Hereditas 133: 73-79 (2000)
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Synaptomenal complex analysis
of
four breeds of
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Bos taurus taurus
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
x
B.
taurus
indicus hybrids
MARCIA D. P. SCAVONE', CLAUD10 OLIVEIRA', LUZIA APARECIDA TRINCA2 and FAUSTO FORESTI'
I
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Departamento de Morfologia, Instituto de Biocigncias, Universidade Estadual Paulista, Botucatu, SP, BrazilDepartamento de Bioestatistica, Instituto de Biocigncias, Universidade Estadual Paulista, Botucatu, S P , Brazil
Scavone, M. D. P., Oliveira,
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
C., Trinca, L. A. and Foresti, F. 2000. Synaptonemal complex analysis of four breeds of Bos taums taurus x B. taurus indicus hybrids.-Hereditas 133: 73-79. Lund, Sweden. ISSN 0018-0661. Received May 3, 2000.Accepted October 3, 2000
The synaptonemal complex (SC) was analyzed in four F, hybrids of Bos taurus taurus and B. faurus zndicus including Gyr-Simmental (G-S), Nelore Simmental (N-S), Gyr-Holstein-Friesian (G-H) and Nelore-Piemontese (N-P). We analysed the frequency of various types of SC abnormalities and the frequency of cells with SC abnormalities. The results were compared with similar observations made on purebred animals. All the animals studied possessed 29 autosomal and one
sex bivalent. The frequency of cells with abnormalities in the hybrids were 28.0 Yn
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
in the N-P, 29.1 %) in the G-S, 33.3 %in the N-S and 40.0 Yn in the G-H. The frequency of cells with abnormalities in the four hybrids was 31.5 %I; 57.9 Yn of
these abnormalities occurred in zygotene and 42.0 Yi
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
occurred in pachytene. The comparisons among the hybrids and among the hybrids and their parental breeds showed that the only significant difference was between Gyr andGyr-Holstein-Friesian animals. Some aspects of the relationship between the frequency of cells with anomalies and the fertility of hybrids are discussed.
C. Oliveira, Departamento de Morjologia, Instituto de Biociincias, Universidade Estadual Paulista, Curnpus de Botucatu,
18618-000, Botucutu, SP, Brazil. E-mail: [email protected]
The hybrid animals produced by cross breeding of
Bos taurus taurus and B. taurus indicus are considered
superior to the pure bred amimals (ALENCAR 1997). The hybrids are more efficient producers of meat and milk than the parental breeds (ALENCAR 1997), and their fertility rates are generally lower than 80% (PEREIRA NETO and LOBATO 1998).
Although many factors can affect the reproductive efficiency in animals, the process of chromosome pairing in the first meiotic prophase is crucial in the production of balanced haploid gametes (GILLIES
1989). The pairing of the chromosomes is mediated by the synaptonemal complex (SC), a protein struc- ture formed in the beginning of the prophase I be- tween the sister chromatids of each bivalent (SCHMEKEL et al. 1993). Techniques have been devel- oped to study the morphology of the SC (COUNCE
and MEYER 1973; SANTOS 1993). They allow a de- tailed analysis of the process of chromosome pairing. We have studied the fertility problems in hybrids between B. taurus taurus and B. taurus indicus, through using the SC of four F, hybrid breeds in order to determine details of chromosome pairing. We compare these results to similar data obtained from pure blood animals of their respective parental breeds.
MATERIAL AND METHODS
Animals
Testes of 18 specimens of F, hybrids from Nelore and Piemontese breeds (N-P) and 10 of F, hybrids from Nelore and Simmental (N-S) were obtained in Estrela do Oeste, State of S5o Paulo, Brazil. Testes of 3 F,
hybrids from the Gyr and Holstein-Friesian (G-H) breeds and of 3 F, hybrids from the Gyr and Sim- mental breeds (G-S) were obtained in Itatinga, State of S5o Paulo, Brazil.
Spread preparation
The SC was analyzed by surface spreading (SANTOS
1993). Briefly, the spermatocytes were lysed with
Triton XlOO (0.01
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
YO),
fixed in paraformaldehyde74
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
M . D. P. Scavone et al. Hereditas 133 (2000)Class$cation
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
of the cellsIn the analysis of the different stages of cellular development, two zygotene sub-stages were recog- nized corresponding to early and late stages. In the pachytene, three sub-stages (early, middle and late) were recognized based on the degree of pairing of the bivalent, the reduction of the size of the axes, conden- sation of the sexual vesicle and dispersion of the
nucleoli in agreement with the proposal of GREEN-
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
BAUM et al. (1986).
