DANIELA KELLER MENEZES
Transferência de alimento aos infantes de
Callithrix jacchus
em
uma situação de escassez de recurso alimentar
Dissertação
apresentada
à
Universidade
Federal
do
Rio
Grande do Norte, para obtenção do
título de Mestre em Psicobiologia.
Natal
DANIELA KELLER MENEZES
Transferência de alimento aos infantes de
Callithrix jacchus
em
uma situação de escassez de recurso alimentar
Dissertação
apresentada
à
Universidade
Federal
do
Rio
Grande do Norte, para obtenção do
título de Mestre em Psicobiologia.
Orientadora:
Maria
de
Fátima
Arruda de Miranda
Natal
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de
Biociências
Menezes, Daniela Keller.
Transferência de alimento aos infantes de Callithrix jacchus em situação de escassez de recurso alimentar / Daniela Keller Menezes. – Natal, RN, 2010.
78 f. : Il.
Orientadora: Profª. Maria de Fátima Arruda de Miranda.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-Graduação em Psicobiologia.
1. Callithrix jacchus – Dissertação 2. Infantes – Dissertação. 3. Transferência de alimento – Dissertação. I. Miranda, Maria de Fátima Arruda de. II. Universidade Federal do Rio Grande do Norte. III. Título.
RN/UF/BSE-CB
CDU 599.822
Título: Transferência de alimento aos infantes de Callithrix jacchus em uma situação de escassez de recurso alimentar.
Autora: Daniela Keller Menezes
Data da defesa:
Banca Examinadora:
_________________________________________
Profa. Maria Adélia Borstelmann de Oliveira
Universidade Federal Rural de Pernambuco
_________________________________________
Profa. Fívia de Araújo Lopes
Universidade Federal do Rio Grande do Norte
_________________________________________
Maria de Fátima Arruda de Miranda
“Nenhuma grande vitória é possível sem
que tenha sido precedida de pequenas vitórias
AGRADECIMENTOS
A Deus, pelas pessoas especiais que Ele sempre coloca em meu caminho.
Aos meus pais e ao meu irmão por todo amor, zelo, compreensão, oportunidades, conselhos,
respeito e apoio incondicional às minhas decisões.
Ao meu noivo Ricardo por todo seu amor, companheirismo, apoio e ajuda em todas as etapas
deste trabalho, especialmente pela paciência em me ensinar como preparar e analisar todos os
meus dados.
À professora Fátima Arruda pela orientação e apoio na realização deste trabalho, pelas
informações compartilhadas durante as reuniões do grupo e pelo respeito ao meu ritmo de
produção.
Aos professores do PPg em Psicobiologia pelos conhecimentos científicos transmitidos nas
aulas que contribuíram para meu crescimento profissional.
À professora Adélia pela oportunidade de iniciar meus estudos do comportamento animal, que
me fizeram chegar até aqui!
Aos meus colegas de mestrado Samuel Limaverde, Luciane Coletti, Tiago Eugênio (Kutako),
Ao Victor Shiramizu por me acompanhar na fase piloto e nas primeiras observações,
esclarecendo minhas dúvidas, e pela coleta dos dados nos dias em que não pude estar
presente.
À minha amiga Rafaela Cobuci pela companhia e ótimas conversas antes dos experimentos,
pela contribuição na coleta dos dados e por todo o empenho em ajudar sempre!
À banca examinadora pelas críticas e sugestões para melhorar o meu trabalho.
Ao CNPq pela bolsa de mestrado (Ó, querida bolsa de mestrado, que saudades de você!).
Aos funcionários do Núcleo de Primatologia da UFRN pela colaboração na realização do meu
estudo.
Aos saguis, especialmente aos infantes, que me divertiram com suas brincadeiras antes,
durante e após as observações!
SUMÁRIO
RESUMO ... 1
ABSTRACT ... 2
INTRODUÇÃO ... 3
OBJETIVOS ... 12
Geral ... 12
Específicos ... 12
HIPÓTESES E PREDIÇÕES... 13
METODOLOGIA ... 14
Animais ... 14
Procedimento experimental ... 16
Coleta de dados antes do procedimento experimental ... 18
Categorias comportamentais coletadas durante o procedimento experimental ... 18
Categoria comportamental coletada antes do procedimento experimental ... 19
Análise dos dados ... 20
RESULTADOS ... 21
Análises por Condição ... 21
Análises entre Famílias ... 22
Análises com Todas as Famílias ... 25
Análises por Família ... 26
Família E1: ... 26
Família E2... 28
Família A1... 31
Família A2... 34
DISCUSSÃO ... 57
Transferência de Alimento ... 57
Distância em Relação à Fonte de Alimento ... 67
Local de Consumo ... 69
Transporte ... 70
ÍNDICE DE FIGURAS
Figura 1
–
Prato com alimento utilizado no procedimento experimental. ... 17
Figura 2
–
Frequência de: (A) solicitação bem sucedida; (B) solicitação mal
sucedida; (C) proximidade dos infantes à fonte de alimento; (D) distância dos
infantes à fonte de alimento; (E) consumo da fonte e (F) consumo fora da fonte,
nas condições de escassez e abundância de alimento. ... 37
Figura 3
–
Frequência de: (A) solicitação bem sucedida nas famílias A1, A2 e
E2; (B) solicitação mal sucedida; (C) proximidade dos infantes à fonte de
alimento; (D) distância dos infantes à fonte de alimento; (E) consumo na fonte
de alimento pelos infantes e (F) consumo fora da fonte de alimento pelos
infantes nas famílias E1, A1, A2 e E2. ... 38
Figura 4
–
Frequência de: (A) transporte da prole e (B) tentativa de roubo
efetuado pelos infantes nas famílias A1, A2, E1 e E2; (C) roubo efetuado pelos
infantes das famílias A1, E1 e E2; (D) eventos em que os infantes das famílias
A1, A2 e E2 tiveram seus itens alimentares roubados por algum membro do
grupo. ... 39
Figura 5
–
(A) Frequência da proximidade dos infantes de todas as famílias à
fonte de alimento no decorrer das 20 primeiras semanas de vida. (B) Frequência
de obtenção de alimento pelos infantes de todas as famílias por partilha passiva
e alimentação independente. ... 43
Figura 6
–
Família E1
–
Frequência de solicitação mal sucedida em relação: (A)
ao sexo dos cuidadores; (B) ao
status
social dos cuidadores; (C) à identidade dos
cuidadores. ... 44
Figura 7
–
Família E1
–
Frequência de: (A) proximidade e distância dos infantes
em relação à fonte de alimento; (B) consumo na fonte e fora da fonte de
alimento; (C) consumo na fonte e fora da fonte de alimento no decorrer das 20
primeiras semanas de vida... 45
Figura 8
–
Família E1
–
Frequência de transporte da prole: (A) em relação ao
sexo dos cuidadores; (B) em relação ao
status
social dos cuidadores; (C) em
relação à idade dos cuidadores; (D) em relação à identidade dos cuidadores; (E)
no decorrer das 20 primeiras semanas de vida. ... 46
cuidadores; (C) mal sucedida em relação ao
status
social dos cuidadores; (D)
mal sucedida em relação à identidade dos cuidadores. ... 47
Figura 10
–
Família E2
–
Frequência de: (A) proximidade e distância dos
infantes em relação à fonte de alimento; (B) consumo na fonte e fora da fonte de
alimento; (C) consumo na fonte e fora da fonte de alimento no decorrer das 20
primeiras semanas de vida... 48
Figura 11
–
Família E2
–
Frequência de transporte da prole: (A) em relação ao
sexo dos cuidadores; (B) em relação ao
status
social dos cuidadores; (C) em
relação à identidade dos cuidadores; (D) no decorrer das 20 primeiras semanas
de vida. ... 49
Figura 12
–
Família A1
–
Frequência de solicitação bem sucedida em relação:
(A) ao sexo dos cuidadores; (B) ao
status
social dos cuidadores; (C) à idade dos
cuidadores; (D) à identidade dos cuidadores. ... 50
Figura 13
–
Família A1
–
Frequência de solicitação mal sucedida em relação:
(A) ao sexo dos cuidadores; (B) ao
status
social dos cuidadores; (C) à idade dos
cuidadores; (D) à identidade dos cuidadores. ... 51
Figura 14
–
Família A1
–
Frequência de: (A) solicitação bem e mal sucedida;
(B) proximidade e distância dos infantes em relação à fonte de alimento; (C)
consumo na fonte e fora da fonte de alimento; (D) consumo na fonte e fora da
fonte de alimento no decorrer das 20 primeiras semanas de vida. ... 52
