Universidade de Trás-os-Montes e Alto Douro
Prevalência de ovotestis na gónada e alterações
histopatológicas no fígado em tainhas (fam. Mugilidae)
capturadas no estuário do Tejo
Dissertação de Mestrado em Biologia Clínica Laboratorial
Sofia Carina Lima Ribeiro
Orientador: João Soares Carrola
Co-orientador: Pedro Miguel Raposo de Almeida
II
Universidade de Trás-os-Montes e Alto Douro
Prevalência de ovotestis na gónada e alterações
histopatológicas no fígado em tainhas (fam. Mugilidae)
capturadas no estuário do Tejo
Dissertação de Mestrado em Biologia Clínica Laboratorial
Sofia Carina Lima Ribeiro
Orientador: João Soares Carrola
Co-orientador: Pedro Miguel Raposo de Almeida
III
Dissertação apresentada à Universidade de Trás-os-Montes e Alto Douro para cumprimento dos requisitos necessários à obtenção do grau de Mestre em Biologia Clinica Laboratorial. Dissertação realizada no laboratório de histologia no Departamento de Biologia e Ambiente (DeBA) sob a orientação científica do Professor Doutor João Soares Carrola e do Professor Doutor Pedro Miguel Raposo de Almeida.
A Sofia Carina Lima Ribeiro é a responsável pelas ideias apresentadas neste trabalho de dissertação de mestrado.
IV
Se queres chegar a onde a maioria não chega, faz o que a maioria não faz. Bill Gates
V
AGRADECIMENTOS
Na sequência da realização deste trabalho vários agradecimentos devem ser feitos. Em primeiro lugar gostaria de agradecer ao meu orientador, o professor João Soares Carrola, por ter aceitado orientar-me e me ter dado a oportunidade de trabalhar numa área que gosto e por ser um orientador competente e exigente – tenho a certeza que será uma mais-valia para mim no futuro. Quero agradecer também o seu empenho na logística das capturas e transporte dos peixes, bem como nas recolhas do material histológico, no apoio que foi fundamental na análise histopatológica (identificação e gradação das lesões observadas). Não posso deixar de realçar o rigor das correções deste trabalho, e também por toda a disponibilidade prestada ao longo da realização de todo o trabalho, paciências, e conhecimentos que me transmitiu durante a realização e escrita da dissertação, incluindo ensinamentos desde a escrita, até ao desenho vetorial para fazer o mapa e esquemas, sem esquecer a formação para a utilização do programa informático Endnote, e ainda por me ter convidado a colaborar nalgumas aulas práticas relacionadas com esta temática.
Ao Professor Pedro Raposo de Almeida, docente na Universidade de Évora e investigador no MARE-UÉvora (Centro de Ciências do Mar e do Ambiente) por ter aceitado ser o co-orientador desta dissertação, pelo apoio, disponibilidade e empenho na revisão da mesma. Quero ainda agradecer a ajuda e apoio por parte da sua equipa na captura e transporte dos peixes, bem como no apoio na recolha do material histológico.
Á Sílvia Pedro investigadora no MARE-UÉvora, pela ajuda e empenho na parte prática de campo, e que foi fundamental para o avançar deste trabalho, desde apoio na logística, recolha das amostras e transporte dos peixes.
Agradecer ainda ao Filipe Ribeiro pela colaboração e ajuda nas recolhas e aos demais voluntários da Universidade de Lisboa e de outras Universidades, por terem colaborado nas recolhas e demais tarefas relacionadas.
Às funcionárias do laboratório de histologia do Departamento de Biologia e Ambiente (DeBA) da UTAD, em particular à Ana Fraga e à técnica Donzília Costa, quero expressar a minha gratidão pela disponibilidade e explicação para a preparação do material histológico. Desejo agradecer, também, à técnica Cesaltina Carvalho o apoio que me deu, a ajuda e empenho que demonstrou na preparação do material para a recolha das amostras nas duas saídas de campo que forma realizadas, bem como a resolver situações um pouco chatas, mesmo não estando envolvida diretamente no trabalho realizado.
VI
Ao André Pires pelo apoio na análise estatística dos dados obtidos.
À Ana Luzio por todo o seu apoio prestado e disponibilidade demonstrada ao longo dos dois anos e à Dércia Santos pela ajuda na preparação da apresentação final da dissertação.
Aos meus colegas e amigos, Paulo Jorge Especial e Hugo Neves, pela ajuda, amizade, conversas e companhia nos intervalos de trabalho.
A nível pessoal não posso deixar de agradecer:
- ao Doutor Carlos Ferreira pelo seu excelente trabalho na melhoria do meu bem-estar;
- á Marta Teixeira, Maria Silva e Cátia Santos pelos seus conselhos, paciência, amizade e momentos de descontração e à Janeth Vieira da Silva que, mesmo à distância, apoiou-me nesta etapa;
- à Sandrina Teixeira, Gisela Alves, Catarina Costa, Cristina Ferreira, Flávia Cardoso, Daniela Alves, Vânia Silva, Bruno Oliveira, Jorge Rafael, Liliana Mota, Joana Teles, Pedro Joel, Rosa Mendes, Vilma, Isabel Carvalho, Carolina Santos, Catarina, Rita, Suzana Parra, Bruna Costa e Leila Salta por me aturarem.
E por último, aos meus pais, José e Maria, pelos conselhos, atenção, puxões de orelhas e apoio incondicional ao longo deste tempo e à minha irmã, Manuela, pela sua capacidade orientadora.
VII
RESUMO
A poluição dos ecossistemas aquáticos, nomeadamente dos estuários, tem origem em diversas atividades antropogénicas e pode afetar os organismos aquáticos, incluindo os peixes. Estes podem ser utilizados como uma fonte de diversos biomarcadores, em particular os histopatológicos para avaliar o efeito da poluição.
Em 2014 foram capturadas 100 tainhas (59 tainhas em fevereiro e 41 em novembro - 40 Liza
aurata, 59 Chelon labrosus e um Liza ramada), das quais apenas 29% (30/100) eram machos.
Com este trabalho pretendemos identificar possíveis oócitos na gónada masculina fazendo a determinação da sua prevalência e severidade utilizando o Índice de Severidade de Ovotestis (ISO) com base na metodologia de Bateman et al., (2004). Por outro lado, fazer a identificação das principais lesões e alterações hepáticas e sua avaliação semi-quantitativa utilizando uma gradação de 0 a 7 (valor máximo) segundo metodologia de Carrola (2003).
Apenas 3% (1/30) dos peixes macho (Chelon labrosus) apresentava baixa densidade de oócitos - 3 oócitos primários, originando um baixo valor de ISO (±0,04) o qual corresponde à
classe 1. Embora não tenham sido detetadas quaisquer macrolesões (externas e internas), após
uma análise histológica ao fígado de 60 tainhas (das quais 30 capturadas em fevereiro e 30 em novembro) observaram-se algumas lesões e alterações hepáticas, sendo que as mais observadas foram, agregados macrofágicos, focos linfóciticos, necrose, focos inflamatórios e granulomas inespecificos, e as que apresentaram maior grau de frequência e severidade foram, os agregados macrofágicos, granulomas inespecificos e focos linfóciticos.
Os dados obtidos neste estudo mostram baixa prevalência e muito baixa severidade de ovotestis refletindo uma baixa exposição a CDE principalmente pela via alimentar, apesar de existirem quantidades consideráveis de CDE neste estuário. Isto deve-se à redução considerável de partículas em suspensão à saída dos esgotos relacionada com as ETARs deixando de ter alimento disponível.
Também foi observada uma baixa prevalência e severidade de lesões e alterações hepáticas o que indica que os peixes estiveram expostos a níveis baixos de poluição. Podemos ainda concluir que os peixes analisados não estiveram expostos ao longo da sua vida a quantidades suficientes de desreguladores endócrinos, que induzissem o aparecimento da condição de intersexo – ovotestis, ou de outros tóxicos que pudessem levar a efeitos hepatotóxicos.
VIII
ABSTRACT
The pollution of aquatic ecosystems, including estuaries, comes from various anthropogenic activities and may affect aquatic organisms, including fish. These may be used as a source of various biomarkers, in particular histopathological biomarkers, to evaluate the effect of pollution. In 2014, 100 mullets were captured (59 mullets in february and 41 in november - 40 Liza
aurata, 59 Chelon labrosus and one Liza ramada), of which only 29% (30/100) were males.
The aims of this work is identify possible oocytes in male gonads making the determination of its prevalence and severity using the Ovotestis Severity Index (OSI) based on the methodology of Bateman et al., (2004). On the other hand, make the identification of main hepatic lesions and alterations, and its semi-quantitative evaluation using a gradient from 0 to 7 (maximum value) using Carrola (2003) methodology. Only 3% (1/30) of male fish (Chelon labrosus) had low density oocytes - 3 primary oocytes, resulting in a low OSI value (± 0.04) which corresponds to class 1. Although not detected any macrolesions (external and internal) after a histological analysis of the 60 mullets liver (30 captured in february and 30 in november) we observed some liver lesions and alterations and the most observed were, macrophage aggregates, lymphocytic foci, necrosis, inflammatory foci and nonspecific granulomas, and those with the greatest degree of frequency and severity were the macrophage aggregates, nonspecific granulomas and lymphocytic foci.
