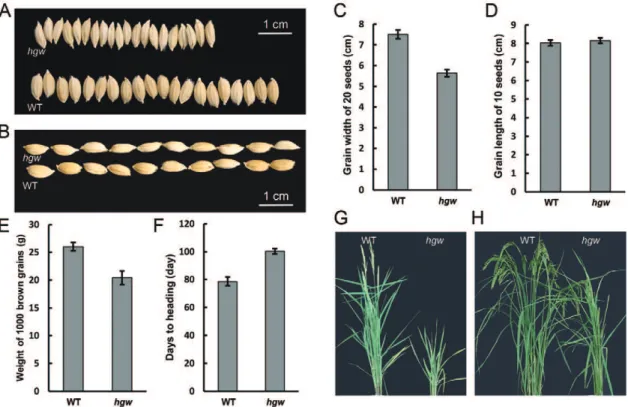
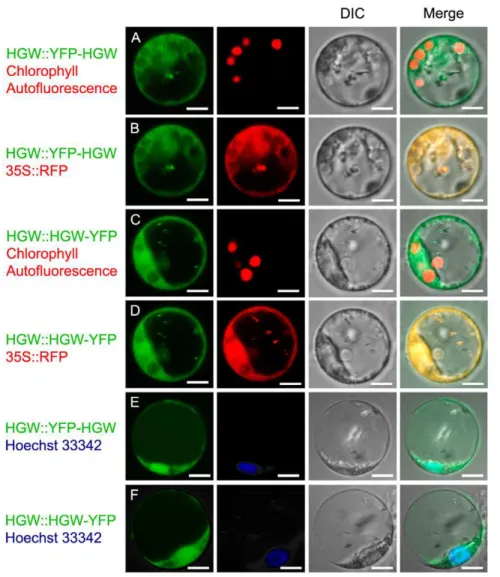
The rice HGW gene encodes a ubiquitin-associated (UBA) domain protein that regulates heading date and grain weight.
Texto
Imagem
Documentos relacionados
The mutant ∆XAC1491, whose knocked-out gene encodes a hypothetical protein, showed an approximately 100-fold reduced growth when compared to the wild strain, suggesting
The broad objective of the study is to analyze male and female access to land for cassava production in Abia state and specifically to describe the
(A) qRT-PCR analysis was performed to analyze SETD7 expression in 20 pairs of HCC tumor tissues and ANLTs, the data shown are the mean of – Δ CT, and the expression of SETD7 in HCC
The results of this study confirm that the tissue expression of the MGMT gene is reduced in neoplastic tissues of patients with sporadic CRC when compared to normal tissues..
Q-PCR analysis also indicated that Las titer in the Las-exposed psyllids (relative to that of the psyllid S20 ribosomal protein gene) was: 1) significantly higher, and increasing at
Comparative analysis of the expression levels of OsDR10 in different rice lines showed that this gene had a similar expression level in the three resistant rice lines (Figure
Using two distinct rice non-GH9 mutants and wild type, we performed integrative analysis of gene expression level by qRT-PCR, cellulase activities in situ and in vitro ,
qRT-PCR results showed that expression levels of miR-144, -153 and -340-5p in ipsilateral hippocampus were significantly elevated at all five time points after TBI, compared with
