforme visto para as operárias de Dorymyrmex
goetschi, que são capazes de carregar alimentos equivalentes a 3,5 vezes sua massa
corporal (Torres-Contreras & Vasquez 2004).
Nas formigas coletoras, que ajustam o esforço de forrageio à disponibilidade de alimento para minimizar a perda de água, o grau de velocidade em que as forrageadoras retornam ao ninho está ligado também à disponibilidade de alimento - quando o alimento é fácil de encontrar, as forrageadoras retornam mais rapidamente (Schafer et al. 2006). Em viagens prévias, Ectatomma ruidum parte em uma direção até alcançar a distância máxima da entrada do ninho. Caso ela capture alguma presa durante esta viagem de longo curso, ela retorna ao ninho em alta velocidade e pouco mais tarde ela parte novamente para o mesmo local, mas, desta vez ao longo de um caminho mais direcionado e com velocidade aumentada em relação à viagem anterior. Se nenhuma presa for capturada, uma forrageadora retornará ao ninho sem presa, algumas vezes depois de uma hora e muitos círculos de procura e a viagem subseqüente será retardada e não direcionada com relação à direção e velocidade (Franz & Wcislo 2003).
2.1 OBJETIVO:
Investigar o papel das rotas e da obtenção de informações introduzidas no ambiente, sobre a eficiência no forrageio de operárias de Dinoponera quadriceps em ambiente natural.
2.1.1 Objetivos específicos:
Verificar se as rotas de forrageio têm relação com o tempo gasto pelas operárias durante a atividade extra-ninho. (artigo 1)
Analisar a influência do desempenho das operárias no tempo de atividade extra-ninho e na rota realizada pelas mesmas. (artigo 1)
Observar a integração de novas informações do ambiente que podem influenciar na otimização da busca de alimentos. (artigo 2)
2.2 HIPÓTESES E PREDIÇÕES:
2.2.1 1ª Hipótese - As distâncias percorridas pelas operárias têm relação com o tempo de
busca por alimento.
Predições:
Quanto maior a distância que a operária percorre, mais tempo ela gasta buscando alimento.
2.2.2 2ª Hipótese - As operárias integram informações do ambiente que facilitam o desvio
de obstáculos.
Predições:
3.1ARTIGO 1º
TÍTULO: EFICIÊNCIA NA ATIVIDADE DE FORRAGEIO DE Dinoponera quadriceps SANTSCHI (1921) (FORMICIDAE, PONERINAE)
AUTORES: AZEVEDO, D. L. O., MEDEIROS, J. M. & ARAÚJO, A.
Programa de Pós-graduação em Psicobiologia, Departamento de Fisiologia, Universidade Federal do Rio Grande do Norte.
PERIÓDICO: BEHAVIORAL ECOLOGY
Qualis: A2 no Comitê de Psicologia da CAPES Fator de impacto: 3.224
EFICIÊNCIA NA ATIVIDADE DE FORRAGEIO DE Dinoponera quadriceps
SANTSCHI (FORMICIDAE, PONERINAE)
RESUMO
Palavras-chave: Dinoponera quadriceps, teoria do ponto central, teorema do valor marginal, fidelidade direcional, tomada de decisão, estratégia de forrageio.
INTRODUÇÃO
De acordo com a teoria de otimização, cada estratégia de forrageio selecionada representa o melhor balanço executável de custos e benefícios que maximizam o lucro energético liquido para a colônia (Detrain e Deneubourg 1997). Forrageadores solitários exploram uma proporção maior de manchas disponíveis do que forrageadores em grupo, porque várias manchas podem ser exploradas concomitantemente por forrageadores independentes (Beauchamp 2005). A interdependência econômica significa que a recompensa através de uma estratégia de forrageio depende simultaneamente do comportamento de todos os competidores (Giraldeau e Caraco 2000). Outra característica importante dentro da estratégia adotada é a orientação. Jander (1975) definiu que orientação espacial é a manutenção do controle próprio ou mudança na posição do corpo relativo ao espaço ambiental e ela é composta de orientação da direção e distância.
pode resultar de estimativas brutas do tamanho da presa e regras de decisões simples no nível individual (Detrain e Deneubourg 1997).
As estratégias de forrageio na subfamília Ponerinae não refletem relação filogenética e são provavelmente o resultado de pressões seletivas únicas enfrentadas pelas espécies (Hölldobler e Wilson 1990). Nesta subfamília, o modo de forrageio se estende de solitário a grupo de caçadoras e de predadores especialista a generalista (Fourcassié e Oliveira 2002). As espécies do gênero neotropical Dinoponera estão entre as maiores formigas conhecidas, podendo ultrapassar o comprimento de 3 cm. A distribuição desse gênero está restrita à America do Sul. A espécie Dinoponera quadriceps (Santschi) é endêmica da área brasileira conhecida como “polígono das secas” (Kempf 1971). D.
quadriceps nidifica no solo e suas operárias forrageiam solitariamente; não realiza em
nenhum momento o recrutamento de companheiras de ninho. Araújo e Rodrigues (2006) verificaram que D. quadriceps deixa a colônia com velocidade de deslocamento lenta e inicia o forrageio logo após a saída. Devido a estes aspectos do forrageio, essa espécie despertou nosso interesse sobre o quanto sua estratégia de forrageio torna eficiente o trabalho da colônia. Assim, partindo de algumas variáveis de decisão individual pretendemos verificar a eficiência da atividade de forrageio das operárias de D. quadriceps, considerando dois modelos possíveis: modelo do forrageio de ponto central com provisionamento e o modelo do teorema do valor marginal.
MATERIAL E METODOS
Local de estudo
sede do município de Nísia Floresta, Rio Grande do Norte, Brasil. A região tem um fotoperíodo claro:escuro de 12:12h ± 15min. A estação de pesquisa abrange uma área de 180 ha, dos quais 80 ha são de Mata Atlântica secundária e 40 ha de plantações de espécies exóticas (Santee e Arruda 1994).
Sujeitos
Duas colônias de Dinoponera quadriceps (Hymenoptera, Formicidae, Ponerinae) foram escolhidas aleatoriamente dentro da área de mata secundária. Uma das colônias possuía mais de uma entrada. Neste caso, escolhemos a primeira entrada que foi encontrada como o ponto de referência das observações. Elas estavam estabelecidas a 13,1 m de distância uma da outra.
As operárias foram marcadas ao longo do estudo. À medida que surgia uma formiga sem marca, essa era capturada e em seguida recebia uma etiqueta plastificada e numerada, colada na região do tórax, utilizando uma cola à base de éster de cianoacrinolato (baseado em Corbara et al. 1986). As operárias de colônias diferentes eram identificadas por etiquetas de cores distintas.