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
A nafyses
For each cell analyzed, the number of the following parameters was registered: 1. intimate connection be- tween the XY axes and autosomes (XY-A); 2. lateral elements with unequal length (UL); 3. Y chromosome broken (YB); 4. association of two telomeres of a SC
(bivalent in arch) (EE); 5. telomere association (TA); 6, univalent (U); 7. inversion (I); 8. break (B); 9. pairing failure (PF); 10. unpaired parts (UP); 11. twister (T); and 12. telomere concentration (TC). Cells with the characteristics 1 to 10 were counted as carriers of abnormalities. The parameter P F (9) was only considered when the cells were not in late pachytene because in that case it was not possible to distinguish between pairing failures and failures due the occurrence of desynapse.
The Kruskal-Wallis test (ZAR 1996) was employed
to analyze the data. Data regarding the proportions of cells possessing abnormalities were statistically an- alyzed by the Fisher exact test (ZAR 1996).
RESULTS
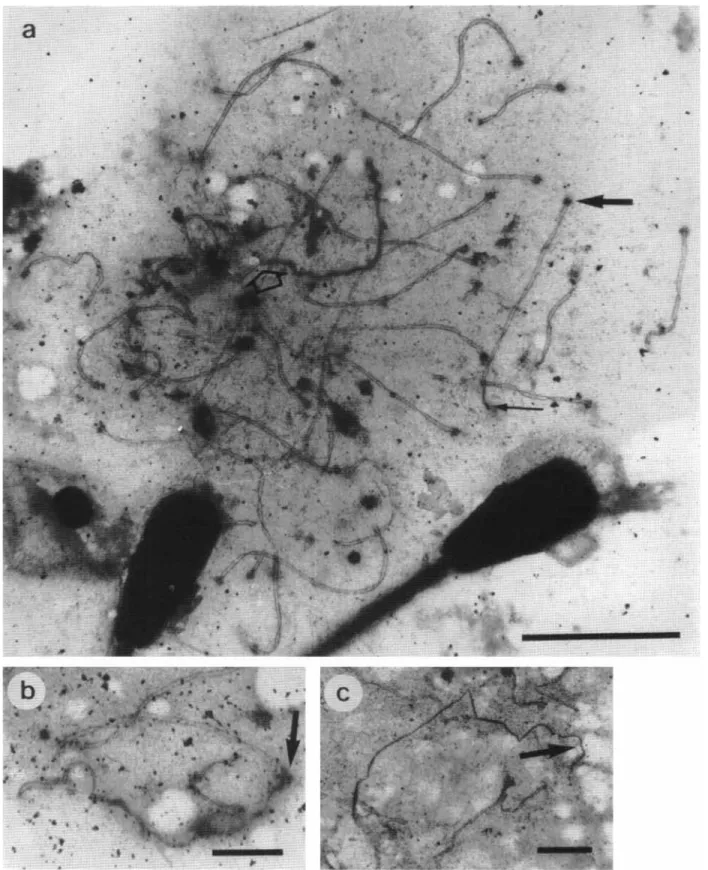
All animals had 29 autosome bivalents and one sex- ual bivalent (Fig. la). The axes of these bivalents had a dense cloud of protein material representing the kinetochore and a terminal plaque in the other ex- tremity (Fig. la). The autosomic bivalents were acro- centric with the pairing beginning in the telomeric region of the long arm and progressing unidirection- ally (Fig. 2a). The distance between the lateral ele- ments was of 100 nm and the average of absolute length of SC in cells in middle pachytene was 180 pm. The behavior of SC in zygotene and pachytene (Fig. la) was similar those described by GREENBAUM
et al. (1986). In the early pachytene, the autosomes were completely paired and the XY chromosomes were completely heteropycnotic. In the middle pachytene, the unpaired region of the X chromosome remained heteropycnotic (Fig. 1 a). Frequently, the Y
chromosome was broken during the pachytene (Fig. Ic).