Figura 15
–
Família A1
–
Frequência de transporte da prole: (A) em relação ao
sexo dos cuidadores; (B) em relação ao
status
social dos cuidadores; (C) em
relação à idade dos cuidadores; (D) em relação à identidade dos cuidadores; (E)
no decorrer das 20 primeiras semanas de vida. ... 53
Figura 16
–
Família A2
–
Frequência de solicitação mal sucedida em relação:
(A) ao sexo dos cuidadores; (B) ao
status
social dos cuidadores; (C) à idade dos
cuidadores; (D) à identidade dos cuidadores. ... 54
Figura 17
–
Família A2
–
Frequência de: (A) proximidade e distância dos
infantes em relação à fonte de alimento; (B) consumo na fonte e fora da fonte de
alimento; (C) consumo na fonte e fora da fonte de alimento no decorrer das 20
primeiras semanas de vida... 55
ÍNDICE DE TABELAS
Tabela I - Composição das famílias e alterações na composição durante o
período de estudo. ... 15
Tabela II
–
Contexto Alimentar ... 40
1 RESUMO
O gênero Callithrix, assim como os demais calitriquídeos, apresenta um sistema de
criação cooperativa da prole, caracterizado pela contribuição dos pais e de indivíduos não
reprodutores do grupo social no cuidado aos infantes. Esse cuidado é fornecido através da
amamentação, do transporte, da supervisão dos infantes e da transferência de alimento. Neste
estudo, investigamos o cuidado à prole em Callithrix jacchus através da transferência de
alimento em grupos familiares sob condições de abundância e de escassez de recursos
alimentares. Buscamos também verificar a relação entre a participação dos indivíduos no
transporte dos filhotes e na partilha de alimento. Foram observadas quatro famílias habitantes
do Núcleo de Primatologia da Universidade Federal do Rio Grande do Norte, totalizando sete
infantes, sendo seis gêmeos e um filhote único. Cada infante foi observado duas vezes por
semana, da 1ª a 20ª semana de vida, através dos métodos focal contínuo e focal instantâneo,
com intervalos de um minuto. As observações eram realizadas por 10 minutos e iniciavam-se
com o fornecimento do alimento para cada família. Duas famílias foram testadas sob condição
de escassez de recurso alimentar (condição experimental) e outras duas foram submetidas à
condição de abundância de alimento, ambas as condições com restrição de acesso à fonte de
alimento. A condição de escassez de alimento não influenciou a taxa de partilha de alimento
com os filhotes que, de um modo geral, foi baixa. Todas as partilhas observadas foram do tipo
passiva. Somente uma família apresentou níveis maiores de solicitação, provavelmente em
decorrência da perda da mãe durante a fase de amamentação da prole. Nessa família, os
machos dominante e subadulto foram os principais responsáveis pelas partilhas. A condição
experimental influenciou a distância dos infantes em relação à fonte de alimento, de modo que
os infantes das famílias que receberam alimento abundante permaneceram mais próximos à
fonte. Entretanto, acredita-se que o local de consumo do alimento não tenha sido influenciado
pela condição experimental. Não houve relação entre a participação dos indivíduos no
transporte e na partilha de alimento com os filhotes. Portanto, sugere-se que, de um modo
geral, a transferência de alimento em C. jacchus seja caracterizada pela tolerância dos
cuidadores às solicitações, representada pela partilha passiva. As taxas de partilha com os
infantes foram baixas, provavelmente devido à competição alimentar intragrupo gerada pelo
2 ABSTRACT
The genus Callithrix, like other marmosets, presents a cooperative breeding system,
characterized by the contribution of parents and non-breeding individuals of the social group
in the care of infants. This care is provided through lactation, transport, supervision of infants
and food transfers. In this study, we investigated the care of offspring in Callithrix jacchus
through the food transfers in family groups under conditions of abundance and scarcity of
food resources. We also attempted to verify the relationship between the participation of
individuals in the transport of infants and in the food sharing. We observed four families at
the Núcleo de Primatologia of the Universidade Federal do Rio Grande do Norte, in a total of
seven infants, six twins and a single infant. Each infant was observed twice a week, from 1st
to 20th week of life using the methods focal continuous and focal instantaneous sampling
with one minute intervals. The observations were carried by 10 minutes and beginning with
the supply of food for each family. Two families were tested under conditions of scarcity of
food resources (experimental condition) and two others were subjected to conditions of
abundant food, both conditions with restricted access to food supply. The condition of food
scarcity did not influence the rate of food sharing with the infants that, in general, was
considered low. All shares observed were passive sharings. Only one family showed higher
levels of food begging, probably due to the loss of the mother during the period of lactation of
her offspring. In this family, the dominant and subadult males were primarily responsible for
the food sharing. The experimental condition influenced the distance of infants in relation to
food source, so the infants in families that received abundant food remained closer to the
source. However, it is believed that the place of food consumption was not influenced by
experimental condition. There was no relationship between the participation of individuals in
the transport and food sharing with their young. Therefore, it is suggested that, in general, the
transfer of food in C. jacchus is characterized by tolerance to the demands of caregivers,
represented by the passive sharing. Rates of sharing with infants were low, probably due to
3 INTRODUÇÃO
A família Callitrichidae é composta por seis gêneros: Callimico, Callithrix, Cebuella,
Mico, Leontopithecus e Saguinus. Atualmente, as espécies que anteriormente pertenciam ao
gênero Callithrix são classificadas da seguinte maneira: as nativas da Amazônia são
identificadas pelo gênero Mico e as que ocorrem fora desse bioma permanecem como
Callithrix (Rylands et al. 2000). De um modo geral, os primatas desta família apresentam
hábito diurno, são arborícolas (Stevenson & Rylands 1988; Rylands et al. 1993) e consomem
itens alimentares bastante variados, sendo considerados animais frugívoros-insetívoros
(Stevenson & Rylands 1988). Alguns gêneros, como Cebuella e Callithrix, são considerados
especialistas por possuírem adaptações morfológicas (Rosenberger 1992; Feagle 1998) e
fisiológicas para a obtenção e digestão de exsudados de plantas (Feagle 1998). Quanto aos
aspectos reprodutivos, as fêmeas apresentam estro pós-parto e proles normalmente gemelares
(Brown & Mack 1978).
Os membros do gênero Callithrix não apresentam dimorfismo sexual. A massa do
animal adulto varia de 300 a 450g e de um filhote recém-nascido, de 22 a 38g. Em média, são
menores que Saguinus e maiores que Cebuella. Habitam preferencialmente matas secundárias
e vivem em grupos sociais cujo tamanho, incluindo os infantes dependentes, varia de três a 15
indivíduos (Stevenson & Rylands 1988). Entretanto, há registro de 17 indivíduos em um
mesmo grupo social de Callithrix jacchus em ambiente natural (Albuquerque & Arruda
1997).
A organização social do gênero Callithrix em cativeiro apresenta várias características
que podem ser atribuídas a espécies com a organização social monogâmica (Yamamoto
4
identificados três possíveis sistemas: monoginia, monoginia com tentativas de reprodução das
subordinadas (pseudo-poliginia) e poliginia (Yamamoto et al. 2007).