Registro comportamental
As observações das colônias aconteceram durante 19 meses, não contínuos, entre fevereiro de 2006 e junho de 2008, pelo menos uma vez por semana, dez horas por dia. Observavamos uma operária por vez, desde a saída até o retorno ao ninho. Para tais observações foi adotada a técnica do focal instantâneo com janelas de observação de quinze minutos e registro a cada minuto, com intervalos de cinco minutos entre as janelas (Martin e Bateson 1994).
• Forrageio: a operária se movimentava vagarosamente, tocando suas antenas no
substrato vasculhando a área em rota tortuosa;
• Captura e transporte do alimento: a operária capturava um recurso alimentar, de
origem vegetal ou animal – vivo ou morto – com a mandíbula;
• Comunicação: duas ou mais operárias, de uma mesma colônia, ao se encontrarem
apresentavam o toque das pernas e/ou antenas;
• Interação agonística: duas ou mais operárias, de colônias diferentes, durante o
encontro prenderam suas mandíbulas uma na outra, tocaram suas antenas e primeiro par de pernas, dobraram o abdômen para tentar esfregar na adversária, ou rolaram juntas no substrato antes de se separarem;
• Marcação química: a operária esfregava a parte ventro-anterior ou ventro-posterior
do abdômen no substrato;
• Parada: quando não havia deslocamento da operária e, poderiam ocorrer movimentos
antenais de um lado para outro, bem como direcionar a cabeça no sentido vertical;
• Locomoção: deslocamento mais acelerado e linear da operária, sem que ocorresse
toque das antenas no substrato.
• Limpeza: a operária passava uma das pernas ou ambas, do primeiro par, nas antenas,
cabeça, abdômen ou pernas; poderia ou não antes disso passá-las na probóscide (entre as mandíbulas).
tempo transcorrido entre o retorno ao ninho e a viagem seguinte. Uma operária poderia ser observada mais de uma vez em dias diferentes. Com exceção da marcação, não houve qualquer manipulação das operárias em relação ao seu percurso e suas atividades extra-ninho.
Registro do deslocamento das operárias
Para mapear os deslocamentos das formigas adaptamos a técnica descrita por Turchin et al. (1991) de modo a assentar bandeirolas numeradas, a cada cinco minutos, durante a observação. Após o retorno da operária ao ninho, eram feitas as medidas de distância e ângulo entre as bandeirolas, usando bússola e trena. A partir desses dados, construímos o percurso estimado de cada operária durante suas atividades, obtivemos os dados da distância total percorrida, da distância do local de captura até o ninho, a maior distância em linha reta da entrada do ninho e a direção utilizada nas atividades de forrageio bem como a fidelidade a esta direção. Com o tempo de retorno ao ninho mais a distância entre ele e o local de captura, calculamos a velocidade média do transporte da presa em cada viagem bem sucedida.
Estimativa do tamanho da presa
Quando uma operária capturava e transportava algum item alimentar, era registrado o tamanho aproximado do mesmo. Este tamanho aproximado das presas foi definido a partir de uma escala construída em função das proporções do tamanho da D. quadriceps. Para tanto foram medidas 23 operárias capturadas em outros ninhos, gerando a seguinte escala (média ± DP) (Fig. 1):
• Proporção 0 - significa que retornou sem alimento;
• Proporção 1- item com medida equivalente ao comprimento da mandíbula até o final da
• Proporção 2- medida equivalente ao comprimento da mandíbula até o final do pecíolo:
16,93 ± 1,44 mm;
• Proporção 3- medida equivalente ao comprimento da mandíbula até o final do
abdômen: 25,83 ± 1,49 mm;
• Proporção 4- item maior do que o comprimento total da operária.
Figura 1. Ilustração da escala de proporções construída segundo o tamanho das operárias
de Dinoponera quadriceps.
Analise estatistica
As variáveis que escolhemos para analisar a eficiência na atividade de forrageio foram: o tempo de permanência da operária fora do ninho, que consistiu no somatório dos tempos de busca de presas e de retorno durante uma viagem; a distância total percorrida pela operária durante o forrageio e o retorno ao ninho; e a direção do forrageio foi verificada utilizando-se os ângulos obtidos entre cada bandeirola das viagens, partindo da entrada do ninho como referência.
Para verificarmos a existência de relação entre tempo de permanência fora do ninho e distância percorrida, realizamos o teste de correlação de Spearman (rs), com todas as
variáveis acima citadas, como também para a latência de saída, a fim de analisarmos a diferença no sucesso das viagens, em relação ao retorno com ou sem alimento.
Para investigarmos se existiu uma preferência das operárias às áreas especificas em volta das colônias, aplicamos o teste de Rayleigh. Para a realização desse teste, consideramos somente as operárias que obtiveram no mínimo 30 pontos de aposição das bandeirolas durante o deslocamento, independente do numero de viagens realizadas. Após a análise individual das operárias, nós consideramos o valor do vetor médio a fim de agruparmos as formigas em setores de atividade. Desta maneira, dividimos a área de forrageio, arbitrariamente, em quatro setores de 90°, em seguida realizamos novamente o teste de Rayleigh (Z) para apresentarmos o resultado da fidelidade ao setor. O teste de Watson U2 (U2) foi aplicado para determinarmos a distribuição uniforme das operárias das colônias na área de forrageio (Batschelet 1981). Consideramos, em todas as análises, o nível de significância de 5% bicaudal. Utilizamos para a realização dos testes os software Statistica 7.1 (StatSoft 2005) e Oriana 2.01 (Kovach 2004).
RESULTADOS
quando a operária saia do ninho depois de uma viagem prévia bem sucedida. Já a marcação foi observada, principalmente, nas áreas onde havia sobreposição da atividade de operárias de colônias diferentes. Devido à baixa freqüência de registros de captura e transporte do alimento, por causa da técnica utilizada, decidimos englobar estes dois comportamentos à atividade de forrageio, para analisarmos a duração e a distância percorrida em sua totalidade.
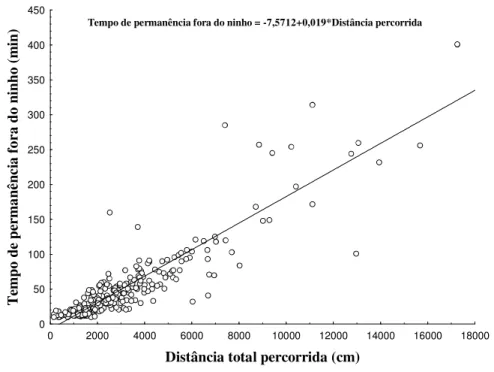
Durante o forrageio, o tempo de permanência fora do ninho e distância total percorrida apresentaram correlação positiva (N=264, rS=0.88, p<0.05) (Fig.2). O tempo de
permanência fora do ninho teve a mediana em 36 min (quartis - 25%: 21.0 - 75%: 62.3 min, extensão: 10 e 401 min). Já a distância percorrida teve sua mediana em 2461 cm, (quartis 25%: 1506 – 75%: 3833 cm, extensão: 152 a 17256 cm).