Our data show low prevalence and very low severity of ovotestis reflecting a low exposure to EDC mainly through food, though there are considerable amounts of EDC in this estuary. This is due to the considerable reduction of particles in suspension at the outlet of sewage related to the WWTP allowing food to be available.
Also a low prevalence and severity of liver lesions and alterations indicating that the fish were exposed to low levels of pollution. We can still conclude that the analyzed fish have not been exposed throughout their lives to sufficient quantities of EDC, which induce the appearance of intersex condition - ovotestis, or other toxic that could lead to hepatotoxic effects.
IX ÍNDICE GERAL AGRADECIMENTOS ... V RESUMO ... VII ABSTRACT ... VIII ÍNDICE GERAL ... IX ÍNDICE DE FIGURAS ... XI ÍNDICE DE TABELAS ... XIII
ÍNDICE DE GRÁFICOS ... XIII LISTA DE ABREVIATURAS ... XIV
1. INTRODUÇÃO GERAL ... 1
2. MONITORIZAÇÃO E BIOMONITORIZAÇÃO AMBIENTAL ... 5
2.1. Bioindicadores ... 7
2.1.1. Peixes como bioindicadores da qualidade do meio aquático ... 8
2.2. Biomarcadores e sua importância em estudos de monitorização ambiental ... 9
2.3. Sistema endócrino da reprodução dos peixes ... 11
2.4. Macrolesões ou “Gross lesions” ... 12
2.5. Compostos desreguladores endócrinos (CDE) ... 13
2.5.1. Efeitos provocados nos peixes ... 15
3. GÓNADA MASCULINA - TESTÍCULO ... 17
3.1. Histologia da gónada masculina ... 18
3.2. Condição de intersexo como biomarcador ... 19
4. TECIDO HEPÁTICO E HISTOPATOLOGIA... 21
4.1. Histologia do fígado de peixe ... 23
4.2. Histopatologia hepática ... 24
X
6. METODOLOGIA ... 29
6.1. Área de estudo ... 30
6.1.1. Caracterização da L. ramada, C. labrosus e L. aurata ... 31
6.1.2. Morfologia da L. ramada, C. labrosus e L. aurata ... 32
6.1.3. Sistema reprodutor da L. ramada, C. labrosus e L. aurata ... 32
6.1.4. Alimentação da L. ramada, C. labrosus e L. aurata ... 33
6.2. Captura dos peixes e recolha das amostras ... 33
6.3. Descrição resumida da técnica histológica... 34
6.4. Índices somáticos (Gross Indices) ... 35
6.4.1. Fator de condição corporal (FCC) ... 35
6.4.2. Índice gonadossomático (IGS) ... 35
6.4.3. Índice hepatossomático (IHS) ... 36
6.5. Índice de severidade de ovotestis (ISO) ... 36
6.6. Gradação das lesões e alterações hepáticas ... 37
6.7. Análise estatística ... 38
7. RESULTADOS ... 39
7.1. Dados morfométricos ... 40
7.2. Análise histológica das gónadas masculinas – testículo ... 41
7.3. Análise histopatológica do fígado ... 42
8. DISCUSSÃO ... 47
8.1. Alterações na histologia das gónadas masculinas ... 48
8.2. Histopatologia hepática ... 51
9. CONCLUSÕES ... 53
XI
ÍNDICE DE FIGURAS
Figura 1. Esquema da indução de vitelogenina controlada pelo sistema endócrino em peixes fêmea (adaptado de: http://mn.water.usgs.gov/projects/CED/summary.html). ... 12 Figura 2. Esquema com alguns mecanismos de disfunção endócrina (adaptado de Ghiselli e Jardim, 2007). A) As hormonas naturais encaixam-se perfeitamente nos recetores e transmitem os sinais indispensáveis às células. B) Os compostos desreguladores endócrinos ocupam o lugar das hormonas naturais encaixando-se perfeitamente nos recetores, enviando sinais diferentes e fora de tempo às células e C) E por fim, os desreguladores endócrinos atuam como bloqueadores dos sinais normais das hormonas naturais que seriam enviados às células. ... 14
Figura 3. Microfotografias do tecido das gónadas masculinas das solhas capturadas no estuário do Douro em 2013. Gónada masculina imatura (A) e gónada masculina madura (B) (H&E). A) Presença de espermatozóides em fase de desenvolvimento (seta preta) e aglomerados de espermatócitos (seta branca), barra = 100 µm; B) Microfotografia do tecido testicular de solha, onde se podem observar cortes de tubos seminíferos com espermatozóides (seta preta), tecido conjuntivo intersticial (asteriscos), bem como aglomerados de espermatócitos (seta branca), barra = 100 µm... 19
Figura 4. Microfotografia do fígado (corte transversal) de tainha capturada no estuário do Tejo (H&E) onde estão representados ductos biliares (setas brancas) sendo que dois deles estão junto a agregados macrofágicos (setas pretas), barra = 200 μm. ... 23
Figura 5. Representação geográfica do estuário do Tejo e do local amostragem (realizado por Sofia Ribeiro segundo a metodologia de Carrola (2011). ... 30
Figura 6. Fotografia da tainha fataça (L. ramada) (disponível em: http://fineartamerica.com/featured/thinlip-mullet-liza-ramada-mulet-morragute-cefalo-tainhatunnlaeppad-multe-roendungur-urft-valley-art.html). ... 31
Figura 7. Ilustração da tainha liça (C. labrosus) (disponivel em: http://www.zeevissers.com/zeevissoorten.html). ... 31
Figura 8. Ilustração da tainha garrento (L. aurata) (disponível em: http://w3.ualg.pt/~madias/guadiana/garrento.html). ... 32
XII
Figura 9. Microfotografias do tecido das gónadas masculinas maduras (A, B e C) (corte transversal) das tainhas capturadas no estuário do Tejo (H&E). A) Gónada diferenciada com presença de espermatozóides (seta branca), barra = 500 µm; B) Ampliação da microfotografia A) onde podemos observar espermatozóides (seta branca) e tecido conjuntivo intersticial (asterisco), barra = 200 µm, C) Nesta microfotografia podemos observar com mais detalhe cortes de tubos seminíferos com espermatozóides (seta branca) e tecido conjuntivo intersticial (asteriscos), barra = 100 µm e D) Microfotografia de 3 oócitos primários distribuídos pelo parênquima apresentando pequena dimensão e com morfologia algo anormal... 42
Figura 10. Microfotografias de secções de fígado das tainhas capturadas no estuário do Tejo (H&E). A) agregado macrofágico (seta preta), granuloma parasitário (seta roxa) e vacuolização hidrópica (asterisco), barra = 100 μm; B) presença de picnose (asterisco) e necrose (seta preta) no tecido hepático, barra = 100 μm; C) picnoses perivasculares (seta preta) junto a um agregado macrofágico (seta branca), barra = 200 μm; D)vacuolização (asterisco), barra = 200 μm; E) foco de células claras (tracejado preto), barra = 200 μm e F) presença de um parasita (seta preta) junto a um agregado macrofágico (tracejado preto) no tecido hepático, barra = 500 μm. ... 44
Figura 11. A) foco de linfócitos (asterisco) junto a um agregado macrofágico (seta preta) e a 2 ductos biliares (seta branca), barra = 200 μm; B) necrose perivascular (seta preta), barra = 500 μm; C) focos inflamatórios (setas pretas) e granulomas inespecificos (setas brancas), barra = 200 μm e D) presença de 2 granulomas inespecificos no parênquima hepático (seta branca), barra = 200 μm. ... 45
XIII
ÍNDICE DE TABELAS
Tabela 1. Principais alterações hepáticas em pleuronectiformes e sistema de classificação usado para a sua quantificação na solha-das-pedras (Lang et al., 2006) ... 25
Tabela 2. Número de tainhas capturadas (n) em fevereiro e novembro de 2014. ... 30
Tabela 3. Tabela com o sistema de classes desenvolvido por Bateman et al. (2004) ... 37
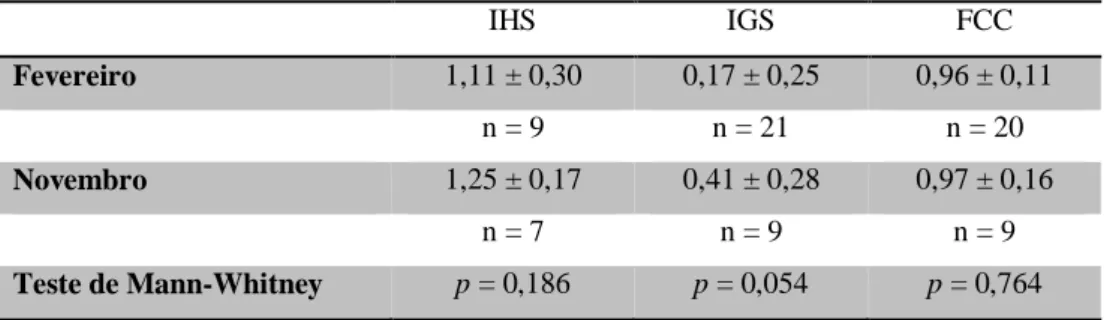
Tabela 4. Análise estatística dos valores de IHS, IGS e FCC utilizando o Teste de Mann-Whitney dos peixes macho capturados (n) em fevereiro e novembro de 2014 no estuário do Tejo e onde p representa o valor de significância. ... 40
