Coordenação editorial: José Morais Arnaud, Andrea Martins Design gráfico: Flatland Design
Produção: Greca – Artes Gráficas, Lda. Tiragem: 500 exemplares
Depósito Legal: 433460/17 ISBN: 978-972-9451-71-3
Associação dos Arqueólogos Portugueses Lisboa, 2017
O conteúdo dos artigos é da inteira responsabilidade dos autores. Sendo assim a As sociação dos Arqueólogos Portugueses declina qualquer responsabilidade por eventuais equívocos ou questões de ordem ética e legal.
Desenho de capa:
Levantamento topográfico de Vila Nova de São Pedro (J. M. Arnaud e J. L. Gonçalves, 1990). O desenho foi retirado do artigo 48 (p. 591).
o crânio humano acheulense
do plistocénico médio da gruta
da aroeira
Joan Daura1, Montserrat Sanz2, Juan Luis Arsuaga3, Rolf Quam4, Dirk L. Hoffmann5, Maria Cruz Ortega6, Elena Santos7, Sandra Gómez8, Ángel Rubio9, Lucia Villaescusa10, Pedro Souto11, Filipa Rodrigues12, João Mauricio13, Artur Ferreira14, Paulo Godinho15, Erik Trinkaus16, João Zilhão17
1. Grup de Recerca del Quaternari (GRQ) del Seminari Estudis i Recerques Prehistòriques (SERP), Dept. d’Història i Arqueologia, Universitat de Barcelona; UNIARQ – Centro de Arqueologia da Universidade de Lisboa; jdauralujan@campus.ul.pt
2. UNIARQ ‑Centro de Arqueologia da Universidade de Lisboa / Centro UCM ‑ISCIII de Investigación sobre la Evolución y Com‑ portamiento Humanos, Universidad Complutense de Madrid / Departamento de Paleontología. Ciencias Geológicas. Universidad Complutense de Madrid; msanzborras@ucm.es
3. Centro UCM ‑ISCIII de Investigación sobre la Evolución y Comportamiento Humanos, Universidad Complutense de Madrid / Departamento de Paleontología. Ciencias Geológicas. Universidad Complutense de Madrid; jlarsuaga@isciii.es
4. Department of Anthropology, Binghamton University; rquam@binghamton.edu
5. Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology; dirk_hoffmann@eva.mpg.de
6. Centro UCM ‑ISCIII de Investigación sobre la Evolución y Comportamiento Humanos, Universidad Complutense de Madrid; mcortega@isciii.es
7. Centro UCM ‑ISCIII de Investigación sobre la Evolución y Comportamiento Humanos, Universidad Complutense de Madrid; esureta@ubu.es
8. Grup de Recerca del Quaternari (GRQ) del Seminari Estudis i Recerques Prehistòriques (SERP), Dept. d’Història i Arqueologia. Universitat de Barcelona; gomezsolers@gmail.com
9. Laboratorio de Antropología, Depto de Medicina Legal, Toxicología y Antropología Física, Universidad de Granada; a_rubiosal‑ vador@hotmail.com
10. Grup de Recerca del Quaternari (GRQ) del Seminari Estudis i Recerques Prehistòriques (SERP), Dept. d’Història i Arqueologia. Universitat de Barcelona; lucia.villaescusa.fernandez@gmail.com
11. Crivarque – Estudos de Impacto e Trabalhos Geo ‑Arqueológicos Lda e Sociedade Torrejana de Espeleologia e Arqueologia; psouto@crivarque.net
12. Crivarque – Estudos de Impacto e Trabalhos Geo ‑Arqueológicos Lda e Sociedade Torrejana de Espeleologia e Arqueologia; UNIARQ ‑FLUL; frodrigues@crivarque.net
13. Crivarque – Estudos de Impacto e Trabalhos Geo ‑Arqueológicos Lda e Sociedade Torrejana de Espeleologia e Arqueologia; jm@ crivarque.net
14. Crivarque – Estudos de Impacto e Trabalhos Geo ‑Arqueológicos Lda; geral@crivarque.net 15. Crivarque – Estudos de Impacto e Trabalhos Geo ‑Arqueológicos Lda; geral@crivarque.net 16. Department of Anthropology, Washington University, trinkaus@wustl.edu
17. UNIARQ ‑Centro de Arqueologia da Universidade de Lisboa; ICREA (Catalan Institution for Research and Advanced Studies); Seminari Estudis i Recerques Prehistoriques (SERP), Dept. d’Historia i Arqueologia, Universitat de Barcelona; joao.zilhao@ub.edu
Resumo
A descoberta de um crânio humano com cerca de 400.000 anos de antiguidade na Gruta da Aroeira vem reforçar o conhecimento da diversidade humana e do povoamento acheulense da Europa. O crânio foi encontrado na Unidade 2, associado a uma importante colecção de bifaces e a uma fauna de grandes mamíferos.
