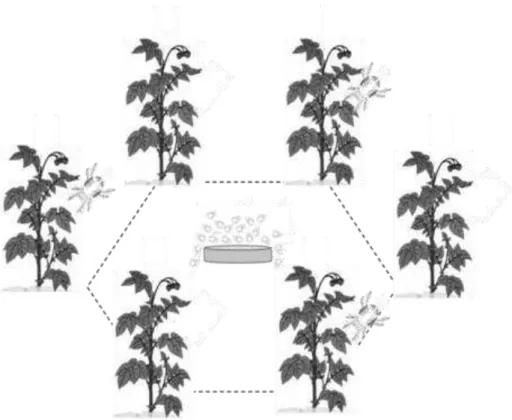
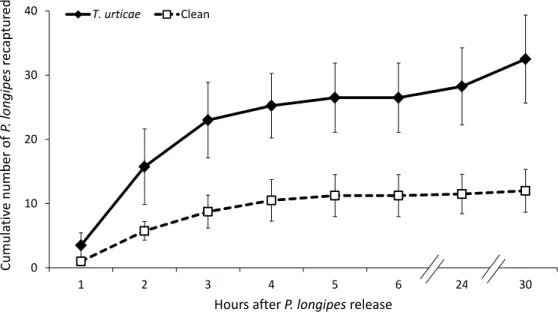
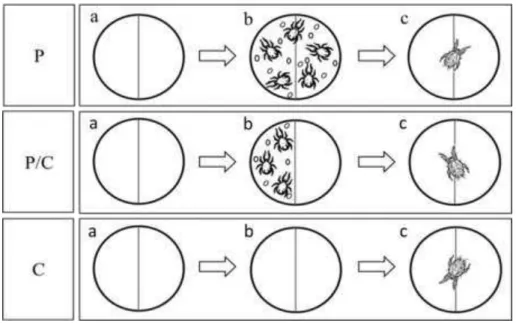
Foraging and antipredator behaviour in an acarine predator-prey system on tomato
Texto
Imagem







Documentos relacionados
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
HTDCs and pre-Ost cells laden in met-CS MAGPL hydrogels evidenced a significant decrease in cell content from day 1 to day 7 (p<0.05), independently of the EMF stimulation or
Os modelos desenvolvidos por Kable & Jeffcry (19RO), Skilakakis (1981) c Milgroom & Fry (19RR), ('onfirmam o resultado obtido, visto que, quanto maior a cfiráda do
Tendo em vista a crescente necessidade de novos modelos para restauração florestal, o presente estudo teve o objetivo de avaliar se alta diversidade de espécies vegetais, funções
Muito embora, a tecnologia de marcadores moleculares seja uma estratégia importante para auxiliar no processo de seleção de novos genótipos, a fenotipagem dos
O processo ocorre como na experiência do bailarino, que sente a energia fluir através dos membros, ao mesmo tempo em que é acompanhado pelo movimento de sua consciência,
Our results indicate that the choline-based ILs were effective functional ingredients, since, when used at non-toxic concentrations, they allowed a higher drug loading, in topical
Os biomarcadores do álcool podem ser divididos em dois principais grupos: aqueles usados para a detecção de consumo crônico (marcadores de longo prazo) e aqueles usados para