Tabela 5. Análise estatística dos valores de IHS, IGS e FCC utilizando o Teste de Mann-Whitney dos peixes fêmea capturadas (n) em fevereiro e novembro de 2014 no estuário do Tejo e onde p representa o valor de significância. ... 41
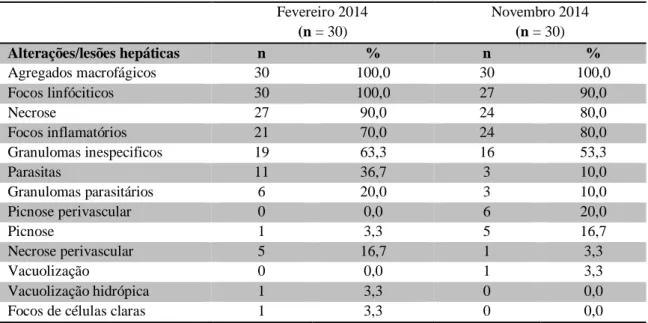
Tabela 6. Alterações/lesões hepáticas observadas nos fígados dos peixes, onde n é o número de fígados analisados e % é a percentagem da alteração e lesão hepática observada relativamente ao número de fígados analisados. ... 43
Tabela 7. Média e erro padrão da gradação feita para cada alteração e lesão hepática, onde n é o número de fígados de tainhas analisadas e p o valor de significância no Teste de Mann-Whitney. ... 46
ÍNDICE DE GRÁFICOS
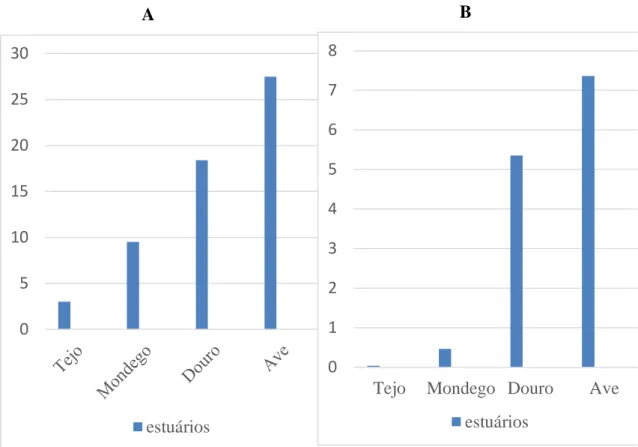
Gráfico 1. A) Percentagem de machos que apresentavam ovotestis no estuário do (Tejo <<Mondego<Douro<Ave) e B) Valor de ISO quando comparados com os dados do Tejo com os dados de Carrola (2011) relativos ao estuário Mondego, Douro e Ave. ... 49
XIV
LISTA DE ABREVIATURAS
AAE - Agência Ambiental Europeia AM - Agregados macrofágicos AP - Alquilfenól
BPA - Bisfenol A
BPC - Bifenilo policlorado
CDE - Compostos desreguladores endócrinos CT – Comprimento total
DA – Dopamina
DDT - Diclorodifeniltricloroetano DQA - Diretiva Quadro da Água
ETAR - Estações de Tratamento de Águas Residuais FCC - Fator de condição corporal
GnRH – Hormona libertadora de Gonadotrofinas GtHI - Gonadotrofina I
GtHII - Gonadotrofina II
HAP - Hidrocarboneto aromático policíclico IGS - Índice gonadossomático
IHS - Índice hepatossomático
ISO - Índice de severidade de ovotestis MBA - Monitorização da bioacumulação MEB - Monitorização do efeito biológico MQ - Monitorização química
MS - Monitorização da saúde
OMS - Organização Mundial da Saúde
PANU - Programa Ambiental das Nações Unidas PT – Peso total
RE - Recetores de estrogénio TBT - Tributilestanho VTG - Vitelogenina
1
1. INTRODUÇÃO GERAL
2 INTRODUÇÃO GERAL
Nas últimas décadas, o ser humano introduziu centenas de novos contaminantes, devido à elevada atividade industrial, intensificação da agricultura e aumento dos resíduos urbanos no ambiente, os quais podem acabar nos ecossistemas aquáticos. Estas fontes de poluição devem ser identificadas rapidamente (Cheng e Jia, 2010), e quanto mais rápido for a sua identificação mais fácil será implementar medidas de mitigação do impacto e evitar efeitos nefastos em vários níveis da organização biológica (Lins et al., 2010).
A maioria dos contaminantes são subprodutos de processos de fabrico que podem entrar no ecossistema aquático intencionalmente, como por exemplo, através de efluentes urbanos (Nyenje et al., 2010), ou de forma não intencional, como consequência de acidentes ambientais ou derrames (Naigaga et al., 2011), pela contaminação do solo – devido ao uso excessivo de pesticidas - através das estações de tratamento de águas residuais – ETAR (Reinen et al., 2010), por escoamentos superficiais (Kibena et al., 2014) e subterrâneos derivados de atividades do setor primário, como a agricultura (Duncan, 2014; Zadorozhnaya et al., 2014; Su et al., 2013; Goldar e Banerjee, 2004; Schock, 2000).
Existe uma elevada variedade de contaminantes presentes nos ecossistemas aquáticos, tais como, bifenilos policlorados (BPC) (Tomasallo et al., 2010; Ozmen et al., 2008), metais (Koukal et al., 2004; Santschi et al., 2001), hidrocarbonetos aromáticos policíclicos (HAP) (Ozmen et al., 2008), fármacos (Madureira et al., 2010; Heberer, 2002) e produtos de uso pessoal (Dai et al., 2015; Brausch e Rand, 2011; Pojana et al., 2007), tais como cosméticos e agentes de proteção solar (Dai et al., 2015). Devido à elevada contaminação deste, ocorre uma diminuição da capacidade deste de autodepuração do ecossistema aquático (Rautenberg et al., 2015) aumentando a perturbação dos organismos (Propper, 2005).
Alguns novos contaminantes começaram a ser detetados, identificados e definidos quanto aos riscos que possam causar no meio ambiente, é o caso dos chamados “contaminantes emergentes”. A classificação deste tipo de contaminante divide-se em três grupos principais: desreguladores endócrinos, produtos de uso pessoal e fármacos (Jasinska et al., 2015). A determinação dos níveis dos contaminantes é realizada através de análises químicas (Ramalhosa et al., 2005), e a avaliação dos seus efeitos pode ser determinada utilizando bioindicadores e recorrendo a biomarcadores (Carrola, 2011).
Introdução Geral
3
Os estuários são ecossistemas complexos e dinâmicos que exercem um papel fulcral no equilíbrio da biosfera. São locais de transição entre o ambiente marinho e o de água doce (Sun et al., 2012) e fornecem alimento e habitat a organismos bênticos, epi-bênticos e pelágicos. Os estuários são locais importantes de reprodução e de maternidade para determinadas espécies de peixes, fornecem também proteção contra predadores devido às vastas zonas de baixa profundidade, incluindo sapais, localizadas no seu interior.
Deste modo, os critérios de avaliação da condição ecológica dos ecossistemas aquáticos, particularmente os estuários, incluem bioindicadores e biomarcadores. É neste âmbito que se utilizam os peixes como instrumento de biomonitorização para a avaliação da qualidade da água em termos de contaminação por poluentes, bem como das características ecológicas destes ecossistemas.
5
2. MONITORIZAÇÃO E
BIOMONITORIZAÇÃO AMBIENTAL
6 MONITORIZAÇÃO E BIOMONITORIZAÇÃO AMBIENTAL
Tendo em conta a diversidade de compostos químicos e seus efeitos é importante fazer a sua identificação e quantificação atempada. Sendo assim a monitorização ambiental é uma ferramenta importante, e que permite fazer o estudo de determinadas variáveis ao longo do tempo e do espaço, com intervalos definidos (em função do tipo de estudo) e que serve para estimar a evolução das variáveis, avaliar a exposição e o risco para um organismo quando comparada com um ponto de referência. Pode ser utilizada para quantificação de um ou mais parâmetros físicos ou químicos, de acordo com um cronograma previamente estipulado e utilizando métodos comparáveis e padronizados, em função do tipo de estudo a realizar, podendo seguir as indicações do Programa Ambiental das Nações Unidas – PANU, ou ainda da Agência Ambiental Europeia - AAE (van der Oost et al., 2003).
Podemos dividir a monitorização ambiental em cinco áreas, a fim de avaliar os riscos dos contaminantes para os organismos, incluindo os peixes, e classificar a qualidade ambiental dos ecossistemas e estado ecológico dos mesmos, tal como definido pela Diretiva Quadro da Água (DQA), 2000/60/CE, nomeadamente:
monitorização química (MQ) - avaliação da exposição, por medição dos níveis de um conjunto selecionado de contaminantes conhecidos no meio ambiente abiótico;
monitorização da bioacumulação (MBA) - avaliação da exposição medindo os níveis de contaminantes na biota ou determinando a dose crítica no local de interesse (bioacumulação);
monitorização do efeito biológico (MEB) - avaliação da exposição e do efeito determinando as primeiras alterações adversas que são parciais ou totalmente reversíveis (biomarcadores);
monitorização da saúde (MS) - Avaliação do efeito através da ocorrência de doenças irreversíveis ou danos nos tecidos dos organismos;
monitorização dos ecossistemas (ME) - avaliação da integridade de um ecossistema, fazendo um inventário, por exemplo da composição, densidade e diversidade das espécies, entre outros.