O fóssil é uma porção considerável do lado direito da caixa craniana, sem o occipital, mas conservando parte do lado esquerdo da escama frontal e do toro supraorbital, bem como a região interorbital, incluindo a parte vertical dos ossos nasais. Algumas características do crânio da Aroeira são traços primitivos presentes em fósseis do Plistocénico médio mas ausentes nos Neandertais.
Palavras ‑chave: Gruta da Aroeira, Acheulense, Restos humanos, Plistocénico médio.
AbstRAct
the middle Pleistocene human cranium from the Acheulean of Gruta da Aroeira
The discovery of a c.400,000 year ‑old cranium at Gruta da Aroeira sheds new light on the diversity of past humans and the Acheulean settlement of Europe. The fossil is a partial cranium encased in the hard breccia forming Unit 2 of the stratigraphic sequence. It was found in association with a rich collection of bifaces and large mammal remains. It preserves most of the right half of the calvarium (with the exception of the occipital bone), as well as a portion of the left side of the frontal squama and supraorbital torus. The combination of traits in the Aroeira 3 cranium increases the diversity previously documented in the Middle Pleistocene fossil record of Europe.
Keywords: Gruta da Aroeira, Acheulean, Human remains, Middle Pleistocene.
1. INtRoDuÇÃo
A evidência antropológica aponta para que as po‑ pulações europeias do Plistocénico médio fossem morfologicamente diversas, embora alguns autores agrupem num só clado os restos humanos de África e da Europa (Rightmire, 1998; Stringer, 2012). Não obstante, os sítios europeus com restos humanos diagnósticos associados a indústrias líticas acheu‑ lenses são escassos, limitando ‑se a Atapuerca – Sima de los Huesos (SH) (Falguères et al., 2004; Ar‑ suaga et al., 2014), Swanscombe (Stringer e Hublin, 1999) e Arago (de Lumley, 2015). Até à data, foram identificados dois grupos humanos no Plistocénico Médio da Europa: um, representado por Atapuerca (SH) e Swanscombe, faz parte do clado Neandertal (Stringer, 2012; Arsuaga et al., 2014; Meyer et al., 2016); o outro, representado pelos fósseis de Arago, tem sido atribuído quer a um estádio incipiente da evolução dos Neandertais (Dean et al., 1998), quer ao Homo heidelbergensis (Stringer, 2012), quer ainda a um Homo erectus tautavelensis (de Lumley, 2015). O crânio recém ‑descoberto na Gruta da Aroeira (Daura et al., 2017a, 2017b) apresenta um mosaico de características que sugerem um panorama mais complexo ainda. Seja como for, certo é que a tecno‑ logia acheulense aparece associada tanto aos fósseis de Arago e da Sima de los Huesos como ao fóssil da
Aroeira. Este facto indica que a introdução no con‑ tinente europeu das inovações do Acheulense não guarda relação directa com processos de migração, ou de expansão demográfica, de um ou outro dos grupos representados pelos fósseis conhecidos. 2. o sItIo DA GRutA DA ARoeIRA
A Gruta da Aroeira (39° 30’ 20’’ N; 08° 36’ 57’’ W) (Figura 1) localiza ‑se no complexo cársico do Al‑ monda (Torres Novas, Portugal) e foi inicialmente escavada entre 1998 e 2002 (Zilhão et al., 1993; Ma‑ rks et al., 2002). Ofereceu uma rica indústria lítica com bifaces acheulenses associada a restos faunísti‑ cos e a dois dentes humanos (Aroeira 1 e 2) (Trinkaus et al., 2003).