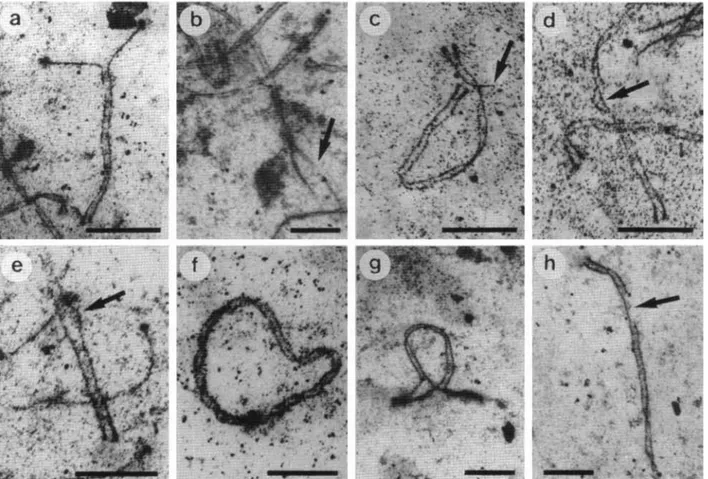
The figures show the following examples: associa- tions XY-autosomes (Fig. lb), lateral elements with unequal length (Fig. 2c, 2d and 2e), broken Y chro- mosome (Fig. lc), association of two telomeres of a same SC (Fig. 20, inversion (Fig. 2g), break (fig. 2h), pairing failure (Fig. 2b) and twister (Fig. la). On Table 1, the totals for each parameter in zygotene and in pachytene cells are shown. The twisteds and telomere concentration were not counted as abnor- malities in spite of having been listed in the Table 1 because they are normal configurations in the phases in which they occurred. Despite a number of statisti- cally significant differences in several of the parame- ters analyzed in zygotene and pachytene sub-stages, the total of all the abnormalities did not show a
statistically significant difference among the hybrids
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
(x’
=zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
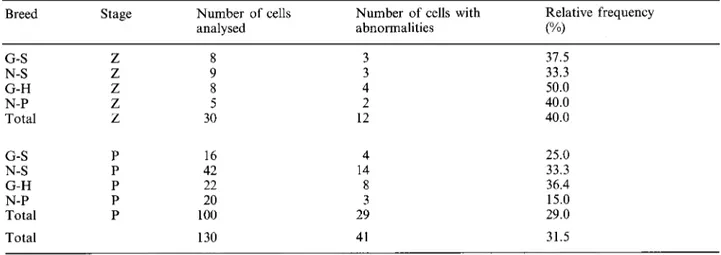
3.940, df = 3, p = 0.268). The percentages of cells with abnormalities in thezygotene and pachytene stages are presented in the Table 2. The total percentage of cells in F, hybrids with abnormalities in the Gyr-Simmental was 29.1 %,
in the Nelore-Simmental there was 33.3 %I, in the
Gyr-Holstein-Friesian was 40.0 9’0
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
and in the Nelore-Piemontese was 28.0
YO.
The total percentage of cells with abnormalities for all the hybrids was 31.5 YO(Table 2) with 57.97 % of abnormalities occurring in the zygotene stage and 42.03% in the pachytene stage.
Statistically significant differences were not found in the total number of abnormalities observed among the Gyr-Simmental hybrids and the parental average
(x2
= 4.637, df = 2, p = 0.098). A comparison of thefrequency of cells with abnormalities between Gyr-Si- mental F, hybrids (29.1 940) and the parental Gyr (1 1.1 %) and Simmental (29.5
YO)
breeds revealed statistically significant differences only between the hybrids and pure bred Gyr animals (p=O.O05).For the Nelore-Simmental hybrids, statistically sig- nificant differences were found in the total number of
abnormalities observed between the hybrids and the parental average
(x’
= 15.609, df = 2, p E 0.000). A comparison of the frequency of cells with abnormali- ties between Nelore-Simmental F1 hybrids (33.3 %)and the parental Nelore (30.0%) and Simmental (29.5 YO) breeds did not indicate any statistically sig- nificant differences between them (p = 1 .OOO; p =
0.825 respectively).
The results for the total number of the abnormali- ties indicated that there were only statistically signifi- cant differences between the Gyr-Holstein-Friesian and the parental Gyr breed
(x’=
8.215, df = 2, p =Hereditas 133
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
(2000) Synaptonemal complex in hybrid bullszyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
1 5Fig. la-c.
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
azyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
The synaptonemal complex of F, hybrids between Nelore-Simmental (2nzyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
= 60)-middle pachytene. The76
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
M. D. P. Scavone et al.zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Hereditas 133 (2000)zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
Fig. 2a-h. a A bivalent partially paired in late zygotene of an F, hybrid animal of Gyr-Holstein-Friesian showing that the
pairing begins in the long arm and proceeds in the direction to the centromere.
