Stimulatory sub-lethal response of a generalist predator to permethrin: hormesis, hormoligosis or homeostatic regulation?
27
0
0
Texto
(2)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)(19)
(20)
(21)
(22)
(23)(24)
(25)
(26)
(27)
Imagem
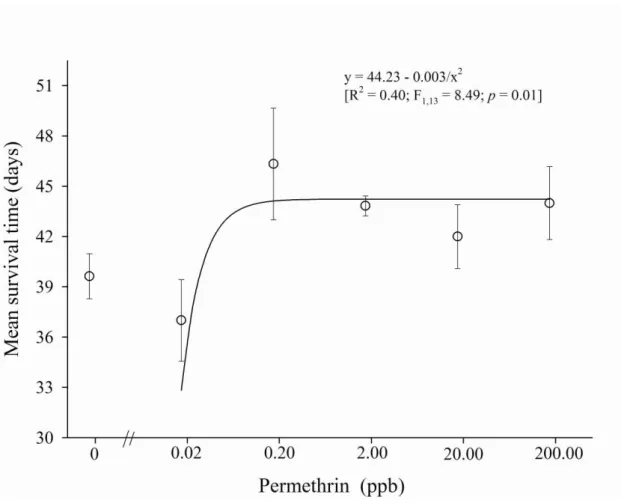
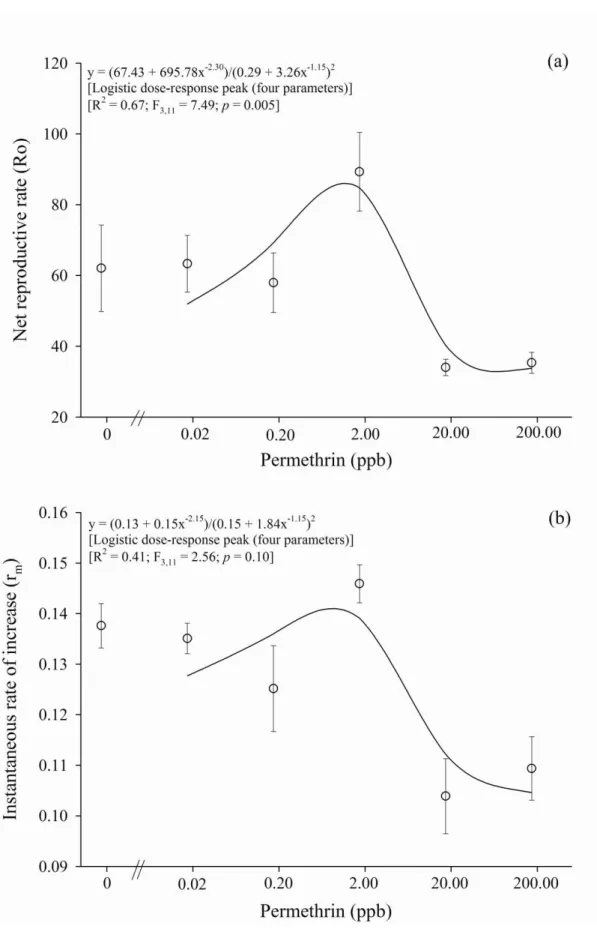
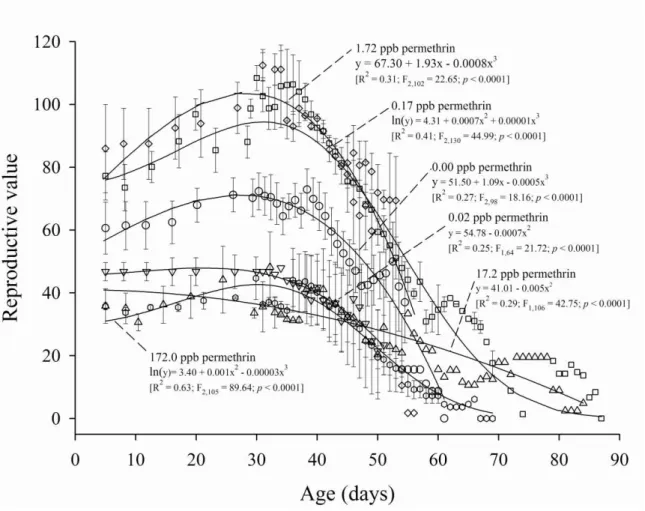
Documentos relacionados
In short, the drug industry clearly had negative implications over Colombia both internally and externally: internally, the cartels promoted violence, fomented
Sempre inquieta, realizando leituras principalmente sobre Física Moderna Contemporânea (FMC), por se tratar de um assunto que faz parte de nosso cotidiano e estar prevista
Este estudo teve como principal objetivo, investigar em que medida a relação entre o foco regulatório coletivo, enquanto estado emergente motivacional, e a eficácia do trabalho
Entre as atividades agroindustriais praticadas nos assentamentos, chamam a atenção as experiências com moinhos coloniais por envolver aspectos como: a) a amplitude de sua
Modelagem numérica e computacional com similitude e elementos finitos : equação preditiva geral para o cálculo da força de retenção em freios de estampagem [livro eletrônico]
Os objetivos deste trabalho são conhecer os problemas orais resultantes da radioterapia feita para o tratamento de neoplasias na área da cabeça e pescoço assim
Deve-se acrescentar, ainda, os jornais vinculados aos novos grupos étnicos presentes na província, como os alemães, com o jornal católico Deutsches Volksblatt, de 1871;
Baixas doses de glyphosate pode estimular o crescimento de uma variedade de espécies de plantas o efeito de hormesis provavelmente esteja relacionado com o sítio de