O sítio foi denominado “Galerias Pesadas” nas pu‑ blicações relativas a esses primeiros trabalhos de escavação (Marks et al., 2002; Trinkaus et al., 2003). Porém, esta designação refere ‑se a condutos inter‑ nos do sistema cársico situados à mesma cota que, presentemente, não têm nenhuma ligação ao exte‑ rior. Foram as prospecções espeleo ‑arqueológicas realizadas em 1991 que conduziram à identificação, nesses condutos, de um cone sedimentar com fau‑ na pleistocénica e materiais líticos (Zilhão et al., 1993), e à localização de uma entrada colapsada e se‑ lada por sedimentos – a Gruta da Aroeira – onde vi‑
riam a realizar ‑se tanto essas primeiras escavações (1998 ‑2002), como a nova fase do projecto, iniciada em 2013.
Estes novos trabalhos, centrados no estabelecimen‑ to de uma cronologia para a sequência através da da‑ tação pelo método do Urânio ‑Tório (U ‑Th) de cros‑ tas estalgmíticas interestratificadas (Hoffmann et al., 2013), levaram à descoberta, perto da base de um crânio humano (Aroeira 3) embalado numa brecha muito cimentada (Daura et al., 2017b).
A estratigrafia da Aroeira tem aproximadamente 4 m de potência e apresenta três grandes unidades estratigráficas. O crânio foi encontrado na Unida‑ de 2, constituída por uma brecha de 2.2 m de es‑ pessura com abundantes clastos angulares e sub‑ ‑arredondados. Esta unidade corresponde à camada acheulense X ‑Xb/c (respectivamente topo e base de um único horizonte estratigráfico, escavado em 1998 ‑2002 e 2013 ‑2015), e a crosta estalagmítica so‑ brejacente tem uma idade basal de 417.7+37.3/ ‑27.5 ka (milhares de anos; 2 sigma). Uma outra datação por U ‑Th de 406 ± 30 ka, obtida para a camada ex‑ terior de uma coluna estalagmítica coberta pela Unidade 2, estabelece a idade máxima da sequência e possibilita a sua correlação com o MIS 11. Duas ou‑ tras datações de 390 ± 14 ka e de 408 ± 18 ka, obti‑ das pelo mesmo método para crostas calcíticas for‑ madas sobre o crânio, fornecem balizas adicionais e coerentes para a idade mínima do mesmo. Portanto, o crânio Aroeira 3 data com uma elevada probabili‑ dade do intervalo entre 390 e 436 ka.
A nossa escavação da camada X ‑Xb/c incidiu sobre uma área de 6 m² e atingiu a rocha de base cerca de 1 m abaixo da cota alcançada em 2002. A indústria lítica (n=387) inclui bifaces e peças bifaciais acheu‑ lenses (n=17), outras peças retocadas (n=27), núcleos (n=43), lascas e fragmentos de lascas (n=180), e sei‑ xos (testados ou não, talvez manuportes) (n=114). A matéria ‑prima de eleição é o quartzito e o sílex é es‑ casso, mas existem bifaces talhados sobre ambos. O método Levallois não está representado na colecção. Os restos faunísticos estão muito fragmentados, consistindo sobretudo em dentes isolados, falan‑ ges, carpos/tarsos, e fragmentos de haste. Entre os 209 restos faunísticos coordenados na camada X ‑Xb/c predominam os cervídeos (NISP=58), in‑ cluindo Dama e Cervus, e os equídeos (NISP=46). Entre as espécies mais raras contam ‑se
Rhinocero-tidae (NISP=2) (provavelmente Stephanorhinus cf. hundsheimensis) e o urso (NISP=4) (Ursus sp.), bem
como um grande bovídeo (Bos/Bison), um caprídeo (Caprinae), e uma tartaruga (Testudo sp.) (NISP = 1 de cada). Foram encontrados 42 fragmentos de osso queimado na base da camada Xb/c, em associação com a indústria lítica e o crânio humano.