0 2000 4000 6000 8000 10000 12000 14000 16000 18000
Distância total percorrida (cm)
0 50 100 150 200 250 300 350 400 450 T em po d e pe rm an ên ci a fo ra d o ni nh o (m in
) Tempo de permanência fora do ninho = -7,5712+0,019*Distância percorrida
Figura 2. Relação entre tempo de permanência (min) fora do ninho e distância percorrida
(cm) durante o forrageio.
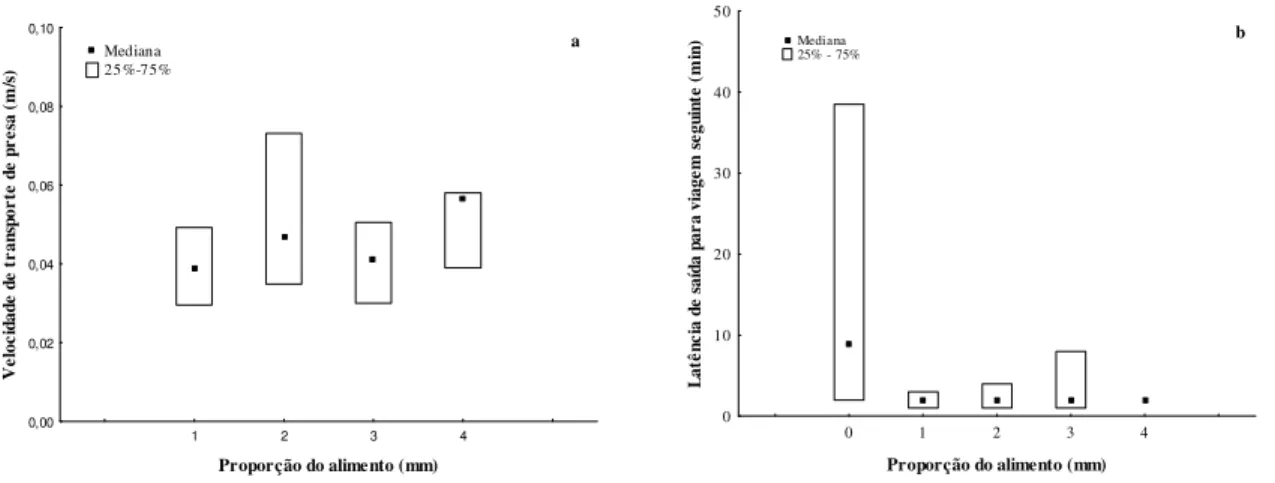
57% delas as operárias retornaram com alimento. Encontrar e capturar um alimento de maior tamanho requereu um período de forrageio mais prolongado (N=264, H= 13.41, df=4, p= 0.009)(Fig. 3a), bem como distâncias maiores (N=264, H= 11.85, df=4, p= 0.018) (Fig. 3b). Durante as observações, registramos ocasionalmente operárias, aparentemente, ingerindo um item alimentar capturado, quando este era tão pequeno que mal dava para ser notado, em seguida, continuava o forrageio.
0 1 2 3 4
Proporção do alimento (mm)
0 50 100 150 200 250 300 350 T em p o de p er m an ên ci a fo ra d o ni nh o (m in ) Mediana 25% - 75%
a
0 1 2 3 4
Proporção do alimento (mm)
0 2000 4000 6000 8000 10000 12000 D is tâ nc ia t ot al p er co rr id a (c m ) Mediana 25% - 75%
b
Figura 3. Avaliação da eficiência do trabalho das formigas em relação ao tempo que elas
permaneceram forrageando (a) e às distâncias percorridas durante o forrageio (b).
1 2 3 4
Proporção do alimento (mm)
0,00 0,02 0,04 0,06 0,08 0,10 V el oc id ad e de t ra ns po rt e de p re sa ( m /s
) Mediana 25%-75%
a
0 1 2 3 4
Proporção do alimento (mm)
0 10 20 30 40 50 L at ên ci a de s aí da p ar a vi ag em s eg ui nt e (m in ) Mediana 25% - 75%
b
Figura 4. Avaliação do efeito do sucesso em uma viagem sobre a seguinte, em relação à
velocidade de transporte (a) e à latência de saída para uma nova busca (b).
intervalo entre as viagens era menor quando a operária retornou da viagem anterior com alimento (N=152, H=36.84, df=4, p=0.001) (Fig. 4b). Nem o tamanho do alimento em si influenciou na redução da latência de saída do ninho (Fig. 4b), nem a velocidade de transporte do alimento na viagem prévia apresentou diferença em relação ao tamanho da presa (N=153, H=5.51, df=3, p=0.137) (Fig. 4a). Salientamos que houve diferenças individuais, tais como sair rapidamente para uma nova busca mesmo sem ter obtido sucesso na viagem anterior, que não implicaram em modificações do comportamento em geral.
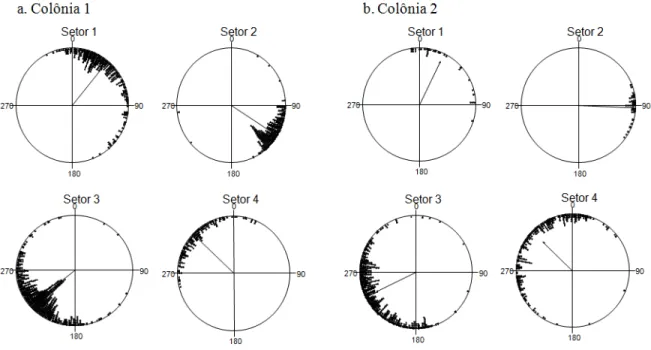
Figura 5. Padrão direcional das operárias das duas colônias (a e b), agrupadas por setores
de atividade. A seta central é o vetor médio (µ) com seu comprimento (r). (Colônia 1: Setor 1- µ=39,142°, r=0,841; Setor 2- µ=122,303°, r=0,951; Setor 3- µ=229,868°, r=0,876; Setor 4- µ=312,743°, r=0,903. Colônia 2: Setor 1- µ=25,773°, r=0,848; Setor 2- µ=91,926°, r=0,977; Setor 3- µ=243,693°, r=0,786; Setor 4- µ=313,778°, r=0,724.).