Monitorização e Biomonitorização Ambiental
7
A utilização regular e sistemática de organismos para avaliar a qualidade da água, como na MBA, MEB, MS e ME, denomina-se de monitorização biológica (MB) ou biomonitorização (van der Oost et al., 2003).
Quando comparada com as vantagens das análises físico-químicas convencionais a biomonitorização pode mostrar a exposição e/ou os efeitos que ocorrem nos organismos afetados por tóxicos ou misturas de tóxicos, refletindo indiretamente a qualidade ecológica geral do ecossistema em que habitam (Iliopoulou-Georgudaki et al., 2003).
Uma das vantagens das análises físico-químicas é que permitem detetar a presença e quantificar a concentração de um contaminante e caracterizar o nível de contaminação dos sedimentos ou da água (Carrola, 2003). No entanto podem apresentar algumas desvantagens, como por exemplo, não traduzem o que está a acontecer num determinado ecossistema aquático, ou os efeitos num organismo vivo, isto é, qual o tipo de toxicidade que está associada aos valores de contaminantes medidos e tendo em conta as diferenças de sensibilidade entre espécies e as condições que podem interferir, não permitem determinar os efeitos cumulativos dos contaminantes, relacionados com o historial de poluição do local ou área (Pinto, 2007).
Podemos resumir e afirmar que o diagnóstico ambiental completo necessita de utilizar ambas as abordagens, monitorização e biomonitorização. Os peixes são organismos importantes para a biomonitorização pois são sensíveis a mudanças que possam ocorrer no ecossistema aquático, através de alterações nas suas respostas biológicas, bioquímicas, morfológicas e comportamentais mesmo com baixos níveis de poluição (Ballesteros et
al., 2009; Das e Chakrabarty, 2007).
2.1. Bioindicadores
Um bioindicador ou indicador biológico corresponde a um determinado organismo selecionado com o objetivo de fornecer informação sobre as condições ambientais de um habitat, pela sua presença ou ausência, pela evolução do número de indivíduos, densidade e riqueza de espécies e tipo de espécie presente (autóctones ou invasoras) (van der Oost
et al., 2003).
Os bioindicadores podem ser agrupados em: bioindicadores de acumulação ou cumulativos, quando um determinado organismo acumula influências antrópicas, sem
8
mostrar danos passíveis de serem reconhecidos num curto espaço de tempo, e bioindicadores sensíveis, caso responda diretamente ao stresse ambiental causado pelos contaminantes.
A DQA dá grande relevância aos bioindicadores, porque são um complemento útil à monitorização permitindo definir o estado ecológico e a qualidade dos ecossistemas onde os bioindicadores habitam (Ceschin et al., 2012) e integram os efeitos de vários agentes de stress fornecendo uma ferramenta para avaliação do seu impacto (Iliopoulou-Georgudaki et al., 2003).
Um exemplo de um bom indicador da qualidade do ar são os líquenes epífitas pois tem um período de vida longo e um crescimento lento estando por isso expostos aos contaminantes presentes na atmosfera, por um longo período de tempo e visto não possuírem cutícula acabam por os absorver mais facilmente (Garty, 2001).
Relativamente aos ecossistemas aquáticos os bioindicadores frequentemente usados são os invertebrados aquáticos e a fauna piscícola, os quais podem ser estudados em conjunto (Pinto et al., 2010). São afetados por fatores químicos (pH, salinidade, e xenobióticos), físicos (temperatura, condutividade elétrica e turbidez), geo-morfológicas (área, densidade de drenagem e perímetro da bacia) e antrópicos (Testi et al., 2009). Estão também expostos a outros fatores naturais de stresse, como, variação nos níveis de nutrientes e hipóxia (Marsden e Baharuddin, 2014) e à introdução e impacto das espécies exóticas invasoras, perturbando as populações de espécies autóctones.
2.1.1. Peixes como bioindicadores da qualidade do meio aquático
Os peixes são considerados bioindicadores úteis porque existem várias espécies de peixes que representam os diferentes níveis tróficos (omnívoros, herbívoros, insetívoros, piscívoros entre outros), e por isso as espécies de níveis tróficos superiores tendem a integrar os efeitos dos níveis tróficos inferiores, tornando-se por isso o reflexo da qualidade dos ecossistemas aquáticos (Freitas e Siqueira-Souza, 2009).
Por outro lado, como são consumidos pelo Homem constituem um risco potencial para a saúde humana, devendo ser considerados nos estudos de toxicologia alimentar (Zhou et al., 2008). Por exemplo, no Bangladesh a água subterrânea está altamente contaminada por arsénico e por outros metais, porém é muito utilizada para cultura de
Monitorização e Biomonitorização Ambiental
9
peixes que são consumidos pelo ser humano, como resultado, os peixes consumidos a partir de água contaminada aumentam os riscos para a saúde humana (Ahmed et al., 2015).
Os peixes também tem sido identificados como um dos mais utilizados bioindicadores na avaliação da qualidade do ecossistema aquático, devido à sua ampla distribuição e fácil identificação (Karr et al., 1986) no entanto, a mobilidade de muitas espécies faz com que seja difícil identificar não só a fonte exata de poluição, mas também o tempo e duração da exposição (Chovanec et al., 2003). Os peixes são também utilizados como organismos sentinela, pois desempenham diversas funções na teia trófica, acumulam substâncias tóxicas nos tecidos, e respondem mesmo a baixas concentrações dos mesmos (Stegeman, 2000).
As tainhas são bons bioindicadores da qualidade do meio aquático porque geralmente alimentam-se junto às descargas de esgotos (principalmente urbanos), ingerindo, deste modo, os detritos potencialmente carregados de numerosos contaminantes, como diversos metabólitos provenientes dos fármacos utilizados pelas pessoas (Proença, 2011), para além de outros compostos derivados do uso de produtos de higiene e de uso pessoal (Brausch e Rand, 2011). Elas também toleram bem a salinidade extrema, são abundantes em sistemas estuarinos da Europa, e portanto, podem ser uma boa ferramenta para programas de monitorização (Ferreira et al., 2005).
2.2. Biomarcadores e sua importância em estudos de monitorização ambiental
Os biomarcadores são uma ferramenta complementar e valiosa na biomonitorização. Um biomarcador corresponde a uma resposta biológica associada à exposição de um organismo vivo a um produto químico tóxico (ou mistura de tóxicos) a nível individual, medida ou avaliada no organismo (bioquímica, fisiologia, histopatologia, etc.) ou nos seus produtos (sangue, bílis, urina, etc.), indicando um desvio ao estado homeostático normal - controlo ou referência (van der Oost et al., 2003). Os biomarcadores indicam a exposição aos contaminantes e/ou os efeitos resultantes dessa exposição, e são cada vez mais usados para avaliar o estado ecológico dos ecossistemas de água doce e marinhos (Liu et al., 2015; Porte et al., 2006).
10
Alguns biomarcadores usados em peixes incluem: alterações na histologia das gónadas e fígado, alterações na concentração de hormonas (hormonas esteróides, hormonas da tiroide), desenvolvimento anormal das gónadas, baixa viabilidade dos gâmetas, alterações na atividade enzimática - i.e. aromatases - (Mills et al., 2014) e de vitelogenina (Kleinkauf et al., 2004).
Existem vários tipos de biomarcadores: moleculares, bioquímicos, citológicos, histológicos, comportamentais, etc. e estão divididos em três categorias, biomarcadores de exposição, efeito e sensibilidade (Sarkar et al., 2006).
De acordo com a Organização Mundial de Saúde (OMS) (1993), os biomarcadores podem ser subdivididos em três classes:
Biomarcadores de exposição - compreendem a deteção e medição de uma substância exógena, ou de um seu metabolito ou produto de uma interação entre um agente xenobiótico e determinada molécula alvo ou célula, que pode ser mensurável numa parte de um organismo (Jesus e Carvalho, 2008);
Biomarcadores de efeito - incluem alterações bioquímicas, fisiológicas e histopatológicas mensuráveis em tecidos ou fluídos corporais de um organismo, podendo estar associados a uma possível disfunção (Feist et al., 2015);
Biomarcadores de suscetibilidade - indicam a capacidade inerente ou adquirida de um organismo para responder a mudanças de exposição a uma substância xenobiótica específica, incluindo fatores genéticos e alterações nos recetores que podem alterar a sensibilidade de um organismo à referida exposição (Ciocan et
al., 2015).
Alguns biomarcadores são inespecificos, ou seja, nenhum biomarcador pode fornecer todas as informações sobre a exposição ou os efeitos da poluição presente no ambiente, por isso, na maioria das avaliações ambientais, são usados diferentes biomarcadores de forma a obter uma avaliação mais completa na biomonitorização (Carrola, 2011).