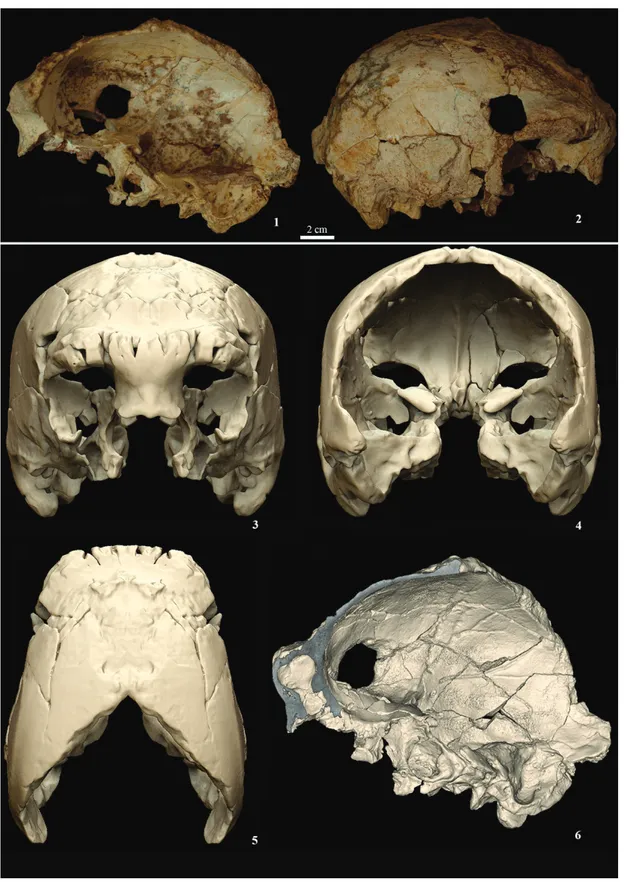
3. o cRÂNIo HumANo
O crânio Aroeira 3 foi laboriosamente extraído da dura brecha calcária e restaurado ao longo de um pe‑ ríodo de dois anos (Figura 2). A sutura coronal está completamente soldada, não se notando vestígios da mesma na superfície endocraniana. Os dentes que se conservam têm raízes completamente for‑ madas, com ápices fechados, e as coroas estão ali‑ sadas por desgaste. O conjunto destas observações indica que se trata de um indivíduo de idade adulta. Existem duas morfologias principais do toro su‑ praorbital entre os fósseis europeus do Plistocéni‑ co Médio. Em muitos casos, os arcos supraorbitais apresentam uma curvatura médio ‑lateral (em vista frontal) e a superfície anterior arredondada. Os dois arcos podem unir ‑se completamente, formando uma região glabelar inflada, ou podem manter ‑se mais ou menos separados por uma depressão gla‑ belar. Esta morfologia supraorbital, com diferentes graus de união na glabela, pode encontrar ‑se na Sima de los Huesos (SH) de Atapuerca e em crânios de Bilzingsleben (crânio B1), Steinheim e Petralona, e ainda nos Neandertais do Plistocénico Superior (Ar‑ suaga et al., 1997). Arago 21 e Ceprano são distintos, parecendo ‑se mais com fósseis africanos do Plisto‑ cénico Médio como os de Kabwe e Bodo, nos quais os dois arcos supraorbitais estão bem separados pela glabela e são mais planos e menos curvos (Manzi, Mallegni e Ascenzi, 2001; Rightmire, 1996). Ape‑ sar da perda da superfície externa de grande parte do toro supraorbital, parece ‑nos claro que, no caso de Aroeira 3, os arcos supraorbitais estão unidos na glabela e que o lado direito do toro, melhor conser‑ vado, é mais arredondado do que plano.
Os arcos supraorbitais direito e esquerdo são espes‑ sos, quando comparados com a maioria dos fósseis europeus ou africanos do Plistocénico Médio e Su‑ perior (Smith e Ranyard, 1980; Arsuaga et al., 1997). A espessura meso ‑orbital máxima do toro de Aroei‑ ra 3 (19.0 mm) pode ser medida no lado direito e é semelhante à dos crânios de Bodo e de Ceprano (17.5 mm em ambos). O toro do crânio B1 de Bilzingsle‑ ben é ainda mais espesso (21 ‑22 mm no lado direito;
medida tomada sobre molde). A distância entre os dois bordos orbitais internos é grande (34 ‑35 mm) e semelhante à observada nos crânios SH 4 de Ata‑ puerca (38.0 mm), de Kabwe (32.0 mm), de Bodo (37.5 mm), e B1 de Bilzingsleben (35.5 mm, sobre molde). Os seios frontais de Aroeira 3 estão bem desenvolvidos mas não são tão grandes lateralmen‑ te (no toro) ou superiormente (na escama frontal) como no fóssil de Petralona.
