Estudo funcional do gene que codifica um transportador de membrana MFS em Colletotrichum lindemuthianum
Texto
Imagem

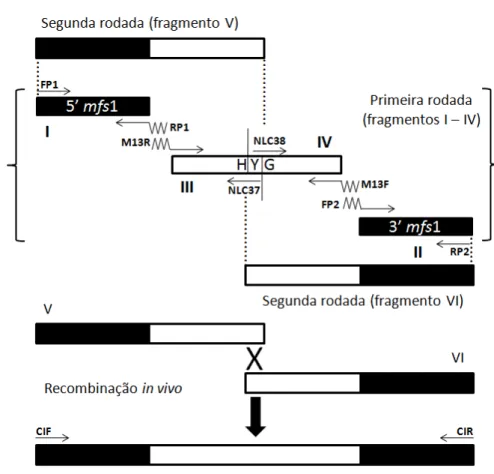


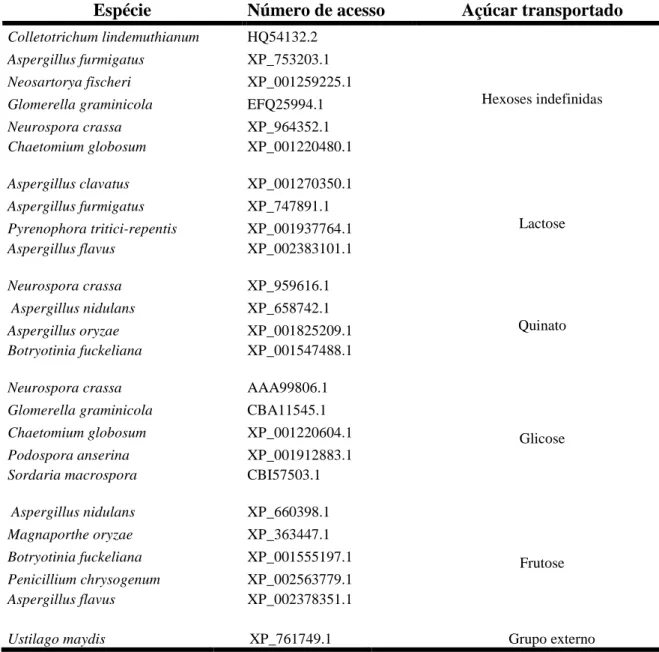
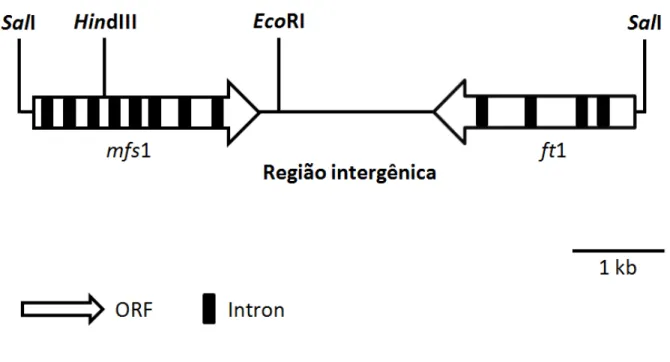

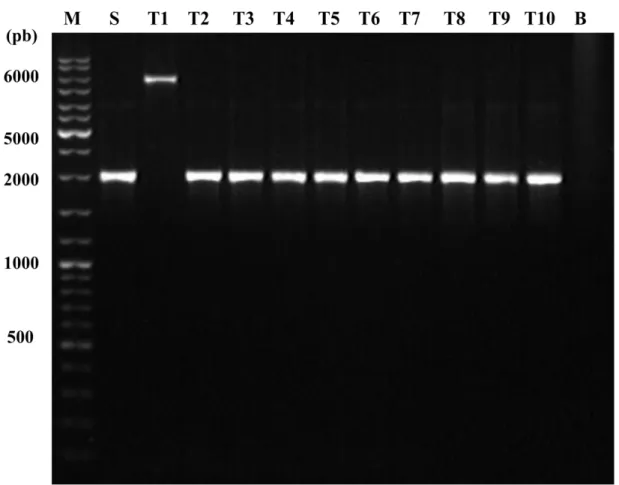
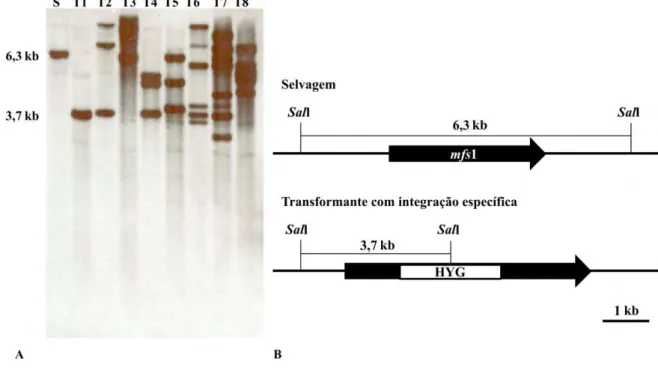
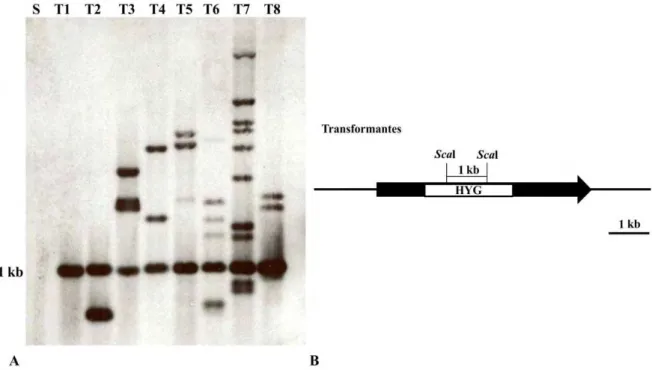
Documentos relacionados
E o efeito da escolarização está relacionado ao empenho da escola no combate às variantes: aquelas que são mais alvo de estigma, aquelas que demarcam frontei- ras entre
a) Institutionally grounded: The institutional dimension of conventional media and telecommunication operators heavily influences the conception of mobile media as
Expressed sequence tag analysis of the human pathogen Paracoccidioides brasiliensis yeast phase: Identification of putative homologues of Candida albicans
The purpose of this work was: (1) to model fluid flow in a tubular continuous processing system by application of the RTD Danckwerts analysis, (2) to assess
RELAÇÃO VAGAS/CURSO POR ÁREA ESPECÍFICA 30,0 85,9 89,1 83,1 62,0 140,2 122,2 125,0 50,0 77,7 100,4 107,6 60,0 91,7 114,2 107,5 112,0 123,2 105,0 Administração de redes Análise
O presente projeto visou a clonagem, expressão, purificação e caracterização cinética e biofísica da enzima fumarato hidratase codificada pelo gene LmjF24.0320 de
A perspectiva de uma luta de longa duração para erradicar as raízes do fascismo e o projeto de construção de uma via nacional, democrática e parlamentar para o socialismo, não
A análise se deu pelo método da Avaliação Qualitativa das Preparações do Cardápio (AQCP), o qual identifica a frequência absoluta e relativa da oferta diária