O gênero Callithrix, assim como os demais calitriquídeos, apresenta um sistema de
criação cooperativa dos filhotes (Stevenson & Rylands 1988). Neste sistema, os indivíduos
não reprodutores do grupo social, além dos pais, contribuem no cuidado à prole dependente
(Yamamoto & Box 1997; Yamamoto 2005). O alto custo energético do cuidado para a fêmea
reprodutora, que por apresentar estro pós-parto pode engravidar novamente enquanto ainda
está amamentando, provavelmente contribuiu para o surgimento deste sistema cooperativo
(Alonso & Langguth 1989; Ximenes & Sousa 1996; Rylands 1996).
O período de infância em Callithrix tem duração de cinco meses (Yamamoto 1993) e
os filhotes são muito dependentes do cuidado, especialmente nas primeiras semanas de vida.
Esse cuidado é fornecido através da amamentação, do transporte, da supervisão dos infantes e
da transferência de alimento (Snowdon 1996; Tardif 1994).
A amamentação é um tipo de cuidado efetuado exclusivamente pela mãe. No ambiente
natural, o desmame dos filhotes de Callithrix jacchus ocorre no 3º mês de vida (Albuquerque
& Arruda 1997); em cativeiro, o desmame é mais precoce, sendo observado no 2º mês de vida
(Yamamoto 1993; Ximenes & Sousa 1996).
O transporte dos filhotes é uma atividade compartilhada entre todos os membros do
grupo, embora ocorra maior participação do pai e da mãe no carregar durante os primeiros
dias de vida (Oliveira 1996; Ximenes & Sousa 1996). Hattori (2004), estudando aspectos do
cuidado em duas proles consecutivas de filhote único de um grupo de Callithrix jacchus em
ambiente natural, observou que o pai foi o principal responsável pelo transporte dos filhotes,
sendo seguido pelos dois machos adultos subordinados do grupo, com contribuições
equivalentes. A participação do pai no transporte da prole é influenciada pelo tamanho do
5
pai é significativamente menor do que em grupos menores (Ximenes & Sousa 1996; Santos et
al. 1997). O nível de participação dos demais membros do grupo no transporte da prole é
influenciada pela idade do cuidador (Yamamoto et al. 1996; Santos et al. 1997; Yamamoto &
Box 1997), de modo que os indivíduos mais velhos tendem a transportar os infantes mais
frequentemente do que os animais mais jovens (Digby 1995; Santos et al. 1997; Yamamoto &
Box 1997). A contribuição dos ajudantes no transporte da prole diminui os gastos energéticos
da mãe com o carregar na fase em que ela está comprometida com a amamentação dos
infantes.
O período compreendido entre as semanas cinco a 10 é caracterizado pelo início do
processo de independência dos infantes. Embora ainda sejam transportados por um tempo
considerável, os infantes deixam o dorso dos cuidadores espontaneamente por curtos
períodos, ao mesmo tempo em que os cuidadores tornam-se menos tolerantes às solicitações
de transporte (Yamamoto 1993). Neste período, observa-se o comportamento de supervisão
dos infantes por parte dos integrantes do grupo social, em que o cuidador mantém-se próximo
ao filhote, aparentemente protegendo-o. No momento em que o filhote começa a realizar suas
primeiras experiências se deslocando de forma independente, o cuidador permanece atento e
responsivo às solicitações de ajuda, por exemplo, quando o filhote cai de uma árvore (Alonso
& Langguth 1989). Neste início do processo de independência, os infantes experimentam
alimentos sólidos (Yamamoto 1993).
A transferência de alimento é outro importante aspecto do cuidado cooperativo (Tardif
et al. 1990; Ruiz-Miranda et al. 1999; Price & Feistner 2001) que facilita o início do processo
de alimentação por conta própria (Yamamoto 1993). A transferência de alimento pode ser
definida como o movimento de alimento entre coespecíficos, excluindo a amamentação, que
normalmente envolve um animal dando alimento a outro (Ruiz-Miranda et al. 1999). Os
6
indivíduo que está com o alimento, e receptor em potencial, indivíduo que poderá receber o
alimento (Hattori 2004). As transferências de alimento envolvem vários comportamentos
(Brown & Mack 1978). De acordo com Ruiz-Miranda et al. (1999), nas transferências de
alimento destacam-se dois componentes: a transferência com ou sem resistência do doador.
Dessa forma, os tipos de transferência potencialmente possíveis de serem registradas são:
partilha ativa e passiva, roubo e tentativa de roubo (Ruiz-Miranda et al. 1999).
Dentre os quatro possíveis tipos de transferência de alimento, em calitriquídeos
enfatiza-se a diferença entre espécies com relação à ocorrência de partilha ativa e passiva. A
partilha passiva, geralmente iniciada pela solicitação do infante, é a forma mais comum e
ocorre na maioria das espécies de primatas estudadas, sendo encontradas em todas as espécies
da família Callitrichidae (Feistner & Price 1999). Já a partilha ativa, iniciada pelo possuidor
do alimento, é o tipo mais raro tendo sido registrada sistematicamente até o momento apenas
em algumas espécies de calitriquídeos como: Saguinus oedipus (Feistner & Price 1990, 1991,
1999; Price & Feistner 1993; Roush & Snowdon 2001; Joyce & Snowdon 2007),
Leontopithecus chrysopygus (Feistner & Price 2000), L. rosalia (Brown & Mack 1978;
Ruiz-Miranda et al. 1999), L. chrysomelas (Feistner & Price 1991), Cebuella pygmaea (Feistner &
Price 1991), Callimico goeldii (Feistner & Price 1991) e Callithrix flaviceps (Ferrari 1987).
Existem diferenças entre espécies e gêneros da família Callitrichidae em relação a
aspectos da partilha de alimento, tais como a idade do filhote em que se inicia a partilha, a
proporção de alimento recebido dos possuidores do alimento e o tipo de partilha ocorrido
(Price & Feistner 2001). Contudo, também são observadas semelhanças considerando-se que
a partilha se inicia quando o filhote imaturo é incapaz de se alimentar sozinho e decresce até
cessar quando este é capaz de se alimentar independentemente (Brown & Mack 1978;
Feistner & Price 1990; Ruiz-Miranda et al. 1999; Feistner & Price 2000). Dados de campo
7
para eles ou que requerem habilidades para serem processados, como insetos grandes e
sementes com envoltório rígido (Ferrari 1987). Em relação à contribuição dos membros do
grupo na partilha de alimento com a prole dependente, observa-se que todos ou a maioria dos
indivíduos podem participar desta forma de cuidado (Cutrim 2007). Através da partilha, os
irmãos mais velhos podem aumentar sua aptidão abrangente. Além disso, a contribuição dos
irmãos mais velhos beneficia os pais tanto diretamente, por meio da redução dos custos
energéticos envolvidos na criação de filhotes, quanto indiretamente, através do aumento na
quantidade de alimento disponível aos infantes (Feistner & Price 1990).
Brown e Mack (1978), estudando 10 grupos cativos de L. rosalia, observou que as
transferências de alimento entre os indivíduos do grupo foram frequentes e não se
restringiram às transferências pais-prole. Ocorreram transferências entre o casal reprodutor,
entre os pais e os juvenis e entre os juvenis e seus irmãos mais velhos. Foram registrados os
seguintes tipos de transferência: partilhas passivas e ativas e roubo. Os animais apresentavam
um repertório de sinais comportamentais que poderiam indicar aos recebedores um convite à
partilha de itens alimentares.
Um estudo com nove famílias cativas de S. oedipus, realizado por Feistner e Price
(1990), mostrou que os itens alimentares foram partilhados pelos pais e irmãos mais velhos
em resposta às solicitações dos infantes, além de terem sido ofertados sem solicitações
prévias. Os indivíduos mais velhos e sexualmente maduros e suprimidos participaram mais
das partilhas do que os imaturos. As partilhas atingiram um valor máximo na 12ª semana e,
então, declinaram gradualmente. O tamanho do grupo influenciou a frequência das partilhas:
infantes de famílias maiores receberam mais alimento provido do que os de grupos menores,
embora a partilha por indivíduo nas famílias menores tenha sida maior.