A reconstrução virtual do crânio Aroeira 3 (Figura 2) permite constatar que as paredes do parietal são quase verticais. Contudo, a largura máxima do crâ‑ nio localiza ‑se na crista supramastóide, tal como noutros fósseis do Plistocénico Médio europeu. Esta morfologia afasta ‑se tanto da condição ances‑ tral (acentuada convergência superior do parietal), observada no Homo erectus, como do contorno mais circular, em vista posterior, dos Neandertais europeus do Plistocénico Médio e Superior (Arsu‑ aga et al., 1997). Quando comparado com a amostra de Atapuerca (SH), o crânio Aroeira 3 assemelha ‑se mais ao crânio 5 (1090 cm3), em termos dos diâme‑
tros transversais do osso temporal, e tem uma posi‑ ção intermédia, ou mais próxima do crânio 4 (1390 cm3), em relação ao resto das medições. Portanto, é
possível definir com segurança uma capacidade cra‑ niana acima dos 1100 cm3.
O osso temporal está quase completo e ostenta diversas características relevantes em termos fi‑ logenéticos. A apófise estilóide está unida ao ba‑ sicrânio. O tubérculo pós ‑glenóide é grande e de forma triangular. Em relação a estas características, o crânio da Aroeira distingue ‑se dos H. erectus asi‑ áticos e assemelha ‑se aos fósseis de Atapuerca (SH) (Martínez et al., 2008). Por outro lado, a eminência articular do crânio Aroeira 3 é saliente, não aplanada como nos crânios de Atapuerca (SH), Steinheim e Petralona, ou entre os Neandertais (Martínez et al., 2008). Em vista inferior, a apófise mastóide direita do crânio Aroeira 3 projecta ‑se bem para lá da sutu‑ ra occipitomastóidea, enquanto que na maioria dos Neandertais essa projecção, em geral, não ultrapassa a base do crânio (Martínez e Arsuaga, 1997). Quan‑ do medida a partir da incisura parietal, a projecção da apófise mastóide é no entanto reduzida, no que o crânio Aroeira 3 se assemelha aos dos Neandertais e de Steinheim (que é do Plistocénico Médio), e difere dos mastóides salientes que se vêem na colecção de Atapuerca (SH) (Martínez et al., 2008).
4. coNcLusÕes
O crânio Aroeira 3 possui diversas características típicas de crânios mais antigos do Plistocénico Mé‑ dio europeu. No entanto, quando tomadas em con‑ junto, a combinação de características que nele se vê não tem paralelo em nenhum outro indivíduo des‑ sas épocas. O seu toro supraorbital é contínuo e es‑ pesso, semelhante ao do crânio de Bilzingsleben, a sua apófise mastóide é curta, como no fóssil de Stei‑ nheim, e o seu tubérculo pós ‑glenóide é triangular, como nos fósseis de Atapuerca (SH) e de Steinheim. Estas características coexistem com uma eminência articular saliente, em contraste com as mais aplana‑ das geralmente presentes na amostra de Atapuerca (SH) e no crânio de Steinheim. Tem sido proposto que as eminências articulares aplanadas seriam uma característica que surge desde muito cedo na evolu‑ ção dos Neandertais (Arsuaga et al., 2014).