Monitorização e Biomonitorização Ambiental
11 2.3.Sistema endócrino da reprodução dos peixes
O sistema endócrino é constituído por glândulas e tecidos que libertam substâncias químicas – hormonas - responsáveis pelo controlo da maioria das funções biológicas.
A ação das hormonas é essencial para o desenvolvimento correto das gónadas (ovários e testículos) dos peixes (Almeida, 2013). Na maioria das espécies de peixes estudadas até à data, o estrógeno (E2) é a principal hormona envolvida na diferenciação sexual, pois dela depende a formação dos ovários nas fêmeas, enquanto que os andrógenos são uma consequência da diferenciação dos testículos (Guiguen et al., 2010).
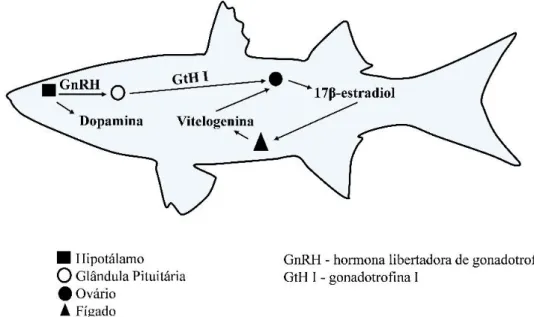
O hipotálamo produz, entre outros, a hormona libertadora de gonadotrofinas (GnRH) e a dopamina (DA) (Figura 1). A GnRH estimula a libertação das gonadotrofinas pela adeno-hipófise, enquanto a DA inibe a libertação destas, de modo direto ou indireto (inibindo o efeito estimulador da GnRH). As gonadotrofinas estimulam a maturação gonadal e a libertação de esteróides das gónadas. Muitos dos processos relacionados com a reprodução envolvem mudanças na secreção da GnRH e alterações dos efeitos da glândula pituitária. Nos teleósteos, existem duas gonadotrofinas: gonadotrofina I (GtH I) e gonadotrofina II (GtH II). Nas fêmeas, a GtH I estimula o crescimento das gónadas, a gametogénese e a entrada de vitelogenina (VTG) nos oócitos (Almeida, 2013) enquanto que a GtH II é importante na maturação dos oócitos e na desova (Moncaut et al., 2003). Os níveis de GtH I são elevados durante as primeiras fases do desenvolvimento das gónadas femininas (Almeida, 2013). Como resposta a este estímulo, o ovário produz 17β-estradiol, cuja função é por exemplo,a libertação de VTG pelo fígado e a sua entrada nos oócitos. A GtH I também é responsável pela produção e armazenamento da GtH II na glândula pituitária(Connaughton e Aida, 1999 cit.por Sanchez, 2006).
Nos machos, a GtH II que chega às gónadas masculinas (testículos), aumenta a atividade da enzima 20β-HDS (20β-hidroxiesteroide dehidrogenase), que atua na conversão do colesterol em 17αOH-P (17-α-hidroxiprogesterona). Nas fêmeas, a GtH II estimula a produção da substância indutora da maturação de oócitos, a 17α20β-P, nas células dos folículos ováricos (Almeida, 2013). Existem outras hormonas, além das libertadas nas gónadas, que podem participar na reprodução, tais como as hormonas da tiróide, a Triiodotironina-T3 e a Tiroxina-T4. Estas hormonas, principalmente a T3, favorece a ação das GtH I e II no início do desenvolvimento das gónadas femininas. Com o aumento de volume dos ovários, há o aumento de secreção do 17α-estradiol, o qual
12
inibe a libertação das hormonas da tiróide. Desta forma, a energia produzida pelo peixe será destinada para o crescimento das gónadas e não para o crescimento corporal. Contudo, em algumas espécies, as hormonas da tiróide aumentam novamente no final da vitelogénese, maturação das gónadas e no processo de desova (Baldisserotto, 2002).
Figura 1. Esquema da indução de vitelogenina controlada pelo sistema endócrino em peixes fêmea
(adaptado de: http://mn.water.usgs.gov/projects/CED/summary.html).
2.4.Macrolesões ou “Gross lesions”
Aquando da captura dos peixes pode ser feita uma primeira avaliação da saúde do peixe, baseada na observação de lesões macroscópicas visíveis a olho nu (macrolesões ou “gross lesions” externas). Durante a necropsia também pode ser feita uma observação de possíveis macrolesões internas. Estas lesões ou alterações aparecem quando os mecanismos de compensação e defesa orgânica foram ultrapassados (Carrola, 2003).
Não se deve confundir as macrolesões com outras alterações “lesões confundíveis” (confunding lesions), pois estas são provocadas por vírus, parasitas, fungos, bactérias ou outro tipo de fator (Black, 2002). Quando, por exemplo, se detetam neoplasias da derme, certamente que já terá havido impacto na população de peixes. As macrolesões e as lesões histopatológicas servem como indicadores primários do tipo de efeitos tóxicos resultantes da exposição aos tóxicos (Au, 2004).
As macrolesões incluem: erosão das barbatanas, pigmentação anormal (peixes com manchas ou com ausência de cor), úlceras na pele, lesões nas brânquias, alterações e
Monitorização e Biomonitorização Ambiental
13
lesões nos olhos, tumores visíveis e deformação do esqueleto (Au, 2004), e estão normalmente relacionadas com sedimentos intensamente contaminados (Carrola, 2003). No entanto, é imprescindível comparar o grau de anomalias obtido com o valor do local de referência, pois podem também ser provocadas por deficiências nutricionais (Carrola, 2003). A epiderme é alvo de úlceras e vários tipos de neoplasias, particularmente, em peixes bentónicos selvagens, pois está em contato direto e continuo com os sedimentos contaminados com níveis elevados de carcinogéneos hidrofóbicos.
A maioria das macrolesões necessita de interação entre vários fatores: sensibilidade dos peixes, stresse, doenças infeciosas, alimentação, entre outros, para se desenvolverem (Schmitt e Dethloff, 2000).
2.5. Compostos desreguladores endócrinos (CDE)
Um composto desregulador endócrino (CDE) é uma substância ou um composto exógeno que altera uma ou várias funções do sistema endócrino (reprodução, desenvolvimento, sistema imunitário, etc.) e tem, consequentemente, efeitos adversos sobre a saúde dum organismo, ou na sua descendência, ou mais tarde nas (sub) populações, sendo definido não pela sua natureza química, mas de acordo com os seus efeitos biológicos (Węsierska-Gądek, 2006). Os CDE podem ser divididos em: compostos naturais, tais como, fitoestrogénios ou micotoxinas, entre outros ou ainda produtos químicos sintéticos, como por exemplo, o bisfenol A (BPA), tributilestanho (TBT), ftalatos, pesticidas, entre muitos outros (Hampl et al., 2015).
Os CDE podem provocar efeitos negativos mesmo em concentrações muito baixas (Silva et al., 2002) causando danos no sistema imunológico, crescimento, comportamento, reprodução (supressão das características sexuais secundárias) (Puy-Azurmendi et al., 2013) e, em casos extremos, numa diminuição das populações de peixes (Harris et al., 2011).
Estes compostos podem atuar de diversas maneiras, nomeadamente: (i) imitando a ação das hormonas naturais: (ii) bloqueando a sua produção; (iii) inibindo ou estimulando o sistema endócrino (Arukwe e Goksoyr, 2003); ou (iv) alterando a quantidade dos recetores hormonais (Figura 2) (Markey et al., 2002).
14
Figura 2. Esquema com alguns mecanismos de disfunção endócrina (adaptado de Ghiselli e Jardim, 2007).
A) As hormonas naturais encaixam-se perfeitamente nos recetores e transmitem os sinais indispensáveis às
células. B) Os compostos desreguladores endócrinos ocupam o lugar das hormonas naturais encaixando-se perfeitamente nos recetores, enviando sinais diferentes e fora de tempo às células e C) E por fim, os desreguladores endócrinos atuam como bloqueadores dos sinais normais das hormonas naturais que seriam enviados às células.
Dependendo da sua persistência no meio ambiente, os CDE são geralmente divididos em compostos de vida curta ou em compostos persistentes (Hampl et al., 2015).
Os compostos químicos com atividade estrogénica incluem: (a) pesticidas - diclorodifeniltricloroetano (DDT); (b) anti-oxidantes – alquilfenóis (AP); (c) plastificantes e monómeros - ftalatos, BPA; (d) desinfetantes - orto-fenilfenol; (e) outros produtos químicos, tais como, BPC e protetores solares (Markey et al., 2002). As três classes de compostos desreguladores endócrinos que têm maior impacto no ecossistema e que por isso suscitam maior interesse científico e público são: bisfenóis, alquilfenóis e ftalatos (Gultekin e Ince, 2007).
Entre os CDE, os xenoestrogénios, como AP, tem a capacidade de “imitar” os estrogénios naturais. Alguns dos efeitos provocados por xenoestrogénios em peixes, tanto em estudos de laboratório como no campo, incluem: desenvolvimento anormal dos gâmetas, alterações nas concentrações de esteróides sexuais (Kortner et al., 2009) e indução da VTG em peixes macho (Gimeno et al., 1996). Causam também perturbações na fase larvar ou na fase de desenvolvimento dos peixes, causando anomalias no desenvolvimento sexual, comportamento e fertilidade (Jobling et al., 2002).