Visando investigar a ocorrência de partilha ativa em diferentes espécies da família
8
ocorrência de partilha ativa em apenas quatro das oito espécies de calitriquídeos estudadas.
Por outro lado, a partilha passiva foi observada em todas as espécies. Assim, os autores
sugeriram que o comportamento de oferta estivesse provavelmente relacionado com a
ecologia de cada espécie.
Ruiz-Miranda et al. (1999), investigando a transferência de alimento em L. rosalia no
ambiente natural em sete grupos nascidos em vida livre e cinco grupos nascidos em um
zoológico e, posteriormente, reintroduzidos, observou que os animais adultos foram
responsáveis por grande parte das partilhas destinadas aos filhotes. A maioria dos itens
partilhados foi presa ou frutos que requeriam habilidade para serem processados. Em geral, as
partilhas foram precedidas por vocalizações e comportamentos de solicitação por parte dos
infantes. Também foram registrados eventos de roubo de alimento.
Feistner e Price (2000), investigando o comportamento de transferência de alimento
em 10 infantes cativos de L. chrysopygus, observaram um pico de ocorrência das partilhas
passivas na 9ª semana de idade. Até a 15ª semana, os infantes ainda consumiam mais itens
providos do que obtidos por conta própria, período que coincidiu com fim do transporte
(semanas 10 e 11) e o desmame (semana 13). A partilha ativa também ocorreu, mas foi baixa,
totalizando 5,3% dos itens transferidos.
Queiroz (2003), estudando partilha de alimento em três famílias de C. jacchus cativos
com restrição de acesso à fonte alimentar, observou que, em geral, as partilhas foram
precedidas por solicitação dos infantes e o pai foi o principal doador de alimento à prole,
seguido pelos demais machos adultos dos grupos. O sucesso das partilhas foi influenciado
pela idade dos filhotes, sendo maior em infantes mais jovens, com redução acentuada a partir
do 4º mês de vida. O tamanho da prole também influenciou a ocorrência de partilhas, de
modo que infantes de proles gemelares receberam mais alimento provido do que o filhote
9
Nascimento (2004), estudando partilha de alimento em cinco famílias de C. jacchus
cativos, observou que a idade dos infantes influenciou a frequência da partilha de alimento, de
modo que os filhotes mais dependentes receberam mais alimento provido do que os
independentes. A partilha diferiu entre os membros do grupo, sendo que os indivíduos adultos
partilharam mais que os subadultos e juvenis. Todos os registros de partilha foram do tipo
passiva e as solicitações bem sucedidas foram superiores às mal sucedidas em todos os
grupos.
Em seu estudo com duas proles consecutivas de filhote único de um grupo de C.
jacchus em ambiente natural, Hattori (2004) observou que o grau de dependência dos infantes
influenciou as interações de partilha de alimento: as partilhas de alimentos sólidos
iniciaram-se no período de transição dos infantes à independência alimentar, até que estes adquirisiniciaram-sem
habilidade para obtenção do alimento. A partir desse momento, houve uma diminuição da
tolerância dos cuidadores, através do aumento da resistência às partilhas. O autor também
verificou uma relação direta na participação dos indivíduos no transporte, supervisão dos
infantes e partilha de alimento, sendo o pai e dois machos subordinados os indivíduos que
mais contribuíram nestas três formas de cuidado com os filhotes.
Brown, Almond e Bates (2005) investigando padrões de solicitação e transferência de
alimento em seis grupos cativos de C. jacchus providos com itens alimentares que variavam
em palatabilidade e novidade, observaram que os infantes solicitavam mais alimentos novos
do que familiares. No entanto, os adultos partilharam mais itens familiares do que novos,
principalmente quando estes eram impalatáveis.
Cutrim (2007) acompanhou, em ambiente natural, o desenvolvimento durante todo o
período de infância de três proles de C. jacchus: uma prole em um grupo que habitava uma
área de pomar e duas proles em um grupo habitante de área de Caatinga. A autora verificou a
10
alimentos mais transferidos foram os tipos considerados mais difíceis de serem obtidos pelos
infantes por conta própria. A partilha passiva foi a principal forma de provisão de alimento
sólido aos infantes em ambos os ambientes. A partilha ativa, evento raramente registrado
nesta espécie, ocorreu apenas com as duas proles do grupo que habitava a área de Caatinga.
Os demais tipos de transferência de alimento relatados neste trabalho foram mais frequentes
no ambiente de Caatinga do que no pomar. A autora sugere que as condições ecológicas desse
ambiente podem ter contribuído para a maior frequência dos comportamentos de transferência
observados, incluindo a ocorrência de partilha ativa.
De um modo geral, observa-se que as solicitações e partilhas de alimento aumentam
com o início do desmame dos infantes, que ainda apresentam pouca habilidade para captura
de presas vivas (Albuquerque & Arruda 1997). Entretanto, à medida que os filhotes se
desenvolvem, a tolerância dos cuidadores em partilhar o alimento diminui (Cutrim 2007).
Quando os infantes estão mais ágeis e mais velhos, eventualmente, eles podem roubar o
alimento com movimentos rápidos (Hattori 2004).
A literatura apresenta várias hipóteses sobre as funções das transferências de alimento
em primatas: (1) favorecer o desmame precoce do infante (Ruiz-Miranda et al. 1999); (2)
assistência no forrageio (Price & Feistner 1993; Ruiz-Miranda et al. 1999); (3) aprendizagem
dos itens alimentares adequados (Price & Feistner 1993; Ruiz-Miranda et al. 1999); (4)
diminuição de neofobia alimentar (Ruiz-Miranda et al. 1999); (5) transferência de itens raros
(Ruiz-Miranda et al. 1999); (6) benefícios sociais (Ruiz-Miranda et al. 1999). Tais hipóteses
não são mutuamente exclusivas, mas dão suporte a predições diferentes acerca da natureza
dos itens partilhados e identidade dos doadores e recebedores (Ruiz-Miranda et al. 1999).
A transferência de alimento em infantes de C. jacchus representa uma atividade com
expressão acentuada apenas em um determinado período do desenvolvimento dos infantes, o
11
padrão de transferências aos infantes em função das condições ecológicas, é interessante
testarmos a influência da variação da disponibilidade de recurso. Em decorrência da escassez
de informações acerca dessa atividade em filhotes C. jacchus, durante todo o período de
infância, ou seja, da 1ª a 20ª semana de vida dos filhotes, observamos este tipo de interação a
partir de experimentos de laboratório com alimentos disponíveis em quantidades diferentes.
Desse modo, a observação em cativeiro fornece novas evidências de transferência de alimento
12 OBJETIVOS
Geral
Investigar o cuidado à prole em Callithrix jacchus através da transferência de alimento
em grupos familiares sob condições de abundância e de escassez de recursos alimentares.
Específicos
1. Comparar os diferentes tipos de transferência de alimento nas duas condições e
quais indivíduos do grupo estão mais envolvidos;
2. Registrar os indivíduos que participam no transporte dos infantes;
3. Verificar a relação entre a participação no transporte dos filhotes e na partilha
13
HIPÓTESES E PREDIÇÕES
Hipótese 1: A disponibilidade de recursos alimentares influencia a partilha de
alimento.
Predição 1: Espera-se que as taxas de solicitação bem sucedidas, ou seja, os eventos de
partilha de alimento, sejam mais frequentes nas famílias submetidas à condição de escassez de
recursos alimentares.
Predição 2: Espera-se que a frequência de proximidade dos infantes à fonte de
alimento seja maior nas famílias testadas na condição de escassez de recursos alimentares.
Predição 3: Espera-se que os níveis de consumo na fonte de alimento por parte dos
infantes sejam maiores nas famílias submetidas à condição de escassez de recursos.