Os sítios da Aroeira, de Atapuerca (SH) e de Arago partilham uma relativa proximidade, tanto em ter‑ mos temporais (400–450 ka) como espaciais (Sudo‑ este europeu), mas os respetivos fósseis são clara‑ mente distintos. Essas diferenças sugerem que este período se caracteriza por uma diversidade intra ou interespecífica das populações humanas muito sig‑ nificativa e por dinâmicas demográficas complexas, incluindo processos de substituição, isolamento e miscigenação em grau diverso (Arsuaga et al., 2014). Aliás, tem sido argumentado que paleodemos arcai‑ cos (como, por exemplo, o representado pelo crânio de Ceprano) poderiam ter perdurado em refúgios eco ‑geográficos (Manzi et al., 2011) ao mesmo tem‑ po que, em outras regiões, se encontravam já paleo‑ demos mais evoluídos, com apomorfias neandertais (como, por exemplo, os representados por Atapuer‑ ca Sima de los Huesos) (Daura et al., 2017b).
É neste período que ocorrem duas grandes inova‑ ções tecnológicas: a expansão da indústria acheulen‑ se (Santonja e Villa, 2006), e as primeiras evidências da utilização generalizada, sistemática e controlada, do fogo (Roebroeks e Villa, 2011). Ambas estão do‑ cumentadas na Aroeira, cuja localização geográfica no extremo sudoeste da Europa sugere que essas inovações se espalharam rapidamente através do continente, independentemente das diferenças morfológicas que possam ter existido entre as suas populações humanas (ainda que, no caso da Europa Ocidental, a possibilidade de um fluxo genético ex‑ traeuropeu deva também ser tida em conta (Meyer
et al., 2016)). Fósseis bem datados e com contextos ecológicos e tecnológicos claros, como é o caso do crânio Aroeira 3, são cruciais para a construção de um quadro robusto da evolução humana na Europa durante o Plistocénico Médio .
AGRADecImeNtos
Agradecemos a J. Trueba a fotografia do crânio res‑ taurado. O TAC do crânio Aroeira 3 foi realizado no Parque Científico da Universidade de Burgos por R. Porres, em colaboração com J. M. Carretero e L. Rodríguez, do Laboratório de Evolução Humana da Universidade de Burgos. Esta investigação foi apoiada pelo Ministerio de Economía y Competi‑ tividad do Governo espanhol, através dos Projec‑ tos HAR2014 ‑55131, CGL2012 ‑38434 ‑C03 ‑01, e CGL2015 ‑65387 ‑C3 ‑2 ‑P (MINECO/FEDER). O tra‑ balho de campo foi financiado pela Câmara Munici‑ pal de Torres Novas e pela Fundação para a Ciência e Tecnologia, com o apoio logístico da Fábrica de Papel A Renova. J.D. foi financiado através de uma Bolsa de Pós ‑Doutoramento (SFRH/BPD/100507/2014) da Fundação para a Ciência e a Tecnologia, com fun‑ dos do Programa Operacional Potencial Humano do Fundo Social Europeu e um contrato Ramon y Cajal (RYC ‑2015 ‑17667), e M.S foi financiada atra‑ vés de um contrato de Pós ‑Doutoramento Juan de la Cierva (FJCI ‑2014 ‑21386).
bIbLIoGRAFIA
ARSUAGA, Juan‑Luis; MARTÍNEZ, Ignacio; ARNOLD, Lee J.; ARANBURU, Arantza; GRACIA‑TÉLLEZ, Ana; SHARP, Warren D.; QUAM, Rolf M.; FALGUÈRES, Chris‑ tophe; PANTOJA‑PÉREZ, Ana; BISCHOFF, James L: PO‑ ZA‑REY, Eva, PARÉS, Josep Maria; CARRETERO, José Miguel; DEMURO, Martina; LORENZO, Carlos; SALA, Nohemi; MARTINÓN‑TORRES, María; GARCÍA, Nuria; ALCÁZAR DE VELASCO, Almudena; CUENCA‑BESCÓS, Gloria; GÓMEZ‑OLIVENCIA, Asier; MORENO, Davinia; PABLOS, Adrian; SHEN, Chuan‑Chou; RODRÍGUEZ, Laura; ORTEGA, Ana Isabel; GARCÍA, Rebeca; BONMA‑ TÍ, Alejandro; BERMÚDEZ DE CASTRO, José María; CAR‑ BONELL, Eudald (2014) – Neandertal roots: Cranial and chronological evidence from Sima de los Huesos. Science. 344, pp. 1358–1363.