Monitorização e Biomonitorização Ambiental
15
Em condições normais, apenas peixes fêmea maduras produzem vitelogenina (lipo-fosfo-glicoproteína ligada ao cálcio) através dos hepatócitos, como resposta aos estrogénios (Puy-Azurmendi et al., 2013), transportada pelo sangue até ao ovário, onde é retida para formar as proteínas do oócito que irão servir como reserva de nutrientes para o desenvolvimento das larvas (Islinger et al., 2002). Embora o gene da vitelogenina seja normalmente silencioso em peixes macho e juvenis, este pode ser expresso após tratamento com estrogénios naturais ou sintéticos (Sumpter e Jobling, 1995). Assim, a indução de vitelogenina em peixes macho ou a variação nos seus níveis de expressão são uma evidência direta da exposição a CDE (Bjorkblom etal., 2008) e torna-se por isso um biomarcador de exposição muito útil.
A interação entre xenoestrogénios e recetores de estrogénio (RE) é um mecanismo importante de desregulação endócrina em peixes (Arukwe e Goksoyr, 2003). Os recetores de estrogénio (RE) regulam a síntese de VTG através da ligação com os estrogénios. Estes tem capacidade para se ligar a diversos análogos de estrogénio e a diversos compostos não-esteróides (Crews et al., 2000). Assim, os xenoestrogénios podem desregular a síntese de VTG através da ligação direta e ativação dos RE (Puy-Azurmendi et al., 2013).
2.5.1. Efeitos provocados nos peixes
As fases iniciais de desenvolvimento dos peixes são bastante sensíveis aos xenobióticos (Selderslaghs et al., 2009). A exposição dos embriões aos CDE pode afetar a expressão dos genes, induzir a apoptose e diminuir a taxa de eclosão e sobrevivência das larvas (Santos et al., 2014). Várias experiências realizadas em laboratório indicam que os CDE tem um impacto negativo na saúde dos peixes, como por exemplo, no crescimento e no sistema endócrino (Mills e Chichester, 2005). Estes podem alterar também a expressão de genes relacionados com estrogénio, tais como cyp19a e cyp19b, resultando em anomalias no desenvolvimento e reprodução em peixes (Kortner et al., 2009).
A exposição aos desreguladores endócrinos pode ser detetada através de mudanças nas características sexuais de uma espécie de peixe (Rautenberg et al., 2015). Foram estabelecidos dois endpoints como indicadores de desregulação endócrina: a indução de VTG e intersexualidade – condição de ovotestis - ou seja, alteração no tecido histológico
16
das gónadas masculinas de peixes com aparecimento de oócitos em diferentes estados de desenvolvimento (Reinen et al., 2010; Bjerregaard et al., 2006; Jobling et al., 1998).
A intersexualidade influencia a reprodução, distribuição e abundância de peixes nativos (Tetreault et al., 2011; Bateman et al., 2004). Vários estudos demonstraram que esta condição está relacionada com a presença dos CDE (Bahamonde et al., 2013) em várias espécies de peixes, tais como, em tainhas (Chelon labrosus, Risso, 1826) como demonstrado no estudo realizado na Reserva da Biosfera do Urdaibai na Costa Basca (Puy-Azurmendi et al., 2013).
Outros efeitos provocados pelos CDEs nos peixes são: impedimento o desenvolvimento das gónadas (van Dyk et al., 2012; Jobling et al., 1998), alterações nas gónadas femininas (ausência de células germinativas) (Soyano et al., 2010), aumento da atresia do ovário (em peixes fêmea), redução do sucesso da desova, inibição do desenvolvimento de características sexuais secundárias (Lange et al., 2001), redução do sucesso de eclosão e/ou sobrevivência das larvas (Jobling e Tyler, 2003a), diminuição da fertilidade (Jobling et al., 1998) e alterações no fígado (van Dyk et al., 2012).
17
3. GÓNADA MASCULINA - TESTÍCULO
18 GÓNADA MASCULINA - TESTÍCULO
A histologia representa uma ferramenta útil na avaliação do estado de saúde dos peixes refletindo o grau de poluição (van der Oost et al., 2003).
3.1. Histologia da gónada masculina
A maior parte dos peixes ósseos apresentam reprodução sexuada, e com fertilização externa, a qual ocorre no meio aquático. A morfologia das células germinativas e os elementos somáticos constituintes do tecido da gónada são semelhantes em todas as espécies. O desenvolvimento das células germinativas femininas e masculinas denomina-se gametogénedenomina-se e decorre de processos meióticos, que denomina-se designam, respetivamente, por oogénese e espermatogénese (Quagio-Grassiotto, 2013). Os machos possuem em geral dois testículos alongados, que são maiores na zona central e mais finos na região posterior, com maior espessura na região dorsal do que na ventral quando o peixe está maduro (Leino et al., 2005), ao invés dos peixes achatados como linguado ou solha, que possuem testículos com a forma de meia esfera.
Os testículos estão envolvidos por uma membrana muito fina (mesotélio) que cobre o estroma – tecido conjuntivo onde se encontram os tubos seminíferos e as células intersticiais. Todos os tubos seminíferos reúnem-se num ducto, chamado ducto espermático, que transporta o esperma para o exterior (Costa, 2004). Durante a espermatogénese, as células germinativas primordiais diferenciam-se, originando as espermatogónias, as quais sofrem transformações que vão dar origem às células reprodutoras masculinas - os espermatozóides (Costa, 2004). Além das células germinativas, outros tipos de células estão presentes nos testículos, nas quais se destacam as células de Sertoli, e as células intersticiais ou de Leydig (Quagio-Grassiotto, 2013).
As espermatogónias mantêm-se junto à membrana basal dos tubos seminíferos e sofrem dois tipos de divisão: mitótica, na qual origina os espermatócitos primários, caracterizados pelo aspeto dos cromossomas em novelo; e meiótica, leva à formação dos espermatócitos secundários, células haplóides com a cromatina nuclear em forma de grânulos (Jensen et al., 2001). A partir dos espermatócitos secundários originam-se os
Gónada masculina - testículo
19
dois estados finais da diferenciação das células sexuais masculinas: os espermatídios e finalmente os espermatozoides, tal como se pode ver na Figura 3 (Costa, 2004).
A formação dos espermatozoides engloba a formação de uma cauda ou flagelo, modificação da célula arredondada numa célula alongada e a redução do volume do citoplasma, tudo para dar maior mobilidade à nova célula. A presença ou ausência destas fases num corte histológico pode servir para avaliar o estado de maturação testicular (Leino et al., 2005).
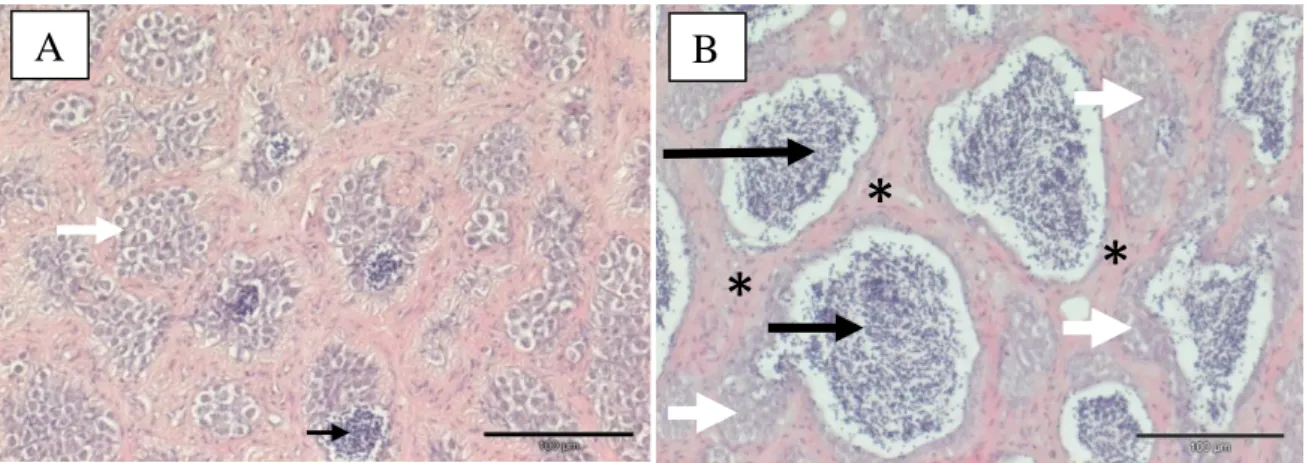
Figura 3.Microfotografias do tecido das gónadas masculinas das solhas capturadas no estuário do Douro em 2013. Gónada masculina imatura (A) e gónada masculina madura (B) (H&E). A) Presença de espermatozóides em fase de desenvolvimento (seta preta) e aglomerados de espermatócitos (seta branca), barra = 100 µm; B) Microfotografia do tecido testicular de solha, onde se podem observar cortes de tubos seminíferos com espermatozóides (seta preta), tecido conjuntivo intersticial (asteriscos), bem como aglomerados de espermatócitos (seta branca), barra = 100 µm.