Hipótese 2: Os diferentes tipos de cuidado são oferecidos pelos mesmos indivíduos.
Predição 4: Espera-se que os indivíduos que participam mais no transporte partilhem
14
MÉTODO
Animais
Foram acompanhadas quatro famílias de C. jacchus do Núcleo de Primatologia da
Universidade Federal do Rio Grande do Norte (Tabela I), totalizando sete infantes, sendo seis
gêmeos e um filhote único. A classificação etária dos indivíduos adotada neste trabalho foi
proposta por Yamamoto (1993).
As famílias estavam alojadas em recintos de alvenaria com portões de ferro e piso de
cimento, com 2x1x2 m, equipados com plataformas e troncos de madeira, comedouros,
bebedouro e caixa ninho. Cada recinto era parcialmente coberto com telhas de cerâmica,
permitindo que os saguis ficassem expostos à temperatura e ciclo de luz naturais. Os animais
mantinham contato visual, auditivo e olfativo com indivíduos de outras famílias.
De acordo com o manejo alimentar dos saguis do Núcleo de Primatologia, a
alimentação era servida duas vezes ao dia, em tigelas de aço inox colocadas nos comedouros
no interior do recinto e a água, oferecida ad libitum no bebedouro. A refeição da manhã,
servida entre 7 e 9 h, consistia de uma papa (pão, banana, leite, fibra e gérmen de trigo,
extrato de soja, ovo cozido e mel) e a da tarde, fornecida entre 12 e 14 h, era formada por
frutos da época (banana, melancia, mamão) ou batata doce e ovos ou frango, todos cozidos. A
alimentação da manhã era suplementada com a adição de vitaminas e cálcio à papa e a da
15
Tabela I - Composição das famílias e alterações na composição durante o período de estudo.
Família E1 E2 A1 A2
Indíviduo
♂
D♂
D♂
D♂
D♀D ♀D ♀D † (8ª) ♀D
♂
A♂
A1 ♀A♂A1 (11ª semana)
♂
J S (11ª semana)♂
A2♂S (7ª semana)
♀A♀I
♂
A3♂
S A (11ª semana)♂A2 (20ª semana)
♂
I ♀A♂
J♂
S-
♂
J ♀I ♀S- ♀J
♂
I ♀J-
♂
I -♂
I-
♂
I - -Total de
indivíduos 6 10 8 9
16
Para a identificação dos animais, foi realizada a tintura dos tufos periauriculares, dorso
e/ou patas dianteiras com solução de ácido pícrico a 5%. As partes do corpo pintadas com
essa substância adquiriram uma cor amarelada.
Procedimento experimental
O experimento foi realizado de modo a simular duas condições: abundância de recurso
alimentar, com a colocação de duas tigelas com frutas para duas famílias (A1 e A2) e escassez
de recurso alimentar, com a apresentação de apenas uma tigela para duas outras famílias (E1 e
E2).
Em ambas as condições, cada prato continha dois pedaços de banana, de
aproximadamente 1 cm3, por indivíduo de cada família. Cada recipiente foi coberto por um
disco de acrílico transparente de 1 mm de espessura com um orifício central de 2 cm de
diâmetro para dificultar o acesso dos infantes ao recurso e propiciar a ocorrência de
comportamentos de transferência dos itens alimentares (Figura 1). Com o disco, os membros
do grupo conseguiram colocar apenas uma mão de cada vez para obter o alimento. A(s)
tigela(s) era(m), então, colocada(s) no(s) comedouro(s) no interior do recinto (fonte de
17
Figura 1 – Prato com alimento utilizado no procedimento experimental.
O acompanhamento dos grupos familiares foi efetuado desde a 1ª semana de vida dos
filhotes até a 20ª semana de idade. Cada infante foi observado duas vezes por semana
alternando-se os dias de observação de cada um deles, através dos métodos: focal contínuo
para o registro dos comportamentos relacionados à partilha de alimento, e focal instantâneo,
com intervalos de um minuto, para o registro da proximidade dos infantes em relação à fonte
de alimento e local de consumo do item alimentar obtido por conta própria. As observações
eram realizadas por 10 minutos e iniciavam-se às 13h com o fornecimento do alimento para
cada família. Para garantir a motivação dos indivíduos adultos, subadultos e juvenis em
relação ao experimento, as tigelas com a alimentação da manhã foram retiradas
18
Coleta de dados antes do procedimento experimental
Antes do procedimento experimental, era realizada a coleta dos dados referentes ao
transporte dos infantes, uma vez que esse comportamento poderia sofrer influência do
procedimento experimental quanto à ocorrência e/ou à freqüência de emissão. Tais dados
eram coletados através do método focal instantâneo, com intervalos de um minuto. Cada
infante era observado em sessões de 15 minutos iniciadas com um intervalo mínimo de 30
minutos antes do início do experimento.
Categorias comportamentais coletadas durante o procedimento experimental
Foram registradas as frequências das seguintes categorias comportamentais:
Proximidade da fonte de alimento: a cada ponto amostral, foi registrado se o filhote
estava a uma distância igual ou inferior a 15 cm da fonte;
Distância da fonte de alimento: a cada ponto amostral, foi registrado se o filhote
estava a uma distância superior a 15 cm da fonte;
Consumo na fonte de alimento: quando o infante pegou o alimento de dentro do
prato por conta própria e o consumiu a uma distância igual ou inferior a 15 cm da
fonte;
Consumo fora da fonte de alimento: quando o infante pegou o alimento de dentro do
prato por conta própria e o consumiu a uma distância superior a 15 cm da fonte;
Partilha ativa: é o tipo de transferência em que o alimento é oferecido pelo possuidor
ao filhote, através de vocalizações e posturas específicas, sem solicitação prévia do
19
Solicitação: quando o filhote visualiza um animal com alimento e emite vocalizações
dirigidas ao mesmo. A solicitação será considerada:
o Bem sucedida: quando o possuidor do alimento atender a solicitação do
infante, transferindo o item alimentar. Caracteriza a ocorrência de partilha
passiva.
o Mal sucedida: quando o possuidor do alimento não atender a solicitação do
infante, caracterizando a não ocorrência de transferência.
Tentativa de roubo: quando o filhote aproxima-se do possuidor do alimento na
tentativa de pegar o item, mas não obtém sucesso;
Roubo: o infante aproxima-se do possuidor do alimento e toma o item para si;
Ser roubado: quando outro indivíduo do grupo pega o alimento que estava em posse
do filhote.
As definições dos comportamentos de partilha ativa, solicitação bem e mal sucedida,
roubo e tentativa de roubo foram baseadas no trabalho de Ruiz-Miranda et al. (1999).
Categoria comportamental coletada antes do procedimento experimental
Transporte dos infantes: registrou-se a identidade do cuidador quando o filhote
encontrava-se em seu dorso;
Na família A1, os dados de transporte começaram a ser coletados a partir da 8ª semana
20 Análise dos dados
Ao fazer o teste de normalidade de Kolmogorov-Smirnov, foi observado que todas as
variáveis medidas apresentaram distribuição não normal (N > 20; D < 0,36; p < 0,01) e,
portanto, os testes utilizados foram não paramétricos.
Nas análises por condição, foi aplicado o teste de Mann-Whitney. Nas análises entre
famílias, foram utilizados os testes de Kruskal-Wallis e Qui-Quadrado. Para uma melhor
compreensão do perfil das famílias estudadas, foram realizadas análises com todas as famílias
utilizando o teste Qui-Quadrado e Mann-Whitney. Nas análises por família, foram
empregados os testes Wilcoxon e ANOVA de Friedman.