Cada operária manteve uma preferência direcional individual durante seu deslocamento, em todas as viagens. Todavia, mais de uma formiga fazia a cobertura de uma determinada área ao redor do ninho (Colônia 1: Setor 1- N=342, Z= 241.61, p<0.000; Setor 2- N=288, Z=260.39, p<0.000; Setor 3- N=625, Z=480.10, p<0.000; Setor 4- N=161, Z=131.41, p<0.000. Colônia 2: Setor 1- N=34, Z=24.45, p<0.000; Setor 2- N=42, Z=40.05, p<0.000; Setor 3- N=453, Z=280.12, p<0.000; Setor 4- N=265, Z=138.93, p<0.000). Da colônia 1 reunimos os pontos de 25 operárias, enquanto que da colônia 2 unimos os pontos de 16 delas, perfazendo 37% do total de operárias observadas durante o estudo (Fig. 5).
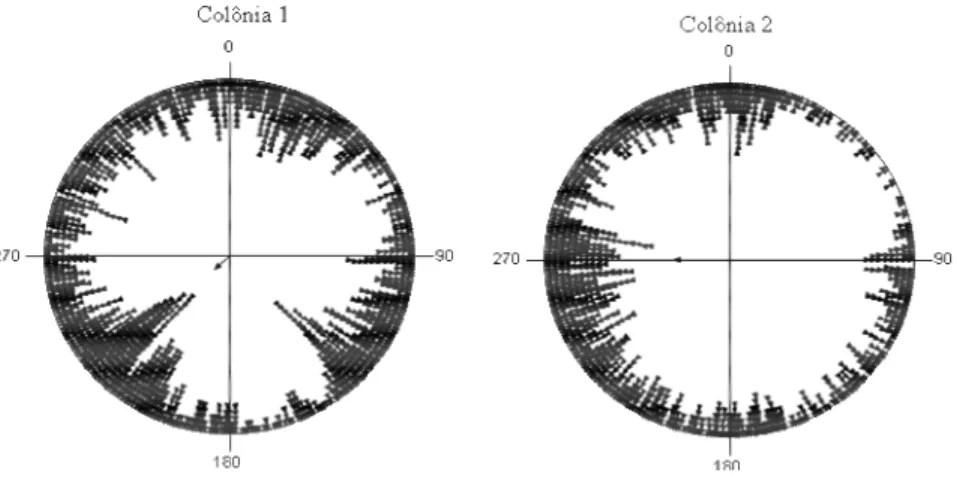
Quando analisamos no nível de colônia, verificamos que as colônias não apresentaram uma preferência direcional e sim uma distribuição espacial uniforme na área circundante aos ninhos (Colônia 1: N=1823, U2=1.29, p=0.005; Colônia 2: N=1336, U2=0.328, p=0.005) (Fig. 6).
Figura 6. Distribuição espacial das 109 operárias observadas (colônia 1: 62 operárias;
colônia 2: 47 operárias) ao redor de cada colônia, durante a atividade de forrageio. (Colônia 1: µ=230,468°, r=0,106; Colônia 2: µ=269,978°, r=0,299).
DISCUSSÃO
informações relevantes do ambiente pode induzir o forrageador a realizar ações para reduzir a incerteza, requerendo algum custo no tempo ou energia (Stephens, Brown e Ydenberg 2007). A recompensa obtida por um indivíduo através de qualquer estratégia de seleção de presa depende da estratégia adotada pelos seus competidores (Giraldeau e Caraco 2000). Para isso, os forrageadores devem decidir onde procurar, quanto tempo procurar num determinado local e se retornar ao local ou não de uma busca prévia (Traniello 1989).
As operárias de D. quadriceps neste estudo investiram a maior parte do tempo de permanência na área de uso da colônia na atividade de forrageio, como conseqüência do deslocamento a distâncias maiores. Ajustaram a decisão do tempo de forrageio e a distância percorrida de acordo com o tamanho do alimento capturado. É provável que itens alimentares maiores fossem exauridos, primeiramente, próximo do ninho, por ser uma área vasculhada por todas as operárias. Isso pode ter forçado as operárias a se deslocarem para distâncias cada vez maiores a fim de retornar ao ninho com alimento, mesmo que o sucesso não fosse garantido.
Todos os alimentos coletados por D. quadriceps foram sólidos, preferencialmente de origem animal conforme Araújo e Rodrigues (2006). Este tipo de recurso requer distâncias de forrageio maiores e sua busca, captura e transporte, em geral, consumindo mais tempo quando comparado a alimento líquido (Fourcassié e Oliveira 2002). De acordo com Bonser et al. (1998), no contexto do forrageio de ponto central, o tempo de permanência na área de uso dependerá da distância do ninho ao local onde há alimento. Os trechos que estão mais distantes do ninho devem ser explorados por um período mais longo do que os mais próximos do ninho.
escolha de uma presa apropriada para levar à colônia, se tamanho ou peso da presa ou mesmo um critério diferente. Todavia, parece que o sucesso da operária numa viagem é um estimulo para ela sair rapidamente do ninho e seguir em direção ao local da captura. Por outro lado, se o sucesso de uma operária motiva outra ou outras forrageadoras a sair ainda não está claro. No caso de carregadores de presa única, como D. quadriceps, que traz apenas um item alimentar por vez, os forrageadores devem estabelecer um tamanho mínimo de presa quando as presas são abundantes; quando viajar a longa distância do ninho para buscá-las e/ou capturá-las (Stephens, Brown e Ydenberg 2007).
O ajuste no esforço de forrageio de acordo com o tipo de recurso varia de uma espécie para outra. Em Lasius niger, o critério de avaliação da quantidade de alimento é fisiológico e tem base na habilidade da formiga para ingerir um volume requerido de honeydew (Mailleux, Deneubourg e Detrain 2000). Para Pheidole pallidula, o elemento principal de escolha é a resistência de tração da presa, que pode promover a mudança de busca individual a coletiva (Detrain e Deneubourg 1997). Na espécie Aphaenogaster
senilis, as operárias respondem a transportabilidade do alimento através da modulação do
recrutamento de companheiras de ninho (Cerdá et al. 2009). Forrageadores de Dorymyrmex
goetschi são capazes de transportar alimentos de peso equivalente a 3.5 vezes sua massa
corpórea (Torres-Contreras e Vásquez 2004). Cataglyphis iberica também limita o tamanho da presa equivalente ao seu próprio tamanho (Cerdá 1988). Já Pogonomyrmex barbatus tem a atividade de forrageio estimulada pelo retorno de forrageadores bem sucedidos, enquanto o retorno dos mal sucedidos não afeta o forrageio (Schafer, Holmes e Gordon 2006).
D. quadriceps mostrou evidências da necessidade de aumentar a distância
entre as variáveis, tempo de permanência fora do ninho e distância percorrida, indica a importância das mesmas nas decisões individuais, que traz conseqüências para a eficiência do trabalho da colônia. O forrageio individual é geralmente favorecido com o aumento da distância do alimento (Carroll e Janzen 1973), no entanto, a disponibilidade de alimento influencia a duração da busca. Quando ela é baixa, as operárias demoram a encontrar alimento e retornar (Schafer, Holmes e Gordon 2006).