ARSUAGA, Juan‑Luis; MARTÍNEZ, Ignacio; GRACIA, Ana; LORENZO, Carlos (1997) – The Sima de los Huesos crania (Sierra de Atapuerca, Spain). A comparative study. Journal of Human Evolution. 33, pp. 219–282.
DAURA, Joan; SANZ, Montserrat; ARSUAGA, Juan‑Luis; HOFFMANN, Dirk. L.; QUAM, Rolf M.; ORTEGA, María Cruz; SANTOS, Elena; GÓMEZ, Sandra; RUBIO, Angel; VILLAESCUSA, Lucía; SOUTO, Pedro; MAURICIO, João; RODRIGUES, Filipa; FERREIRA, Artur; GODINHO, Paulo; TRINKAUS, Erik; ZILHÃO, João (2017a) – The Mid‑ dle Pleistocene Human Cranium from Gruta da Aroeira Acheulian site (Almonda Karst System,Torres Novas, Por‑ tugal). American Journal of Physical Anthropology. 162, pp. 157‑158.
DAURA, Joan; SANZ, Montserrat; ARSUAGA, Juan‑Luis; HOFFMANN, Dirk. L; QUAM, Rolf M.; ORTEGA, María Cruz; SANTOS, Elena; GÓMEZ, Sandra; RUBIO, Angel; VILLAESCUSA, Lucía; SOUTO, Pedro; MAURICIO, João; RODRIGUES, Filipa; FERREIRA, Artur; GODINHO, Pau‑ lo, TRINKAUS, Erik; ZILHÃO, João (2017b) – New Middle Pleistocene hominin cranium from Gruta da Aroeira (Por‑ tugal). Proceedings of the National Academy of Sciences. 114, pp. 3397‑3402.
DE LUMLEY, Marie‑Antoinette (2015) – L’homme de Tau‑ tavel. Un Homo erectus européen évolué. Homo erectus tautavelensis. L’Anthropologie. 119, pp. 303‑348.
DEAN, David; HUBLIN, Jean‑Jacques; HOLLOWAY, Ralph L.; ZIEGLER, Reinhard (1998) – On the phylogenetic posi‑ tion of the pre‑Neandertal specimen from Reilingen, Ger‑ many. Journal of Human Evolution. 34, pp. 485‑508.
FALGUÈRES, Christophe; YOKOYAMA, Yuji; SHEN, Guanjun; BISCHOFF, James L.; KU, Teh‑Lung; DE LUMLEY, Henry (2004) – New U‑series dates at the Caune de l’Arago, France. Journal of Archaeological Science. 31, pp. 941‑952. HOFFMANN, Dirk L.; PIKE, Alistair W.G; WAINER, Kar‑ ine, ZILHÃO, João (2013) – New U‑series results for the spe‑ leogenesis and the Palaeolithic archaeology of the Almonda karstic system (Torres Novas, Portugal). Quaternary Inter-national. 294, pp. 168‑182.
MANZI, Giorgio; MAGRI, Donatella; PALOMBO, Ma‑ ria Rita (2011) – Early‑Middle Pleistocene environmental changes and human evolution in the Italian peninsula. Qua-ternary Science Reviews. 30, pp. 1420‑1438.
MANZI, Giorgo, MALLEGNI, Francesco, ASCENZI, Anto‑ nio (2001) – A cranium for the earliest Europeans: phyloge‑ netic position of the hominid from Ceprano, Italy. Proceed-ings of the National Academy of Sciences. 98, pp. 10011‑10016. MARKS, Anthony E.; BRUGAL, Jean‑Philip; CHABAI, Vic‑ tor P.; MONIGAL, Katherine; GOLDBERG, Paul; HOCK‑ ETT, Bryan S.; PEMAN, Edurardo; ELORZA, Mikelo, MAL‑ LOLL, Carolina (2002) – Le gisement Pléistocène moyen de Galeria Pesada, (Estrémadure, Portugal): premiers résultats. Paléo. 14, pp. 77‑100.
MARTÍNEZ, Ignacio; ARSUAGA, Juan‑Luis (1997) – The temporal bones from Sima de los Huesos Middle Pleistocene site (Sierra de Atapuerca, Spain). A phylogenetic approach. Journal of Human Evolution. 33, pp. 283‑318.