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
b A bivalent with pairing failure (arrow)of a F, hybrid Gyr-Holstein-Friesian.
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
c A bivalent with interstitial asymmetry (arrow) of F, hybrid between Nelore-Piemon-tese. d A bivalent with interstitial asymmetry (arrow) of a F, hybrid Nelore-Piemontese. e A bivalent with interstitial
asymmetry (arrow) of a F, hybrid between Nelore-Piemontese. f Association of the two telomeres of an autosome with a SC (bivalent in arch) in a F, hybrids between Gyr-Holstein-Friesian. g A bivalent with inversion in F , hybrid of
Gyr-Holstein-Friesian. h Bivalent with a break (arrow) in F , hybrid between Gyr-Holstein-Friesian. Bars
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
= 1 pm.Table 1. Number
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
of observations of the analyzed parameters of Fl hybrids among breeds of Bos taurus taurus andB. taurus indicus: Gyr and Simmental (G-S), Nelore and Simmental (N-S), Gyr and Holstem-Friesian (G-H), Nelore and Piemontese (N-P). Z-zygotene, P-pachytene, XY-A-intimate connection between the axes X Y and autosomes, UL-unequal length; YB- Y chromosome broken, EE-association of two telomeres of a SC (bivalent in arch); TA- telomere associations; U-univalent, I-inversion; B-break, PF-pairing failure, UP-unpaired part, T- twisted and TC- telomeres concentration
Breed Stage Number of cells Number of observations
XY-A UL YB EE TA U I B P F UP T TC
G-S Z
N-S Z
G-H Z
N-P Z G-S P
N-S P
G-H P
N-P P
8
9
8
5
16 42 22 20
0 0 0 0
3 14 10 0
20
0
21
0
15 42 19 17
0 0 0
0 0 0
0 0 0
0 0 0
1 0 0
10 11 77 12 11 77
0 1 19
21 1 0 3 0 0 24 1 0 12 1 0
0 0 0
3 0 10 63 0 10
0 0 1
10 45 20 91 30 137 37 47
197 2 320 7 691 331 230 10
59 0 62 0
64 3 25 0
602 16
Hereditas
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
133zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
(2000) Synaptonemal complex in hybrid bulls 77Table 2. Frequency
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
of cells with abnormalities in hybrid Fl bulls between the Gyr and Simmental (G-S), Neloreand Simmental (N-S), Gyr and Holstein-Friesian (G-H), Nelore and Piemontese (N-P) breeds. Z-zygotene and P-pachytene
Breed Stage Number of cells Number of cells with Relative frequency
analysed abnormalities
("/I
G-S N-S G-H N-P Total G-S N-S G-H N-P Total Total Z Z Z Z Z P P P P P 8 9 8 5 30 16 42 22 20 100 130 3 3 4 2 12 4 14 8 3 29 41 37.5 33.3 50.0 40.0 40.0 25.0 33.3 36.4 15.0 29.0 31.5
cally significant differences between Holstein-Friesian
and Gyr-Holstein-Friesian (p =
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
0.540) but there wasa significant difference between Gyr and Gyr-Hol- stein-Friesian (p = 0.005).
Statistically significant differences were not found in the total number of abnormalities observed be- tween the Nelore-Piemontese hybrids and the
parental average
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
(x'
= 0.882, df = 2, p = 0.643). Acomparison of the frequency of cells with abnormali- ties between Nelore-Piemontese hybrids (20.0 %) and the parental Nelore (30.0 YO) and Piemontese (30.0 YO)
breeds did not indicate any statistically significant differences between them (p = 0.5001; p = 0.869 respectively).
A comparison between the number of cells with abnormalities between the different hybrids indicated that there were not statistically significant differences amongst them.
DISCUSSION
The synaptonemal complex of the hybrids between B. taurus taurus and B. taurus indicus showed 30 biva-
lents, 29 being acrocentric autosomal bivalents and one sex bivalent as had been previously observed by
DOLLIN et al. (1991b) in other bovine hybrids. The distance between the lateral elements in the ho- mologous pair was 100 nm; a result similar to other species of mammals (GILLIES 1989). The pairing of the chromosomes began in the telomeric region of the long arm and progressed unidirectionally as observed by GREENBAUM et al. (1986) for other mammals and by DOLLIN et al. (1989, 1991a,b) for bovines.