O nível de significância adotado foi de 5% ou menor de acordo com o ajuste dos testes
post-hoc. Para a análise estatística dos dados foi utilizado o programa Statistica 8, Copyright©
21
RESULTADOS
Primeiramente, apresentamos os resultados referentes à transferência de alimento
através das seguintes análises: por condição, para verificarmos a provável influência da
disponibilidade de alimento; entre as famílias estudadas, para analisarmos a possível
influência de alguma família sobre os resultados obtidos na comparação por condição; com
todas as famílias, para constatarmos o padrão observado pelos grupos estudados; por família,
para compreendermos o perfil de cada grupo.
Posteriormente, são apresentados os resultados acerca do transporte das proles e a
relação entre as formas de cuidado analisadas.
As figuras com os resultados estão localizadas após o texto dos resultados, nas páginas
37 a 56. As principais informações envolvendo as variáveis analisadas no nosso trabalho
foram resumidas nas tabelas II e III, nas páginas 40 e 42, respectivamente.
Análises por Condição
Solicitação Bem Sucedida (Partilha Passiva) e Solicitação Mal Sucedida (Ausência de
Transferência):
Não houve diferença significativa entre as frequências das solicitações bem sucedidas
na condição de escassez e na condição de abundância de recursos alimentares (Figura 2A;
Teste de Mann-Whitney U = 18; N = 26; p = 0,60). Também não foi constatada diferença
significativa entre as frequências das solicitações mal sucedidas na condição de escassez e na
condição de abundância de alimento (Figura 2B; Teste de Mann-Whitney U = 575; N = 77; p
= 0,09).
22
A proximidade dos infantes à fonte de alimento foi mais frequente na condição de
abundância de recursos (Figura 2C; Teste de Mann-Whitney U = 743,5; N = 111; p < 0,001).
Já a distância dos filhotes à fonte foi maior na condição de escassez (Figura 2D; Teste de
Mann-Whitney U = 670,5; N = 111; p < 0,001).
Consumo na Fonte e Fora da Fonte de Alimento:
O consumo na fonte de alimento (Figura 2E) foi significativamente mais freqüente na
condição de abundância de recursos (Teste de Mann-Whitney U = 959; N = 111; p < 0,01).
Em relação ao consumo fora da fonte (Figura 2F), não houve diferença significativa entre as
condições (Teste de Mann-Whitney U = 1398; N = 111; p = 0,52).
Análises entre Famílias
Solicitação Bem Sucedida (Partilha Passiva):
Na família E1 não ocorreu solicitação bem sucedida. Portanto, esta análise foi
realizada com os dados das outras três famílias. Assim, observa-se que não houve diferença
significativa entre os níveis de solicitação bem sucedida nestas famílias (Figura 3A; Teste de
Kruskal-Wallis = 0,43; N = 26; p = 0,81).
Solicitação Mal Sucedida (Ausência de Transferência):
Houve diferença marginalmente significativa (Figura 3B; Teste de Kruskal-Wallis =
7,05; N = 77; p = 0,07). O teste post-hoc não evidenciou diferença nos pares analisados.
Entretanto, no gráfico, observa-se a tendência da família A1 ter apresentado as maiores
23
Proximidade da Fonte de Alimento:
Houve diferença significativa (Figura 3C; Teste de Kruskal-Wallis = 23,92; N = 111; p
< 0,001). O teste post-hoc mostrou diferença nos pares A2/E1, A2/E2 e A1/E2. Destes, as
famílias A2 e A1 apresentaram as maiores frequências de proximidade da fonte.
Distância da Fonte de Alimento:
Houve diferença significativa (Figura 3D; Teste de Kruskal-Wallis =27,93; N = 111; p
< 0,001). O teste post-hoc revelou diferença nos pares E1/A1, E1/A2, E2/A1 e E2/A2. Deste
modo, os indivíduos das famílias E1 e E2 apresentaram-se menos próximas da fonte.
Consumo na Fonte de Alimento:
Houve diferença significativa (Figura 3E; Teste de Kruskal-Wallis =15,76; N = 111; p
< 0,01). O teste post-hoc evidenciou diferença apenas no par A1/E2, tendo a família A1
apresentado a maior frequência de consumo na fonte.
Consumo Fora da Fonte de Alimento:
Não houve diferença significativa (Figura 3F; Teste de Kruskal-Wallis = 6,79; N =
111; p = 0,08), de modo que as proles das famílias apresentaram níveis similares de consumo
fora da fonte de alimento.
Transporte:
Houve diferença marginalmente significativa (Figura 4A; Teste de Kruskal-Wallis =
7,53; N = 63; p = 0,06). O teste post-hoc não evidenciou diferença nos pares analisados. No
entanto, no gráfico constata-se a tendência da família A1 ter apresentado maior frequência de
24
Tentativa de Roubo:
Considerando-se os eventos em que os infantes tentaram, sem sucesso, obter itens
alimentares em posse de outros indivíduos, foi constatada frequência observada maior do que
o esperado na família A1 (Figura 4B; frequência observada = 18; frequência esperada = 8;
Teste Qui-Quadrado = 18; gl.= 3; p < 0,000).
Roubo:
Para esta análise, foram comparados os dados das famílias A1, E1 e E2, uma vez que a
família A2 não apresentou episódio de roubo realizado pelo infante. Assim, analisando-se os
episódios em que os filhotes roubaram alimento em posse de outros membros do grupo, as
frequências observadas das famílias A1 e E2 foram maiores do que o esperado, mas tal
diferença não foi estatisticamente significativa (Figura 4C; frequência observada na família
A1 = 4; frequência observada na família E2 = 4; frequência esperada = 3; Teste
Qui-Quadrado = 2; gl.= 2; p = 0,368).
Ser Roubado:
Para esta análise, foram comparados os dados das famílias A1, A2 e E2, pois na
família E1 não ocorreu evento de roubo ao infante. Dessa forma, considerando-se os episódios
em que os infantes tiveram o alimento roubado por um membro do grupo, foi constatada
frequência observada maior do que o esperado na família A1, embora tal diferença não tenha
sido estatisticamente significativa (Figura 4D; frequência observada = 6; frequência esperada
25 Análises com Todas as Famílias
Proximidade da Fonte de Alimento:
Considerando-se a proximidade dos infantes de todas as famílias à fonte de alimento a
partir das primeiras incursões fora do dorso do transportador (4ª semana de vida) no decorrer
das 20 semanas, foram constatadas frequências observadas maiores do que o esperado nas
semanas oito a 16 (Figura 5A; média das frequências observadas = 48,67; frequência esperada
= 38,9; Teste Qui-Quadrado = 115,933; gl. = 19; p < 0,001) e nas semanas 18 a 20 (média das
frequências observadas = 45,67; frequência esperada = 38,9), com pico na semana 10
(frequência observada = 59). A semana 17 apresentou frequência observada próxima do
esperado (frequência observada = 36; frequência esperada = 38,9). As semanas quatro a sete
não apresentaram frequências observadas maiores do que o esperado (Figura 5A; média da
frequências observadas = 12,5; frequência esperada = 38,9).
Partilha Passiva versus Alimentação Independente:
Durante o procedimento experimental, os infantes podiam receber o alimento de um
cuidador, através de partilha passiva após um episódio de solicitação bem sucedida, ou
podiam se alimentar de modo independente, obtendo o item alimentar por conta própria
diretamente da fonte, evento considerado como consumo na fonte ou consumo fora da fonte
dependendo da distância entre a fonte de alimento e o local onde o item foi ingerido. Para
verificar qual a principal forma de obtenção de alimento pelos infantes de todas as famílias,
realizou-se a comparação entre a frequência dos itens recebidos através de partilha passiva
com a frequência dos alimentos obtidos por alimentação independente. Desse modo,
constatou-se diferença significativa, sendo a alimentação independente mais frequente (Figura
26 Análises por Família
Família E1:
Solicitação Bem Sucedida (Partilha Passiva):
Não houve evento de solicitação bem sucedida nesta família.