Convencionalmente, as formigas se deslocam através de seu ambiente de várias maneiras conforme buscam, porém após a localização de uma presa, elas voltam aos seus ninhos em linha reta (Herbers e Choiniere 1996). Em certas ocasiões, a forrageadora de D.
quadriceps ingeriu o alimento capturado e continuou forrageando, até encontrar uma presa
maior do que a ingerida e então retornar ao ninho. Essas, em geral, foram operárias que passaram muito tempo fora do ninho. Observações de Gigantiops destructor, em ambiente natural e laboratório, demonstraram que durante o forrageio, as operárias podem comer uma ou duas presas antes de retornar para o ninho (Beugnon, Chagné e Dejean 2001). As operárias que ingerem o alimento, provavelmente, são forrageadoras experientes que de alguma maneira avaliam uma característica do recurso alimentar obtido, como tamanho ou peso, por exemplo, e decidem a viabilidade de retornar com o mesmo. A imprevisibilidade do recurso alimentar no ambiente de uma colônia pode requerer que o forrageio seja regulado através da mudança na experiência comportamental de operárias individuais para aumentar a eficiência (Traniello 1989).
Araújo e Rodrigues (2006) afirmaram que, em geral, quando uma operária de D.
quadriceps coleta alimento, ela retorna para o ninho em velocidade variável e deslocamento
percurso de volta ao ninho, porém, aparentemente não em função do tamanho do alimento. Provavelmente, o que influenciou a variação na velocidade foi a distância do local de captura ao ninho. Myrmica punctiventris, em ambiente natural, também retorna ao ninho ao longo de uma trajetória próxima a linear (Herbers e Choiniere 1996). Gigantiops destructor após coletar néctar retornam ao ninho de uma maneira preferivelmente direta, a partir de distâncias acima de 20 m (Beugnon, Chagné e Dejean 2001). Ectatomma ruidum, ao capturar uma presa durante a viagem de longo curso, retorna ao ninho com maior velocidade e a seguir, parte ao longo de uma trilha aperfeiçoada em sua direção preferida (Franz e Wcislo 2003).
Após o retorno das operárias de D. quadriceps ao ninho, o tempo transcorrido dentro do mesmo antes de uma nova excursão aparentemente sofreu interferência da viagem prévia. Isto se justifica uma vez que as forrageadoras que retornaram sem alimento, isto é, não obtiveram sucesso na busca anterior, demoraram mais para sair novamente. Já as formigas bem sucedidas ficaram menos tempo dentro do ninho, porém, não houve diferença marcante quanto à proporção do alimento obtido. Ao que parece, nessa espécie apenas retornar com alimento já é um bom indicativo para sair de novo, padrão já descrito para outra Ponerinae, Neoponera (=Pachycondyla) apicalis (Deneubourg et al. 1987).
As operárias de D. quadriceps bem sucedidas na viagem prévia, saíram em direção ao local onde a presa anterior foi capturada. Inicialmente, ao chegar à área, as forrageadoras faziam vários movimentos em circulo ao redor do local da captura com um pequeno aumento do diâmetro a cada circulo; passado certo tempo, elas começavam a ampliar a busca na sua área de atividade central. Desta maneira, as operárias de D. quadriceps apresentaram fidelidade direcional que já havia sido demonstrada nas forrageadoras de
procuram por alimento dentro de um setor restrito ao redor do ninho (Fourcassié e Oliveira 2002), o que pode indicar uma característica intrínseca do gênero. Em Cataglyphis bicolor, as operárias também exibem uma forte tendência a retornar para determinada direção do alimento, denominada de fidelidade de setor (Buchkremer e Reinhold 2008). Também
Neoponera (=Pachycondyla) apicalis retorna à zona de captura da presa (Deneubourg et al.
1987). A repetição desta ação induz memórias de longo prazo de um nível de atividade elevado e fidelidade espacial (Deneubourg et al. 1987).
As forrageadoras, ao viajarem na mesma direção, tendem a cobrir uma região em forma de leque a partir do ninho. A região pode ser irregular na forma e com freqüência torna-se alongada à medida que a área próxima do ninho se esgota (Gordon 1995). Segundo Fourcassié e Oliveira (2002), essa fidelidade espacial pode ser restringida pelo custo de estar perdida quando as operárias perambulam fora de seu setor familiar. Cada direção de forrageio não está associada com qualquer distribuição de recompensa, mas, com uma probabilidade de encontrar um item alimentar (Buchkremer e Reinhold 2008).
uma função da heterogeneidade do ambiente (Deneubourg et al. 1987). O modelo matemático de Harkness e Maroudas (1985) mostra que um simples processo de reforço espacial pode levar no nível individual a uma alta fidelidade espacial e no nível de colônia a uma divisão da área de forrageio entre as operárias.
Cada operária D. quadriceps mostrou-se especializada em forragear numa determinada área de atividade que pode aumentar com o tempo de vida favorecendo a eficiência de forrageio no nível da colônia. No entanto, aqui não analisamos a experiência individual. O processo de aprendizagem permite um indivíduo tornar-se menos randômico e mais especializado (Deneubourg et al. 1987), enquanto o padrão de busca da colônia se aproxima de um processo de difusão randômica, que pode ser uma solução eficiente para buscar alimento cuja distribuição é imprevisível (Traniello 1989). O aumento no sucesso do forrageio com o tempo de vida pode ser explicado tanto com viagens de forrageio mais longas no fim da vida da formiga, como, pelo aumento no tempo de busca porque sua chance de estar num setor de alta recompensa aumenta durante sua ocupação tornando o esforço extra no tempo de procura recompensado com uma maior probabilidade de sucesso no forrageio (Buchkremer e Reinhold 2008).
forrageador (Harrison et al. 1988). A regulação da atividade de forrageio somente através da aprendizagem pode ser abaixo do ótimo (Deneubourg et al. 1987).
As colônias de D. quadriceps distribuíram uniformemente ao redor do ninho o esforço de forrageio, através da fidelidade direcional das operárias. Mas, se a estratégia adotada pelas operárias individuais torna a atividade de forrageio eficiente, depende da interação das decisões individuais a respeito do tempo, distancia e direção. A especialização por aprendizagem, no nível individual em relação a tempo e espaço, parece uma solução apropriada para espécies que vivem num ambiente que varia durante o ano (Deneubourg et al. 1987). Todavia, o simples fato do forrageador ter memória e retornar ao mesmo local não garante que ele capture uma presa, porque à medida que aumenta o número de operárias que descobrem uma fonte de recurso, diminui o valor da mesma (Goss et al. 1989). Assim, as consequências da ação individual para a aptidão da colônia devem depender do número de outros indivíduos da colônia que estão empenhados em tarefas similares (Herbers e Choiniere 1996).