The most frequent abnormality observed in the hybrids was pairing failure. This abnormality was also found in high frequency in the hybrids studied
by DOLLIN et al. (1991b). Lateral elements with unequal length found in several F, hybrids had also been observed in the deer mouse (HALE and GREEN-
BAUM 1988), other bovines (DOLLIN et al. 1989; SWITONSKI et al. 1990; DOLLIN et al. 1991a,b) and buffaloes (DAI et al. 1994). The pairing failures and the asymmetric bivalents may have originated through heterochromatin polymorphisms, chromoso- mal rearrangements or disturbances in the pairing mechanism (SHARP 1986; HALE and GREENBAUM
1988; SUDMAN et al. 1989; SWITONSKI et al. 1990). The presence of some cells with inversions in the hybrids examined in this study show that these rear- rangements were present in some groups of cells of the germ line.
Breaks were observed in the Y chromosome in 23 Yn of the analyzed hybrids. The presence of these breaks in the Y chromosome of cattle has been described in pure bred bulls (DOLLIN et al. 1989), hybrids (SWITONSKI et al. 1990; DOLLIN et al. 1991a,b) and in buffaloes (DAI et al. 1994). Since the X chromosome is about three times longer than the Y chromosome and the telomenic plaques are attached to the nuclear envelope, it is possible that the forces exerted to move the terminal areas of the X and Y chromosomes could potentially break the Y chromo- some during SC preparations. DOLLIN et al. (1989) suggested that the frequently observed Y chromo- somes breakage in cattle might indicate that a weak point exists in the Y chromosome causing it to be highly susceptible to breakage during the preparation of the cells for electron microscopy.
18
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
M . D.zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
P.
Scavone et al. Hereditas 133 (2000)justment occurred in the cells to repair the anomalies, even though the mechanism is not completely accu- rate. This is shown by the presence of abnormal configurations in late pachytene cells. Similar obser- vations have also been made on bulls (DOLLIN et al.
1991a,b) and buffaloes (DAI et al. 1994).
Since the abnormalities observed in the hybrids always occurred in a reduced number of cells it may be supposed that a genetic unbalance, more than specific differences in the chromosomes, could be responsible for the abnormalities observed in the hybrid cells, as proposed by DOLLIN et al. 1991a,b). On the other hand, gross pairing abnormalities such as the extensive pairing failures observed in sterile hybrid horses (CHANDLEY et al. 1974) and foxes (GUSTAVSSON et al. 1988) were not observed in the hybrids studied in the present paper.
The comparisons between the number of cells with abnormalities in the hybrids showed that there were
not significant differences among them.
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
A compari-son of the frequency of cells with abnormalities in the
F, hybrids (31.5 YO)
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
and the parental breeds of B.taurus taurus (29.6 YO) and B. taurus indicus (16.9 YO)
(SCAVONE et al., in preparation) showed that there was greater similarity between the hybrids and the European bulls. On the other hand, the total of all the abnormalities in all the phases showed that there were only significant differences between the hybrids Nelore-Sinimental and their parental breeds and be- tween the Gyr-Holstein-Friesian hybrids and Gyr.
DOLLIN et al. (1991a) analyzing bovine hybrids between Brahman and Hereford, Sahiwal and Here- ford, Sahiwal and Shorthorn and between Guzerat
and Hereford found 23
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
% of cells with abnormalities.We observed that the hybrids had 31.5 YO of cells with abnormalities. A statistical comparison between the
data of DOLLIN et al. (1991a) and the present results indicates that there is a statistically significant differ- ence between them (p = 0.021).
The smaller frequency of anomalies in B. taurus
taurus and
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
B. taurus indicus than those observed inthe hybrids contrasts with observations that indicate the hybrid animals are more fertile than the pure breeds (TREMATORE 1996; ALENCAR 1997). To ex- plain why the hybrids have a larger number of cells with chromosome abnormalities than the pure breeds but are still more fertile, it is reasonable to suggest that the greater fertility may be related to some beneficial effects of heterosis not detected in our SC analysis. Considering that the occurrence of synapsis adjustment could minimize the importance of the SC
anomalies, the higher incidence of SC anomalies in
hybrids could be minimized by the positive effect of heterosis. It would be desirable to study a higher number of animals and the fertility rate in relation to
the occurrence of abnormalities of SC in order to discuss the causes of fertility problems in bulls of F, hybrids between Bos taurus taurus and B. taurus indicus.
ACKNOWLEDGEMENTS
The authors are grateful to R. Devide for technical assis- tance, to Dr. E.A. Gregorio for making available the Electron Microscope Unit, to Dr. A.A. Ramos for several important suggestions and to Dr. S. Kasahara, Dr. J.F. Lui, Dr. L.L.S. Mota and Dr. M.K. Tansey for a critical review of the original manuscript. Funds supporting this study were provided by CNPq and FAPESP.