Solicitação Mal Sucedida (Ausência de Transferência):
Não foram observadas diferenças quanto ao sexo (Figura 6A; Teste de Wilcoxon = 11;
N = 20; p = 0,61) e status social (Figura 6B; Teste de Wilcoxon = 6; N = 20; p = 0,35) dos
animais que reagiram negativamente às solicitações de partilha por parte dos infantes.
Diferenças relacionadas à faixa etária não puderam ser observadas, pois o tamanho da
amostra foi insuficiente para a aplicação do teste estatístico.
Em relação à participação individual, não houve diferença significativa (Figura 6C;
Teste ANOVA de Friedman = 3,27; gl = 3; N = 20; p = 0,35).
Solicitação Bem Sucedida (Partilha Passiva) versus Solicitação Mal Sucedida
(Ausência de Transferência):
Não foi possível a realização desta análise, pois não ocorreu evento de solicitação bem
sucedida nesta família.
Proximidade da Fonte versus Distância da Fonte de Alimento:
Os infantes permaneceram significativamente mais distantes da fonte (Figura 7A;
Teste de Wilcoxon = 2; N = 32; p < 0,001).
Consumo na Fonte versus Consumo Fora da Fonte de Alimento:
O consumo fora da fonte foi mais frequente entre os infantes (Figura 7B; Teste de
27
Analisando-se a distribuição desses comportamentos ao longo das 20 primeiras
semanas de vida dos infantes, verificaram-se alguns episódios de consumo na fonte na 4ª
semana. No entanto, tais eventos foram mais frequentes entre a 10ª e a 20ª semana, período
em que também foi registrada a ocorrência de consumo fora da fonte (Figura 7C).
Tentativa de Roubo:
Entre a 10ª e a 18ª semana de vida, foram registrados seis episódios em que os filhotes
tentaram roubar itens alimentares em posse do macho adulto e do gêmeo.
Roubo:
Foi observado apenas um evento em que um dos infantes roubou o alimento em posse
do macho reprodutor, ocorrido na 17ª semana.
Ser Roubado:
Não ocorreram eventos em que os filhotes foram roubados nesta família.
Transporte:
Analisando-se a participação no transporte da prole quanto ao sexo dos cuidadores, os
machos foram os responsáveis por esse comportamento de cuidado aos infantes (Figura 8A;
Teste de Wilcoxon = 17; N = 18; p < 0,01).
Considerando-se o status social, os dominantes transportaram os infantes com maior
frequência (Figura 8B; Teste de Wilcoxon = 33,5; N = 18; p < 0,05).
Em relação à idade dos transportadores, não houve diferença significativa (Figura 8C;
Teste ANOVA de Friedman = 2; gl = 2; N = 2; p = 0,37).
Observou-se diferença significativa na participação individual quanto ao
comportamento de transporte da prole (Figura 8D; Teste ANOVA de Friedman = 19,48; gl =
28
significativamente mais do que os demais cuidadores adultos do grupo, com diferença
marginalmente significativa entre ele e o macho juvenil (p < 0,0125).
Neste grupo, o último registro de transporte da prole ocorreu na 9ª semana de vida
(Figura 8E).
Família E2
Solicitação Bem Sucedida (Partilha Passiva):
As análises relacionadas ao sexo, status social e idade dos animais que atenderam
positivamente às solicitações dos infantes não puderam ser realizadas, pois o tamanho da
amostra foi insuficiente para o emprego dos testes estatísticos.
Considerando-se a contribuição individual na solicitação bem sucedida, não houve
diferença significativa (Figura 9A; Teste ANOVA de Friedman = 7; gl = 7; N = 2; p = 0,43).
Vale ressaltar que, durante o período de estudo, ocorreu apenas um evento deste tipo e o
doador do alimento foi a fêmea juvenil.
Solicitação Mal Sucedida (Ausência de Transferência):
Analisando-se o sexo dos animais que não atenderam às solicitações dos infantes, não
houve diferença significativa (Figura 9B; Teste de Wilcoxon = 22; N = 18; p = 0,18).
Observa-se diferença significativa quanto ao status social (Figura 9C; Teste de
Wilcoxon = 0; N = 18; p < 0,01), de modo que os indivíduos subordinados foram os
responsáveis pelas solicitações mal-sucedidas dos infantes.
Em relação à classe etária dos envolvidos nas solicitações mal sucedidas, não foi
possível averiguar a ocorrência de diferenças, pois o tamanho da amostra foi insuficiente para
29
Observando-se a contribuição individual neste tipo de solicitação, houve diferença
significativa (Figura 9D; Teste ANOVA de Friedman = 26,07; gl = 7; N = 18; p < 0,001). No
entanto, o teste post-hoc não evidenciou diferença significativa em nenhum dos pares
analisados. Porém, no gráfico observa-se a tendência de que a fêmea juvenil e o macho
adulto1 apresentaram as maiores frequências de solicitação mal sucedida.
Solicitação Bem Sucedida (Partilha Passiva) versus Solicitação Mal Sucedida
(Ausência de Transferência):
As possíveis diferenças entre os tipos de solicitação não puderam ser comprovadas,
uma vez que o tamanho da amostra foi insuficiente para a aplicação do teste estatístico.
Proximidade da Fonte versus Distância da Fonte de Alimento:
Houve diferença significativa (Figura 10A; Teste de Wilcoxon = 5,50; N = 32; p <
0,001), sendo que os infantes permaneceram mais frequentemente distantes da fonte durante o
experimento.
Consumo na Fonte versus Consumo Fora da Fonte de Alimento:
Houve diferença significativa (Figura 10B; Teste de Wilcoxon = 0; N = 32; p < 0,001),
sendo o consumo fora da fonte a forma mais frequentemente observada entre os infantes.
Analisando-se a distribuição desses comportamentos ao longo das semanas de estudo,
observou-se que o consumo fora da fonte ocorreu entre a 10ª e a 20ª semana, enquanto o
consumo na fonte foi registrado entre a 15ª e a 20ª semana (Figura 10C).
Tentativa de Roubo:
Foram observados seis eventos em que os filhotes tentaram roubar o alimento em
posse dos machos adultos, macho juvenil e do gêmeo, ocorridos entre a 12ª e a 15ª semana de
30
Roubo:
Entre a 9ª e a 18ª semana, foram registrados quatro episódios em que os infantes
roubaram itens alimentares em posse da fêmea juvenil e do gêmeo.
Ser Roubado:
Ocorreram dois eventos em que os filhotes tiveram seus alimentos roubados. Tais
roubos foram efetuados pelo macho adulto 2 e pelo gêmeo, na 13ª e 18ª semana de vida,
respectivamente.
Transporte:
Considerando-se o comportamento de transporte dos infantes quanto ao sexo do
cuidador, houve diferença marginalmente significativa (Figura 11A; Teste de Wilcoxon = 55;
N = 22; p = 0,06). Percebe-se a tendência dos machos terem transportado mais
frequentemente a prole.
Em relação ao status social, houve diferença significativa (Figura 11B; Teste de
Wilcoxon = 10,50; N = 22; p < 0,001), sendo que os subordinados foram os principais
responsáveis pelo transporte dos infantes.
Não foi possível verificar a existência de diferenças relacionadas à idade dos
transportadores, porque o tamanho da amostra não foi suficiente para a realização do teste
estatístico.
Em relação à participação dos indivíduos, houve diferença significativa (Figura 11C;
Teste ANOVA de Friedman =25,43; gl = 7; N = 22; p < 0,001). O teste post-hoc revelou
diferença nos pares macho adulto1/macho dominante, macho adulto1/fêmea dominante,
macho adulto 1/fêmea juvenil, macho adulto 2/fêmea juvenil e fêmea adulta/fêmea juvenil.
Destes pares, os indivíduos que mais contribuíram no transporte dos infantes foram os machos
31
O último registro de transporte da prole foi efetuado na 11ª semana de vida (Figura
11D).
Família A1
Solicitação Bem Sucedida (Partilha Passiva):
Considerando-se o sexo, os machos foram os responsáveis pela maior parte das
solicitações bem-sucedidas (Figura 12A; Teste de Wilcoxon = 0; N = 22; p < 0,01).