AGRADECIMENTOS
Agradecemos a Marconi C. Rodrigues pelo software desenvolvido para desenhar o caminho (Mopi). Altay A. L. Souza pela ajuda na estatística. M. F. Arruda e D. L. O. Azevedo-Guilhermino pela leitura crítica do manuscrito. A Universidade Federal do Rio Grande do Norte pelo apoio logístico. Ao Instituto Chico Mendes de Biodiversidade (ICMBio) pela licença para coleta (Licenças nº 10602-1 e 12547-1) e o apoio logístico na FLONA de Nisia Floresta/RN. Ao Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq) (Grant no 302012/2006-0 e 401738/2007-8), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) e Fundação de Apoio à Pesquisa do Estado do rio Grande do Norte (FAPERN) (PRONEX Proc. MCT-01.0009-00/2002) pela ajuda financeira.
REFERENCIAS BIBLIOGRAFICAS
Araújo, A. and Rodrigues, Z. 2006. Foraging Behavior of the queenless ant Dinoponera
quadriceps Santschi (Hymenoptera: Formicidae). Neotrop Entomol. 35:159-164.
Batschelet, E. 1981. Circular Statistics in Biology. London: Academic Press.
Beauchamp, G. 2005. Does group foraging promote efficient exploitation of resources?. Oikos. 111:403-407.
Beugnon, G, Chagné, P and Dejean, A. 2001. Colony structure and foraging behavior in the tropical formicine ant, Gigantiops destructor. Insectes Soc. 48:347-351.
Carroll, CR and Janzen, DH. 1973. Ecology of Foraging by Ants. Annu Rev Ecol Syst. 4:231-257.
Cerdá, X. Food collection by Cataglyphis iberica (EM.) (Hymenoptera, Formicidae). 1988. Ann Zool (Wars).30:515-525.
Cerdá, X, Angulo, E, Boulay, R and Lenoir, A. 2009. Individual and colletive foraging decisions: a field study of worker recruitment in the gypsy ant Aphaenogaster senilis. Behav Ecol Sociobiol. 63:551-562.
Charnov, EL. 1976. Optimal Foraging, the Marginal Value Theorem. Theor Popul Biol. 9:129-136.
Corbara, B, Fresneau, D, Lachaud, J-P, Leclerc, Y and Goodall, G. 1986. An automated photographic technique for behavioural investigations of social insects. Behav Process. 13:237-249.
Deneubourg, JL, Goss, S, Pasteels, JM, Fresneau, D and Lachaud, JP. 1987. Self-organization mechanisms in ant societies (II): learning in foraging and division of labor. Experientia Supplementum 54:177-196.
Detrain, C and Deneubourg, JL. 1997. Scavenging by Pheidole pallidula: a key for understanding decision-making systems in ants. Anim Behav. 53:537-547.
Fourcassié, V, Henriques, A and Fontella, C. 1999. Route fidelity and spatial orientation in the ant Dinoponera gigantea (Hymenoptera, Formicidae) in a primary forest: a preliminary study. Sociobiology. 34:505-524.
Fourcassié, V and Oliveira, PS. 2002. Foraging ecology of the giant Amazonian ant
Dinoponera gigantea (Hymenoptera, Formicidae, Ponerinae): activity schedule, diet and
Franz, NM and Wcislo, WT. 2003. Foraging behavior in two species of Ectatomma (Formicidae: Ponerinae): individual learning of orientation and timing. J Insect Behav. 16:381-410.
Giraldeau, LA and Caraco, T. 2000. Social Foraging Theory. Princeton: Princeton University Press.
Gordon, DM. 1995. The development of an ant colony’s foraging range. Anim Behav. 49:649-659.
Gordon, D. 2002. Formigas em ação: Como se organiza uma sociedade de insetos. Rio de Janeiro: Jorge Zahar.
Goss, S, Deneubourg, JL, Pasteels, JM and Josens, G. 1989. A model of noncooperative foraging in social insects. Am Nat. 134:273-287.
Harkness, RD and Maroudas, NG. 1985. Central place foraging by an ant (Cataglyphis
bicolor Fab.): a model of searching. Anim Behav. 33:916-928.
Harrison, JF, Fewell, JH, Stiller, TM and Breed, MD. 1988. Effects of experience on use of orientation cues in the giant tropical ant. Anim Behav. 37:869-871.
Herbers, JM and Choiniere, E.1996. Foraging behaviour and colony structure in ants. Anim Behav. 51:141-153.
Hölldobler, B and Wilson, EO. 1990. The Ants. Cambridge: Harvard University Press. Jander, R. 1975. Ecological Aspects of Spatial Orientation. Annu Rev Ecol Syst. 6:171-188.
Kempf, WW. 1971. A preliminary review of the Ponerinae ant genus Dinoponera Roger (Hymenoptera: Formicidae). Studia Entomologica, 14:369-394.
Mailleux, AC, Deneubourg, JL and Detrain, C. 2000. How do ants assess food volume? Anim Behav. 59:1061-1069.
Martin, P and Bateson, P. 1994. Measuring behaviour: an introductory guide. Cambridge: Cambridge University Press.
Santee, DP and Arruda, MF. 1994. The Nísia Floresta Common Marmoset Research Station. Neotropical Primates. 2:8-11.
Schafer, RJ, Holmes, S and Gordon, DM. 2006. Forager activation and food availability in harvester ants. Anim Behav. 71:815-822.
StatSoft, Inc. 2005. STATISTICA (data analysis software system), version 7.1. www.statsoft.com
Stephens, DW and Krebs, JR. 1986. Foraging economics: the logic of formal modeling. In: Foraging Theory. Princeton, New Jersey: Princeton University Press. p. 3-12.
Stephens, DW, Brown, JS and Ydenberg, RC. 2007. Foraging: Behavior and Ecology. Chicago: University of Chicago Press.
Torres-Contreras, H and Vásquez, RA. 2004. A field experiment on the influence of load transportation and patch distance on the locomotion velocity of Dorymyrmex goetschi (Hymenoptera, Formicidae). Insectes Soc. 51:265-270.
Torres- Contreras, H and Vásquez, RA. 2007. Spatial heterogeneity and nestmate encounters affect locomotion and foraging success in the ant Dorymyrmex goetschi. Ethology. 113:76-86.
Traniello, JFA. 1989. Foraging Strategies of Ants. Annu Rev Entomol. 34:191-210.
3.2ARTIGO 2º
TÍTULO: INTEGRAÇÃO DE INFORMAÇÕES AMBIENTAIS DURANTE O FORRAGEIO EM Dinoponera quadriceps (FORMICIDAE, PONERINAE)
AUTORES: AZEVEDO, D. L. O., MEDEIROS, J. C. & ARAÚJO, A.
Programa de Pós-graduação em Psicobiologia, Departamento de Fisiologia, Centro de Biociências, Universidade Federal do Rio Grande do Norte.