REFERENCES
Alencar MM, (1997). Los cruzamientos para la produccion
de carne bovina. In: 5Q Congreso Internacional de Transferencia Tecnologica Agropecuaria, Asuncion CEA, Asuncion, p. 111-122.
Chandley AC, Jones RC, Dott HM, Allen WR and Short
RV, (1974). Meiosis in interspecific equine hybrids. I. The male mule (Eqqus asinus x E. caballus) and hinny (E. caballus x E. asinus). Cytogenet. Cell Genet. 13: 330-341.
Counce SJ and Meyer GF, (1973). Differentiation of the synaptonemal complex and the kinetochore in Locusta, spermatocytes studied by whole mount electron mi- croscopy. Chromosoma 44: 23 1-253.
Dai K, Gillies AE, Dollin AE and Hihr M , (1994). Synap-
tonemal complex analysis of hybrids and purebred water buffaloes-(Bubalus bubalis). Hereditas 121: 171- 1994. Dollin AE, Murray JD and Gillies CB, (1989). Synaptone- ma1 complex analysis of hybrid cattle. I. Pachytene substaging and the normal full bloods. Genome 32: 856-864.
Dollin AE, Murray JD and Gillies CB, (1991a). Synaptone-
ma1 complex analysis of hybrid cattle.
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
11. Bos indicus xBos taurus FI and backcross hybrids. Genome 34: 220- 227.
Dollin AE, Murray JD and Gillies CB, (1991b). Synaptone- ma1 complex analysis of hybrid cattle. 111. Meiotic pair- ing mechanisms in F1 Brahman x Hereford hybrids. Genome 34: 228-235.
Gillies CB, (1989). Fertility and Chromosome Pairing: Re- cent Studies in Plants and Animals. Boca Raton, CRC Press.
Greenbaum IF, Hale DW and Fuxa KP, (1986). The mechanism of autosomal synapsis and the substaging of
zygonema and pachynema from deer mouse spermato- cytes. Chromosoma 93: 203-212.
Gustavsson I, Switonski M, Larsson K and Ploen L, (1988). Synaptonemal analysis of spermatocytes in hy- brids of silver fox and blue fox. J. Hered. 79: 338-343. Hale DW and Greenbaum FF, (1988). Chromosomal pair-
ing in deer mice heterozygous for the presence of hete- rochromatic short arms. Genome 30: 44-47.
Howel WM and Black DA, (1980). Controlled silver-stain- ing of nucleolus organizer regions with a protective colloidal developer: a 1-step method. Experientia 36:
Hereditas 133
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
12000)zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
SvnaDtonemal complex in hvbrid bulls 79Pereira Net0 OA and Lobato JFP, (1998). Efeitos da utilizaqBo de pastagens nativas melhoradas no desen- volvmiento e comportamento reprodutivo, de novilhas de corte. R. Bras. Zootec. 27: 60-65.
Santos JL, (1993). Spreading synaptonemal complex from the grasshopper Chorthippus jacobsi: pachytene and observations. Hereditas 118: 235-241.
Schmekel K, Wahrman J and Daneholt B, (1993). The central region of the synaptonemal complex in Blaps cribrosa studied by electron nuicroscope tomography. Chromosoma 102: 669-681.
Sharp PJ, (1986). Synaptic adjustment a t a C-band het- erozygosity. Cytogenet. Cell Genet. 41: 56-57.
Sudman PD, Greenbaum IF, Hale DW and Smith AS,
(1989). Synaptic adjustment in Peromyscus beatae
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
(RO-dentia: Cricetidae) heterozygous for interstitial hete- rochromatin. Cytogenet. Cell Genet. 50: 1-5.
Switonski M, Ansari HA, Jung HR and Stranziger G,
(1990). Synaptonemal complex analysis in primary sper- matocytes of cattle x zebu hybrids (Bos taurus x Bos
indicus). J. Anim. Breed. Genet. 107: 229-238.
Trematore RL, (1996). Estimativas de efeitos aditivos e heterdticos, para caracteristicas de crescimento e des- mama, em bezerros da raqa Nelore e cruzados Charoles-
Nelore. M.S. Thesis, Umiversidade Federal de
zyxwvutsrqponmlkjihgfedcbaZYXWVUTSRQPONMLKJIHGFEDCBA
S5oCarlos,