Em relação ao status social, os dominantes responderam positivamente às solicitações
de alimento por parte dos filhotes com maior frequência (Figura 12B; Teste de Wilcoxon =
12,5; N = 22; p < 0,05).
Quanto à idade, houve diferença significativa (Figura 12C; Teste ANOVA de
Friedman = 6,9; gl = 2; N = 8; p < 0,05). O teste post-hoc mostrou que os subadultos
atenderam mais frequentemente às solicitações de alimento por parte dos infantes (p <
0,01666).
Analisando-se a participação individual na solicitação bem sucedida, houve diferença
significativa (Figura 12D; Teste ANOVA de Friedman = 22,96; gl = 3; N = 22; p < 0,01). O
teste post-hoc evidenciou diferença nos pares macho dominante/fêmea adulta, macho
dominante/macho juvenil, macho subadulto/fêmea adulta e macho subadulto/macho juvenil.
Destes pares, destacam-se o macho dominante e o macho subadulto com as maiores
freqüências de solicitações bem sucedidas (p < 0,0125).
Solicitação Mal Sucedida (Ausência de Transferência):
Observando-se o sexo dos envolvidos em solicitações mal sucedidas, constatou-se
diferença marginalmente significativa (Figura 13A; Teste de Wilcoxon = 28; N = 28; p =
0,07) e tendência dos machos terem apresentado as maiores frequências neste tipo de
32
Não houve diferença significativa quanto ao status social (Figura 13B; Teste de
Wilcoxon = 45,5; N = 28; p = 0,66) e à classe etária (Figura 13C; Teste ANOVA de Friedman
= 3,06; gl = 2; N = 12; p = 0,22).
Em relação à contribuição dos indivíduos, não houve diferença significativa (Figura
13D; Teste ANOVA de Friedman = 7,03; gl = 4; N = 4; p = 0,13), de modo que todos os
membros do grupo apresentaram frequências similares de respostas negativas às solicitações
de alimento por parte da prole.
Solicitação Bem Sucedida (Partilha Passiva) versus Solicitação Mal Sucedida
(Ausência de Transferência):
A solicitação bem sucedida foi mais frequente (Figura 14A; Teste de Wilcoxon = 0; N
= 22; p < 0,01).
Proximidade da Fonte versus Distância da Fonte de Alimento:
Não houve diferença significativa (Figura 14B; Teste de Wilcoxon = 178,50; N = 31;
p = 0,40).
Consumo na Fonte versus Consumo Fora da Fonte de Alimento:
Houve diferença significativa (Figura 14C; Teste de Wilcoxon = 13; N = 31, p < 0,01),
sendo a consumo na fonte a forma mais frequente de consumo pelos infantes.
Analisando-se a ocorrência desses comportamentos durante as 20 primeiras semanas
de vida da prole, observou-se que tanto o consumo na fonte quanto o consumo fora da fonte
ocorreram entre a 12ª e a 20ª semana (Figura 14D).
33
Foram registrados 18 eventos em que os filhotes tentaram obter itens alimentares por meio
de roubo, mas sem sucesso. Todos os membros do grupo, incluindo o gêmeo, foram alvo
dessas tentativas de roubo efetuadas pela prole entre a 7ª e a 14ª semana de vida.
Roubo:
Entre a 11ª e a 18ª semana, foram observados quatro episódios em que os infantes
roubaram alimento em posse do macho juvenil e do gêmeo.
Ser Roubado:
Ocorreram seis eventos em que os filhotes tiveram um item alimentar roubado pelos
machos adulto e juvenil, ocorridos entre a 14ª e a 20ª semana de vida da prole.
Transporte:
Houve diferença significativa em relação ao sexo dos transportadores (Figura 15A;
Teste de Wilcoxon = 2; N = 12; p < 0,01), tendo as fêmeas se destacado em tal atividade.
Não foi observada diferença quanto ao status social (Figura 15B; Teste de Wilcoxon =
29; N = 12; p = 0,72).
Analisando-se a faixa etária, houve diferença significativa (Figura 15C; Teste
ANOVA de Friedman = 14,97; gl = 2; N = 8; p < 0,001). O teste post-hoc mostrou diferença
nos pares adulto/subadulto e adulto/juvenil. Em ambos os pares, os adultos foram os
principais transportadores da prole (p < 0,01666).
Verificando-se a participação dos indivíduos no transporte dos filhotes, houve
diferença significativa (Figura 15D; Teste ANOVA de Friedman = 19,48; gl = 3; N = 18; p <
0,001). O teste post-hoc evidenciou diferença nos pares fêmea adulta/macho dominante,
fêmea adulta/macho subadulto, fêmea adulta/macho juvenil, macho dominante/macho
34
apresentou a maior frequência neste tipo de cuidado com os infantes, sendo seguida pelo
macho dominante (p < 0,0125).
Neste grupo, a coleta dos dados referentes ao transporte da prole iniciou-se apenas na
8ª semana de vida da prole. O último registro desse comportamento ocorreu na 13ª semana
(Figura 15E).
Família A2
Solicitação Bem Sucedida (Partilha Passiva):
As possíveis diferenças relacionadas ao sexo e ao status social dos animais que
atenderam às solicitações de alimento por parte dos filhotes não puderam ser averiguadas,
uma vez que o tamanho da amostra impossibilitou a aplicação do teste estatístico.
As diferenças quanto à classe etária não foram significativas (Teste ANOVA de
Friedman = 4; gl = 2; N = 2; p = 0,13). Vale ressaltar que nesta família ocorreram apenas dois
eventos de solicitação bem sucedida durante todo o período de estudo, ambos tendo a fêmea
dominante como o indivíduo doador do alimento. Sendo assim, as diferenças individuais
também não puderam ser verificadas, pois o tamanho da amostra foi insuficiente para a
utilização do teste estatístico.
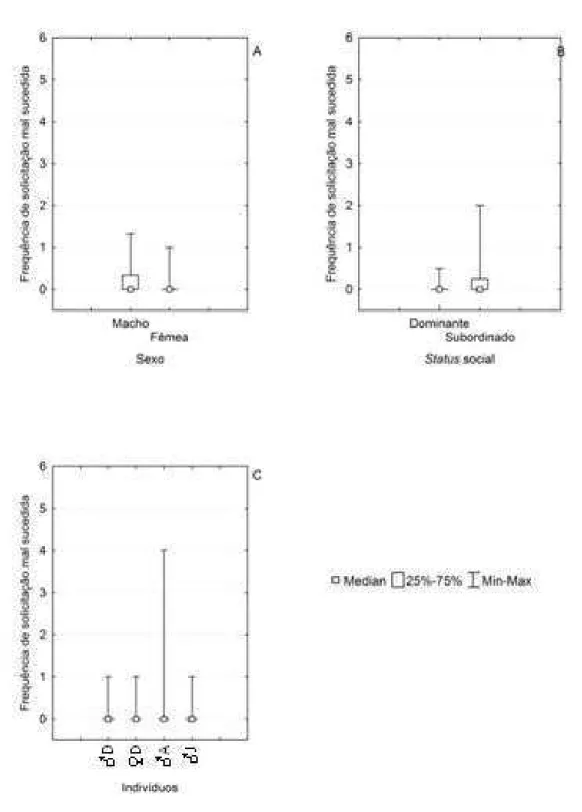
Solicitação Mal Sucedida (Ausência de Transferência):
Houve diferença significativa quanto ao sexo (Figura 16A; Teste de Wilcoxon = 3; N
= 11; p < 0,05) e ao status social (Figura 16B; Teste de Wilcoxon = 3; N = 11; p < 0,05) dos
animais que responderam negativamente às solicitações de alimento por parte do infante.
Dessa forma, as fêmeas e os indivíduos dominantes foram responsáveis pela maior parte das