MARTÍNEZ, Ignacio; QUAM, Rolf M.; ARSUAGA, Juan‑ Luis (2008) – Evolutionary trends in the temporal bone in the Neandertal lineage: a comparative study between the Sima de los Huesos (Sierra de Atapuerca) and Krapina samples. Em: MONGE, Janet M.; MANN, Alan E; FRAYER, David Wayne; RADOVCˇIC´, Jakov. (Eds.), New Insights on the Krapina Neandertals: 100 years after Gorjanovic´ – Kramberger. Croatian Natural History Museum, Zagreb., pp. 75‑80.
MEYER, Matthias; ARSUAGA, Juan‑Luis; DE FILIPPO, Cesare; NAGEL, Sarah; AXIMU‑PETRI, Ayinuer; NICK‑ EL, Birgit; MARTÍNEZ, Ignacio; GRACIA, Ana; BERMÚ‑ DEZ DE CASTRO, José María; CARBONELL, Eudald; VIOLA, Bence; KELSO, Janet; PRÜFER, Kay; PÄÄBO, Svante (2016) – Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins. Nature. 531, pp. 504‑507.
RIGHTMIRE, Philip G. (1996) – The human cranium from Bodo, Ethiopia: evidence for speciation in the Middle Pleis‑ tocene. Journal of Human Evolution. 31, pp. 21‑39.
RIGHTMIRE, Philip G. (1998) – Human evolution in the Middle Pleistocene: The role ofHomo heidelbergensis. Evo-lutionary Anthropology: Issues, News, and Reviews. 6, pp. 218‑227.
ROEBROEKS, Wil; VILLA, Paola (2011) – On the earliest evidence for habitual use of fire in Europe. Proceedings of the National Academy of Sciences of the United States of Ameri-ca. 108, pp. 5209‑5214.
SANTONJA, Manuel; VILLA, Paola (2006) – The Acheu‑ lian of Western Europe. Em: GOREN‑INBAR, Naama; SHARON, Gonen (Eds.), Axe Age – Acheulian Tool-making from Quarry to Discard. Equinox, London, pp. 429‑478. SMITH, Fred H.; RANYARD, Gail C. (1980) – Evolution of the supraorbital region in Upper Pleistocene fossil hominids from South‑Central Europe. American Journal of Physical Anthropology. 43, pp. 589‑610.
STRINGER, Chris (2012) – The status of Homo heidelber‑ gensis (Schoetensack 1908). Evolutionary Anthropology: Is-sues, News, and Reviews. 21, pp. 101‑107.
STRINGER, Chris; HUBLIN, Jean‑Jaques (1999) – New age estimates for the Swanscombe hominid, and their signifi‑ cance for human evolution. Journal of Human Evolution. 37, pp. 873‑877.
TRINKAUS, Erik; MARKS, Anthony E.; BRUGAL, Jean‑ Philip; BAILEY, Shara E; RINK, Jack W.; RICHTER, Daniel (2003) – Later Middle Pleistocene human remains from the Almonda Karstic System, Torres Novas, Portugal. Journal of Human Evolution. 45, pp. 219‑226.
ZILHÃO, João; MAURÍCIO, João; SOUTO, Pedro (1993) – Jazidas arqueológicas do sistema cársico da nascente do Al‑ monda. Nova Augusta. 7, pp. 35‑54.
Figura 1 – O sítio da Gruta da Aroeira. 1: Localização geográfica. 2: O Maciço Calcário Estremenho, com a posição da nascente do Almonda, o rio Tejo e a Serra d’Aire. 3: A escarpa do Almonda, com a posição da Gruta da Aroeira e o rio Almonda em primeiro plano. 4: Área de escavação da Gruta da Aroeira e entrada da gruta. 5 ‑6: Cortes estratigráficos, vistas S e N, respectivamente. 7: Planta da Gruta da Aroeira.
Figura 2 – O crânio original Aroeira 3, completamente restaurado, vista endocraniana (1) e lateral (2). Recons‑ trução virtual do crânio Aroeira 3 em vista frontal (3), posterior (4), superior (5) e endocraniana (6).