PERIÓDICO: INSECTES SOCIAUX
Qualis: A2 no Comitê Biológicas I da CAPES Fator de impacto: 1.391 (ano 2007)
INTEGRAÇÃO DE INFORMAÇÕES AMBIENTAIS DURANTE O FORRAGEIO EM DINOPONERA QUADRICEPS (FORMICIDAE, PONERINAE)
RESUMO
O comportamento de forrageio requer que os animais apresentem flexibilidade comportamental, para a expressão de estratégias eficientes que permitam o sucesso na busca por alimento. As formigas, durante o forrageio, apresentam estratégias de navegação que possibilitam viagens de longo curso e de retorno ao ninho, com o uso de pistas de orientação. Nós analisamos a flexibilidade no comportamento das operárias de Dinoponera
quadriceps na integração de informações introduzidas no ambiente. Observamos duas
colônias numa área de mata atlântica secundária e diante das entradas posicionamos um obstáculo físico a dois metros de distância, sem posição angular fixa em relação à entrada da colônia. Não houve diferença significativa quanto ao tempo de resolução das operárias para ultrapassar o obstáculo, após o primeiro contato elas continuaram desviando do mesmo, como qualquer outro obstáculo natural. A presença do obstáculo foi rapidamente incorporada pelas operárias não impedindo que elas mantivessem a direcionalidade das rotas para área de forrageio individual. O comportamento demonstrado pelas operárias de
D. quadriceps é uma indicação de que elas apresentam flexibilidade para aprender novas
informações do ambiente familiar.
Palavras-chave: Dinoponera quadriceps, forrageio, direcionalidade, integração de informação, aprendizagem.
INTRODUÇÃO
ações alternativas. O mecanismo envolvido em conseguir, estocar e lembrar esta informação econômica é a aprendizagem (Giraldeau e Caraco, 2000). Preferivelmente à perda de tempo numa busca randômica, os animais demonstram recorrer a dois sistemas de orientação: um sistema de rota associativa e um sistema de mapa cognitivo. Como a experiência espacial é geralmente seqüencial, esses sistemas, certamente, não são mutuamente exclusivos (Beugnon et al., 1996). Aprendizagem é a aquisição de representações neurais de nova informação. Discussão sobre aprendizagem, implicitamente, assume a existência de memória, que é a capacidade de reter a informação adquirida recentemente por no mínimo um curto período (memória de curto prazo), mas, frequentemente, durante períodos longos (memória de longo prazo) (Dukas, 2008).
Segundo Benhamou e Poucet (1996) a evolução dos processos de memória espacial está ligada ao surgimento da área de uso. Um indivíduo ganha familiaridade com uma área através da aprendizagem de pontos de referência e associando-os com a informação de navegação (Collett e Collett, 2000). A navegação local é o processo de movimento ao redor do ambiente imediato, em que apenas objetos dentro da extensão de percepção são úteis (Trullier et al., 1997).
ninho, com as coordenadas de locais significantes armazenadas na memória de longo prazo; a segunda são as memórias de pontos de referência ligadas a vetores locais que provêm os insetos com uma sequência de memórias direcionando-os ao longo de uma rota a partir do ponto de início até o objetivo (Collett e Collett, 2004).
Em formigas, o forrageio é estruturado para que elas permaneçam tão distante quanto possível dentro do território familiar (Gallistel, 1989; Collett e Collett, 2000). Uma fidelidade setorial preserva o investimento feito na aprendizagem de pontos de referência (Collett e Collett, 2000). Os pontos de referência indicam preferivelmente para formigas operárias que ação realizar, em vez de sua localização dentro de um sistema de coordenada global. Manter sistemas independentes pode ter desvantagem, mas significa que os erros de um sistema não se propagam dentro do outro (Collett e Collett, 2004).
As formigas operárias com múltiplas experiências em um local de alimentação não aprendem a distância, mas se familiarizam com as rotas (Narendra et al., 2007). Franz e Wcislo (2003) mostram que o efeito acumulativo de múltiplos sucessos imediatos no comportamento de forrageio de Ectatomma ruidum, indica que experiências passadas podem ser integradas durante intervalos longos e não em eventos simples. Formigas experientes de Paraponera clavata viajam mais rápido do que formigas inexperientes. É possível que elas usem outras pistas de orientação além das trilhas feromonais, provavelmente um sistema de orientação baseado nas pistas memorizadas de pontos de referência local (Harrison et al., 1988). Formigas Serrastruma lujae são provavelmente capazes de memorizar a localização de seu ninho pelo processo de informação baseado na rota durante os movimentos de busca superficial e local (Dejean e Benhamou, 1993).
presença de objetos cromaticamente bem camuflados (Shashar et al., 1998). A polarização da luz pode ser vista mesmo abaixo do dossel e pode ser usada por animais que vivem no solo. Dentro de uma floresta a folhagem densa limita a penetração de luz (Shashar et al., 1998), e a efetividade de um dossel como pista de orientação parece depender da conspicuidade do padrão visual (Hölldobler, 1980). A imagem de um objeto na retina muda com a distância e algumas vezes com a direção visualizada. Uma visão simples armazenada pode ser suficiente para guiar um inseto do ponto de início até o objetivo (Judd e Collett, 1998).
A interação contínua entre vetores locais e outras estratégias de navegação dão às rotas a flexibilidade para localizar o recurso (Collett e Collett, 2009). A flexibilidade do comportamento de forrageio demonstra que as formigas podem fazer ajustes complexos (Dussutour et al., 2009). Mesmo com a habilidade para usar integração da trilha e aprender vetores locais, a formiga de deserto Cataglyphis fortis não pode aprender ou reter rotas arbitrárias que são desprovidas por pontos de referência (Collett e Collett, 2009).
METODOLOGIA
Local de estudo
O estudo foi desenvolvido na Floresta Nacional de Nísia Floresta (FLONA) do Instituto Chico Mendes de Conservação e Biodiversidade (6°5’S, 35°12’W), localizada a 2 km da sede do município de Nísia Floresta, Estado do Rio Grande do Norte, Brasil. A estação de pesquisa tem uma área de 180 ha, dos quais 40 ha são de plantações de espécies exóticas e 80 ha de Mata Atlântica secundária. Nesta ultima área, as arvores podem atingir cerca de 20 m de altura, o solo é arenoso e a camada de húmus, em geral, tem aproximadamente 20 cm de profundidade (Santee e Arruda, 1994).