3.2. Condição de intersexo como biomarcador
Muitas espécies de teleósteos são gonocóricas - indivíduos que se desenvolvem em machos ou fêmeas e assim permanecem todo o seu tempo de vida (Devlin e Nagahama, 2002). Outras espécies são hermafroditas, produzindo tanto gâmetas femininos como masculinos alguma vez durante o tempo de vida (Bahamonde et al., 2013). Isto inclui: estratégias síncronas hermafroditas, em que os indivíduos desenvolvem gâmetas masculinos e femininos, ao mesmo tempo, ou hermafroditas sequenciais; classificadas como protandria (primeiro desenvolve-se como macho) ou protoginia (primeiro desenvolve-se como fêmea) (Devlin e Nagahama, 2002). A condição de intersexo é uma anomalia definida como a presença simultânea de tecido testicular e ovárico no mesmo
A
B
*
*
20
indivíduo de uma espécie gonocórica (sexos separados) (Tyler e Jobling, 2008; Lyons et
al., 2004). No entanto, esta condição é também referida como a presença de oócitos testiculares, folículos testiculares, testis-ova (Hirakawa et al., 2012; Ferreira et al., 2004), ou ovotestis (Carrola, 2011), às vezes também aparece o termo ova-testis (Bahamonde et
al., 2013). Por questões de simplificação, ao longo deste trabalho utilizarei o termo ovotestis.
Nem todas as espécies de peixes gonocóricas desenvolvem a condição de intersexo (mesmo em locais muito poluídos com CDE) enquanto que outras podem apresentar intersexo, isto indica que pode haver espécies mais sensíveis à poluição do que outras. No entanto, em algumas espécies de peixes, o intersexo pode ocorrer espontaneamente devido ao stresse ou à presença de parasitas (Jobling e Tyler, 2003b). A condição de intersexo pode variar de acordo com a exposição aos CDE (Bahamonde et al., 2013) podendo tornar-se num processo de feminização (presença de oócitos no tecido testicular) (Vasconcelos et al., 2006) ou de masculinização (presença de tecido testicular nos ovários) (Ghiselli e Jardim, 2007). O tipo mais frequente de intersexo é ovotestis, onde os gâmetas femininos estão distribuídos pelo tecido testicular (Getsfrid et al., 2004).
Há fatores que devem ser considerados quando se avalia a prevalência de ovotestis, tais como, a estação de amostragem, nível de maturidade e a idade do individuo (peixe) (Bahamonde et al., 2013).
21
4. TECIDO HEPÁTICO E
22 TECIDO HEPÁTICO E HISTOPATOLOGIA
A observação e análise das alterações histológicas no fígado (Carrola, 2011) ou na gónada (Miranda, 2006) de peixes constitui um meio importante para avaliar o impacto causado pela crescente poluição dos ecossistemas aquáticos. O fígado desempenha um papel importante no metabolismo e é o principal órgão de acumulação, biotransformação, e excreção de contaminantes, como por exemplo DDT nos peixes (Souza et al., 2013;
Figueiredo-Fernandes et al., 2006). As principais funções do fígado incluem ainda, a síntese e secreção de proteínas e bílis e formação de ureia a partir de iões de amónia (Ross
et al., 1989).
De acordo com Ross et al. (1989), o fígado possui funções endócrinas pois a maioria dos seus produtos são libertados diretamente na corrente sanguínea e exócrinas porque um dos seus produtos (bílis) é secretada através de um sistema de ductos até ao duodeno. As células hepáticas apresentam diversas funções essenciais para a sobrevivência do organismo, tais como, metabolismo de proteínas, lípidos e hidratos de carbono, servem, também, de local de armazenamento para muitos nutrientes como a glucose (Balula, 2011).
O fígado é um órgão importante no estudo de biomarcadores histopatológicos devido às suas funções (anteriormente referidas) e porque os compostos persistentes acumulam-se em maior extensão no acumulam-seu interior do que em outros tecidos (Stentiford et al., 2003). Por ser um órgão fundamental na biotransformação de xenobióticos, expõe as suas células a diversos agentes químicos, o que pode em função do tempo de exposição e concentração dos tóxicos provocar diversas lesões neste órgão (Pacheco e Santos, 2002).
A biotransformação pode ser definida como uma conversão catalisada por enzimas de um composto numa forma mais solúvel em água, de forma a ser excretado mais facilmente do que o composto original (Lech e Vodicnik, 1985). Essas alterações podem refletir-se numa variedade de processos patológicos tais como, lesões hepáticas externas e internas, incluindo neoplasmas, ou até levar à morte do animal (Ben Ameur et al., 2012). A toxicidade de um composto pode ser afetada pelo seu metabolismo podendo ser benéfico (desintoxicação), ou prejudicial (bioativação dos compostos) no organismo (van der Oost et al., 2003).
Tecido hepático e histopatologia
23 4.1. Histologia do fígado de peixe
A fim de avaliar a histologia de um órgão de uma dada espécie exposta a contaminantes, é importante analisar e conhecer bem a histologia do mesmo órgão em indivíduos (macho e fêmea) saudáveis dessa espécie, assumindo ser a estrutura histológica normal desse órgão. Isto permitirá identificar facilmente alterações que possam estar presentes no tecido. De acordo com Hinton eLaurén (1990) o fígado compreende dois compartimentos: parênquima (as células epiteliais desempenham funções importantes do órgão) e estroma (vasos sanguíneos e tecido conjuntivo). O parênquima inclui células (hepatócitos), bem como os respetivos espaços extracelulares (Figura 4). O fígado dos peixes não mostra qualquer organização lobular clara, mas apresenta uma estrutura tubular, que varia entre diferentes espécies (Rocha et al., 2010). O fígado da maioria dos peixes teleósteos é composto por dois lobos, o lobo direito que está próximo da vesícula biliar, e o lobo esquerdo próximo ao baço (Silva, 2004). As células que constituem o fígado são: hepatócitos, células epiteliais dos ductos biliares, macrófagos, células sanguíneas e as células endoteliais (Takashima e Hibiya, 1995). As principais células do fígado, os hepatócitos, são células poliédricas com um ou dois núcleos grandes, esféricos e centralmente localizados, com nucléolo evidente, e o citoplasma com granulações e aspeto vacuolizado (Silva, 2004), e tem como funções, por exemplo, armazenar quantidades consideráveis de glicogênio e processar lípidos.
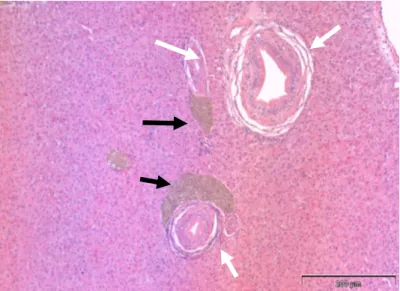
Figura 4.Microfotografia do fígado (corte transversal) de tainha capturada no estuário do Tejo (H&E) onde estão representados ductos biliares (setas brancas) sendo que dois deles estão junto a agregados macrofágicos (setas pretas), barra = 200 μm.
24 4.2. Histopatologia hepática
A desintoxicação de resíduos endógenos e xenobióticos nos peixes, como em todos os vertebrados, é realizada principalmente no fígado, causando consequentemente, alterações patológicas neste órgão (Grinwis et al., 2000). A histopatologia é o estudo microscópico de tecidos que apresentam lesões. As lesões são, por isso consideradas como biomarcadores histopatológicos de exposição aos xenobióticos e como resultado, a histopatologia do fígado dos peixes tem sido cada vez mais reconhecida como uma ferramenta valiosa para a deteção de efeitos crónicos (Stehr et al., 1998).
Várias lesões hepáticas, incluindo neoplasias, pré-neoplasias, proliferação não-neoplásica, e lesões necróticas/degenerativas foram associadas com a exposição a xenobióticos (Myers et al., 1990).O contraste de cor existente entre um nódulo e tecido hepático é importante para a sua deteção, visto que são de tamanho reduzido e sobressaem pouco do tecido normal adjacente, apresentando também uma textura distinta que os torna mais facilmente detetáveis pelo tato (Carrola, 2003). No entanto a diminuição das lesões hepáticas deve-se à redução da exposição de contaminantes, tais como HAP, porém, na maioria dos sistemas aquáticos é difícil estabelecer uma relação causa-efeito ou uma correlação entre os xenobióticos e a saúde dos peixes devido à grande quantidade de fatores (e.g. idade, espécie, época de migração) que podem influenciar ou modificar as respostas dos organismos sujeitos a um agente xenobiótico particular (Ham et al., 1997). Os biomarcadores histopatológicos dos peixes oferecem vantagens tais como, facilidade de colheita de amostras, capacidade de avaliar vários sistemas do corpo e tipos de células do mesmo individuo (Ben Ameur et al., 2012).
As respostas histopatológicas à exposição a xenobióticos podem ser avaliadas de forma mais segura em peixes jovens em que os efeitos devido à migração entre habitats contaminados e não contaminados (por exemplo, migrações de desova) ainda não ocorreram (Myers et al., 1998). Além disso, as neoplasias hepáticas raramente são detetadas em peixes juvenis selvagens (Myers et al., 1998). Quando expostos aos contaminantes os peixes podem desenvolver diferentes lesões e alterações hepáticas que podem ser agrupadas em vários tipos e classificadas de acordo com a sua severidade (Carrola, 2011) segundo a classificação de Lang et al. (2006) (Tabela 1).
