Effects of landscape structure and Eucalyptus genotype on the abundance and biological control of Glycaspis brimblecombei Moore (Hemiptera: Psyllidae).
Texto
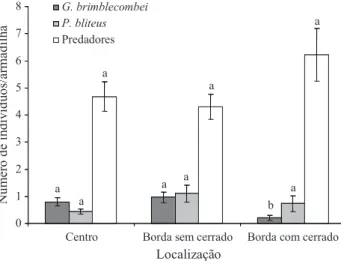
Imagem

Documentos relacionados
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
We present a hybrid method to numerically solve the inverse acoustic sound- soft obstacle scattering problem in R 3 , given the far-field pattern for one incident direction..
Na entrevista a seguir, Capovilla revisita a própria trajetória intelectual, debate a sua passagem pelo Centro Popular de Cultura de São Paulo e as críticas que escreveu para
It has been shown that non-linear FEA is capable of predicting with accuracy not only the curvatures developed during the curing of these laminates but also the snap-through loads
Procurámos, com a análise das características do terreno e das condições meteorológicas, determinar os seus efeitos na manobra dos exércitos de Massena e Wellington, que escolheram
“desvelamento da realidade”, a problematização, a desmistificação de mitos, construídos historicamente, sobre as interações entre Ciência-Tecnologia-Sociedade (CTS),
Tabela 2 – Biomassa seca g vaso-1 da parte aérea de plantas de variedades de caupi, inoculadas com diferentes estirpes ou na ausência de inoculação testemunhas, com e sem aplicação
O indivíduo, assim, deixar-se-ia atravessar por essa relação entre educação e projeto social como em uma via de mão dupla: por um lado torna-se “objeto” da