Sujeitos
Duas colônias de Dinoponera quadriceps Santschi (Hymenoptera, Formicidae, Ponerinae) foram escolhidas de forma aleatória, dentro da área de Mata Atlântica secundária. Elas localizavam-se 13,1 m de distância uma da outra. Nas duas colônias existiam mais de uma entrada (Colônia C - 4 entradas; Colônia A - 2 entradas), porém, apenas uma entrada era monitorada. O critério de escolha foi a ordem no qual encontramos as entradas, a primeira a ser encontrada, em ambas as colônias, tornou-se o ponto de partida das observações durante todo o estudo. As operárias foram marcadas individualmente e continuamente, com etiqueta numerada e plastificada, colada na região do tórax, utilizando uma cola à base de éster de cianoacrinolato (Corbara et al., 1986). Adotamos cores diferentes para as etiquetas de acordo com a colônia as quais pertenciam.
Observações
mínimo 10 horas/dia, pelo menos uma vez por semana, das 05h30min às 17h30min. Para as observações escolhemos a técnica do focal contínuo (Martin e Bateson, 1994), com registro do início e do termino do forrageio; também se o indivíduo retornou com ou sem alimento. As operárias foram observadas no mínimo duas vezes consecutivas por dia.
Registro do deslocamento das operárias
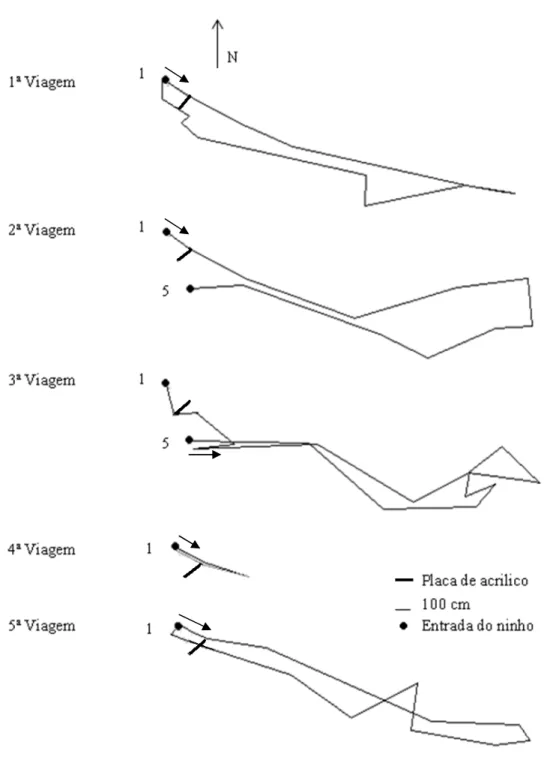
Para mapear os deslocamentos das formigas ajustamos a técnica descrita por Turchin et al. (1991). Assim, durante o forrageio, a cada cinco minutos assentávamos uma bandeirola numerada, e independente deste tempo fixo, também assentamos bandeirolas no momento que a formiga se aproximava e/ou ultrapassava uma placa de acrílico. Ao final das observações mediamos a distância e o ângulo entre as bandeirolas numeradas, usando bussola e trena. Estes dados foram levados ao Mopi, um software desenvolvido exclusivamente para desenhar o deslocamento realizado pelas operárias, obtendo uma estimativa do caminho real. A partir deste desenho estimado, verificamos o posicionamento de cada bandeirola em relação ao ninho (Azevedo et al. submetido).
Procedimento experimental
desde a saída do ninho até a ultrapassagem do obstáculo. Registramos ainda a direção que a operária tomava após contornar a placa conforme descrito acima.
Analise estatística
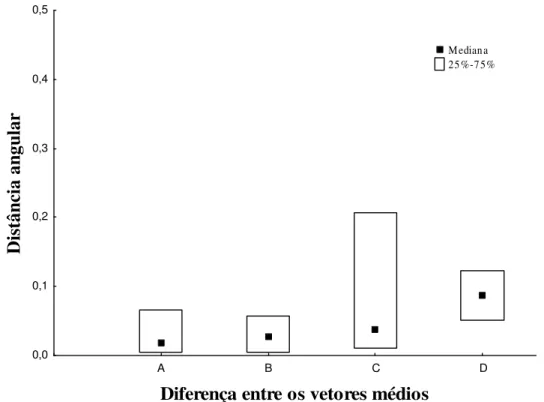
Com os ângulos obtidos a partir do posicionamento das bandeirolas em relação ao ninho realizamos o teste de Rayleigh, utilizando o software Oriana 2.01 (Kovach, 2004). Obtive o vetor médio de cada viagem e calculamos a diferença angular entre os vetores (Batschelet, 1981) das viagens nas quais a operária atingia e ultrapassava a placa. Se uma operária fosse observada em seis viagens diferentes, só consideramos na nossa analise as viagens nas quais ela manteve contato com a placa de acrílico. Todas as diferenças angulares foram feitas referente ao 1º contato, pois este era o momento que a operária entrava em contato com a nova informação do ambiente imediato ao ninho. Analisamos as diferenças em relação ao tempo de resolução e à diferença entre ângulos com o teste de Kruskal-Wallis utilizando o software Statistica 7.1 (StatSoft, 2005). O tamanho das amostras dos testes variou conforme as variáveis analisadas.
RESULTADOS
Tabela 1 Resultado das mudanças ocorridas em três variáveis do forrageio nas 316 viagens. Quartil
Variáveis de forrageio Mediana
25% - 75% Extensão Latência de saída (min) 4.0 2.0 – 13.0 0.0 – 217.0 Tempo de viagem (min) 16.5 4.0 – 48.3 1.0 – 337.0 Distância percorrida (cm) 966.0 99.8 – 2817.3 0.0 – 17290.0
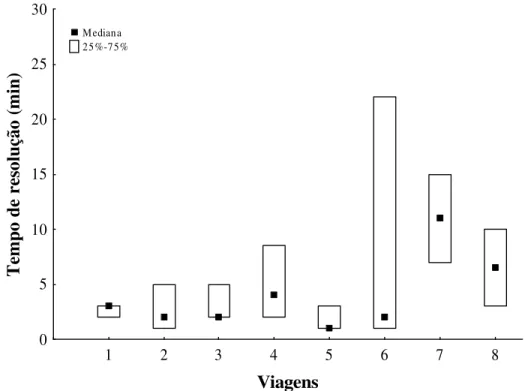
O tempo de resolução, que era o tempo transcorrido desde a saída do ninho até a ultrapassagem da placa de acrílico, não apresentou uma diferença significativa entre as viagens (N=140) (Kruskal-Wallis: H=11.9, df=7, p=0.1) (Fig. 1). O tamanho da amostra representa 44,3% do total de viagens observadas durante o estudo e foi composta pelas viagens realizadas por 47 operárias diferentes, uma vez que este foi o numero de operárias que ultrapassaram a placa de acrílico.
1 2 3 4 5 6 7 8
Viagens 0 5 10 15 20 25 30 T em po d e re so lu çã o (m in ) Mediana 25%-75%
Fig. 1 Diferença entre os tempos de resolução nas 140 viagens nas quais as operárias
Fig. 2 Representação gráfica das viagens da operária 61.