Tecido hepático e histopatologia
25
Tabela 1. Principais alterações hepáticas em pleuronectiformes e sistema de classificação usado para a sua
quantificação na solha-das-pedras (Lang et al., 2006)
Categoria da lesão Tipo Grau Ponto
0 Nenhuma anomalia detectada
- - 0
1 Lesão não-específica Alteração inflamatória (infiltração/granulomatose) Alteração degenerativa (necrose celular
coagulativa ou simples, atrofia)
Alteração proliferativa (regeneração hepatocelular, fibrose, hiperplasia dos ductos biliares)
Moderado 1
Aumento do número/área de agregados macrofágicos
Médio 2
Lipidose; Parasitas Severo 3
2 Lesões não neoplásicas toxicopáticas precoces
Pleomorfismo hepatocelular/ nuclear
Vacuolização hidrópica (hepática/ biliar/ pancreática)
Fosfolipidose dos hepatócitos Moderado 4
Inclusões fibrilares Médio 5
Peliose e espongiose hepática Severo 6
3 Lesões pré-neoplásicas Focos de alteração celular:
Focos de células claras; Vacuolizadas; Eosinofílicas
Moderado 7
Focos Basofílicos; Mistos Médio 8
Severo 9
4 Neoplasias benignas Adenoma hepatocelular
Colangioma Moderado 10
Hemangioma Médio 11
Adenoma dos ácinos pancreáticos e outros Severo 12 5 Neoplasias malignas Carcinoma hepatocelular
Colangiocarcinoma Moderado 13
Carcinoma dos ácinos pancreáticos Médio 14
Hemangiocarcinoma Severo 15
Outros
Os macrófagos no fígado de peixes estão relacionados com a fagocitose e defesa do organismo. Um aumento do tamanho e número de agregados macrofágicos (AM) ou melanomacrofágicos, local de acumulação de células fagocitárias saturadas de dejetos hepatocelulares, foi observado em peixes capturados em locais contaminados (Triebskorn
26
et al., 2002), no entanto a sua utilização como biomarcador não especifico deva ter em atenção alguns fatores, tais como, idade, estado nutricional e etiologias multifatoriais.
A vacuolização hidrópica pode ser das células epiteliais biliares ou hepatocelulares, caracterizada pelo aumento hidrópico severo do espaço citoplasmático. A vacuolização hidrópica dos hepatócitos pode ser difícil de diferenciar da acumulação de lípidos no citoplasma, contudo, as células afetadas contêm poucos vacúolos, embora de grandes dimensões e pouco citoplasma. As necroses hepáticas dos peixes (difusas ou extensas) é uma das expressões morfológicas da morte celular e podem ser provocadas experimentalmente pela exposição prolongada a produtos hepatotóxicos e/ou por concentrações elevadas de xenobióticos (Carrola, 2003). A picnose carateriza-se pela redução de volume do núcleo da célula e identificando-se por estar híper corado. Os granulomas são pequenos nódulos de caráter inflamatório formados por macrófagos mas que também podem conter outros leucócitos, e servem para isolar bactérias, fungos ou substâncias estranhas insolúveis que o organismo foi incapaz de expulsar.
Os linfócitos são um tipo de célula de defesa do organismo, que são produzidos em maior quantidade quando existe uma infeção. A inflamação é um processo tecidular constituído por uma série de processos moleculares, celulares e vasculares de finalidade defensiva frente a agressões físicas, químicas ou biológicas. A histopatologia gera dados sobre as lesões não especificando a causa pontual da lesão, mostrando apenas ser uma resposta biológica ao fator de stresse. No entanto, quando associados a outros métodos de análises, a análise histológica pode auxiliar na compreensão de determinadas situações.
27
5. OBJETIVOS
28 OBJETIVOS
O principal objetivo deste trabalho consiste na análise histopatológica do fígado e no estudo da gónada masculina – testículo - em mugilideos (fam. Mugilidae) capturados no estuário do Tejo de maneira a avaliar possíveis alterações associadas à poluição, e inferir sobre o estado ecológico e a poluição do estuário.
O trabalho envolve a identificação das principais lesões hepáticas e a determinação da respetiva prevalência e gradação, com o intuito de permitir avaliar a integridade do fígado e consequências para saúde do peixe para poder inferir sobre a qualidade da água e do seu nível de compostos hepatotóxicos.
Pretende ainda avaliar o efeito de potenciais compostos desreguladores endócrinos, usando a histopatologia da gónada, nomeadamente do testículo. Para isso, envolve a pesquisa da condição de ovotestis, avaliação da sua prevalência, bem como a avaliação da sua severidade (ISO – Índice de severidade de ovotestis) segundo a metodologia de Bateman et al. (2004).
A informação será depois analisada de forma conjunta e comparada com os resultados observados em tainhas de outros estuários.
29
6. METODOLOGIA
30 METODOLOGIA
A realização deste trabalho envolveu a captura de 100 tainhas, tainha fataça (1 Liza ramada, Risso, 1826), 59 tainha liça (Chelon labrosus, Risso, 1827) e 40 tainha-garrento (Liza aurata, Risso, 1810) no estuário do Tejo em fevereiro e novembro de 2014 (Tabela 2).
Tabela 2. Número de tainhas capturadas (n) em fevereiro e novembro de 2014.
Fevereiro de 2014 Novembro de 2014 L. aurata 22 18 C. labrosus 37 22 L. ramada 0 1 Total 59 41 n= 100 6.1. Área de estudo
O rio Tejo na Serra de Albarracín (Espanha), e desagua no oceano Atlântico através de um estuário - estuário do Tejo -, onde existe uma reserva natural que abrange cerca de 14.560 há. O rio Tejo é a principal fonte de água doce deste estuário (Figura 5).
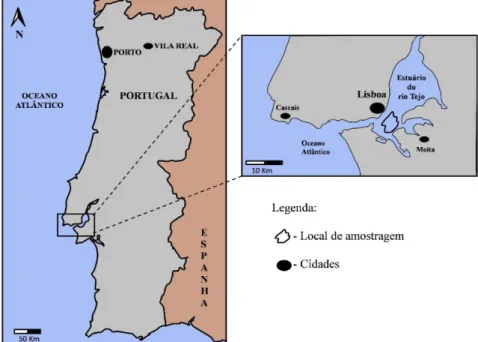
Figura 5.Representação geográfica do estuário do Tejo e do local amostragem (realizado por Sofia Ribeiro segundo a metodologia de Carrola (2011).
Metodologia
31
O estuário do Tejo tem uma área com cerca de 320 km2 (maré baixa) e que se extende por cerca de 80 km (Almeida et al., 1993). É constituído por um canal de água profundo e por uma baía larga de baixa profundidade. As zonas Sul e Este da baía são constituídas por extensos sapais intertidais, com um predomínio típico de vegetação halófita.
6.1.1. Caracterização da L. ramada, C. labrosus e L. aurata
O termo comum tainha integra várias espécies, tais como, L. ramada, C. labrosus, L. aurata (Figura 6, Figura 7 e Figura 8) e habitam diferentes estuários portugueses (Carrola, 2011). As espécies são muito semelhantes nos aspetos fisiológicos e histológicos, apresentando diferenças ligeiras na morfologia o que, em termos práticos, pode dificultar a sua identificação (Carrola, 2011) encontram-se por todo o mundo podendo ser facilmente encontradas em estuários situados na Europa incluindo, os estuários portugueses (Carrola, 2011).
Figura 6. Fotografia da tainha fataça (L. ramada) (disponível em:
http://fineartamerica.com/featured/thinlip-mullet-liza-ramada-mulet-morragute-cefalo-tainhatunnlaeppad-multe-roendungur-urft-valley-art.html).
Figura 7. Ilustração da tainha liça (C. labrosus) (disponivel em:
32
Figura 8. Ilustração da tainha garrento (L. aurata) (disponível em:
http://w3.ualg.pt/~madias/guadiana/garrento.html).
Classificação taxonómica
A taxonomia da tainha pode ser resumida como se segue:
Reino: Animalia Filo: Chordata
Classe: Actinopterygii Ordem: Mugiliformes Família: Mugilidae
6.1.2. Morfologia da L. ramada, C. labrosus e L. aurata
As características morfológicas destas três espécies são semelhantes, pois são peixes de médias dimensões, apresentam duas barbatanas dorsais bem separadas, tendo primeira barbatana dorsal 4 raios. O lábio superior é estreito e o olho não apresenta membrana ocular adiposa (Guerra, 1976). À exceção da L. ramada que possui cabeça mais pontiaguda e achatada e o contorno da cabeça forma um ângulo obtuso. A média relativa ao peso total e comprimento total das tainhas adultas ronda 1 kilo e 50 cm.
6.1.3. Sistema reprodutor da L. ramada, C. labrosus e L. aurata
A L. ramada pode ser considerada peixe catádromo (McDowall, 1998) pois, quando adultas, efetuam migrações dos rios ou estuários em direção ao mar para aí se reproduzirem. Esta migração, muitas vezes em grandes cardumes, estende-se pelos meses