Palynological evidence of vegetation change during the last 10,000 years in the mid-valley of the Rio Doce, Brazil
Texto
Imagem


Documentos relacionados
O fato é que esses leitores de folhetins com moldes franceses buscavam, cada um a seu modo e com suas exigências, “reencontrar sua existência pela auto-idealização a
Ao Dr Oliver Duenisch pelos contatos feitos e orientação de língua estrangeira Ao Dr Agenor Maccari pela ajuda na viabilização da área do experimento de campo Ao Dr Rudi Arno
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
IJKLJMLNOPQRJMSNLJKSTURLSLPVOTSMLWOXLQSYOZL[SLNKOMOPOP J\YL]PLV]VTOPMLLJP]JORNSVRTKRZS^ZONOKOZSMLSK]KRMLP_VS^SV]MSMLPL
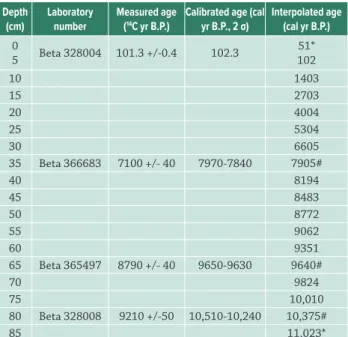
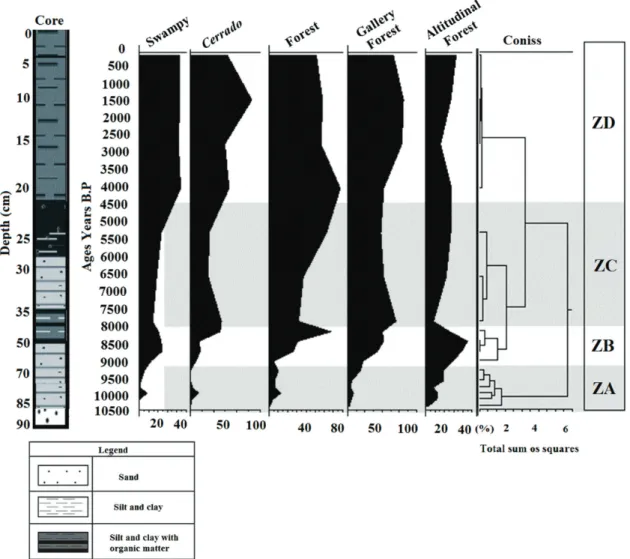
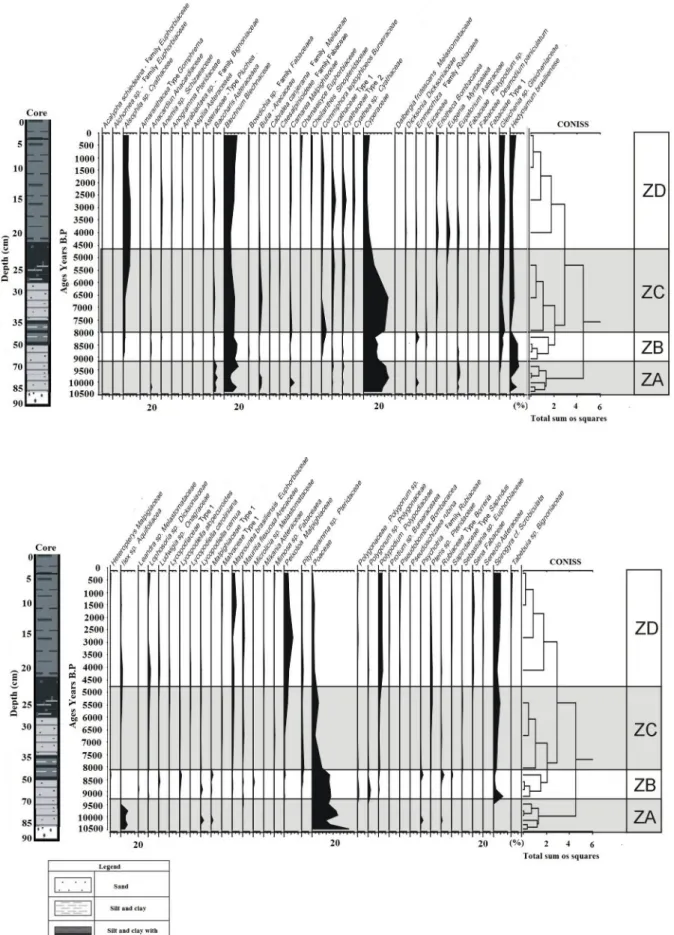
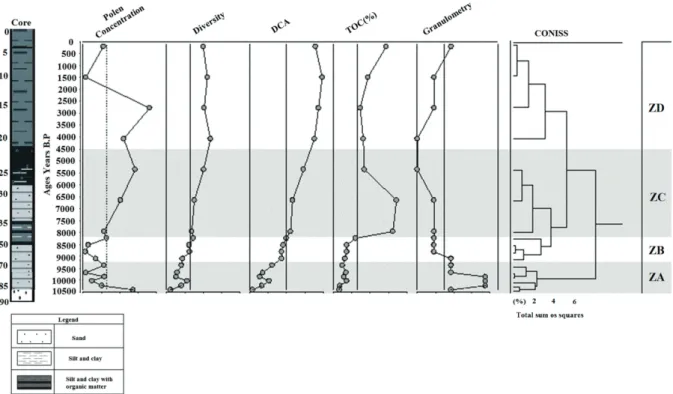
With the aim of reconstructing the temporal dy- namics of the vegetation during the last 7,000 years, correlated to climatic changes and the sea level oscilla- tion,
The probability of attending school four our group of interest in this region increased by 6.5 percentage points after the expansion of the Bolsa Família program in 2007 and
O objetivo deste estudo foi identificar as incompatibi- lidades físico-químicas entre medicamentos administrados por via intravenosa em pacientes internados no Centro de
Transformou-se em galinha como as outras, apesar das asas de quase três metros de extensão.. A ÁGUIA E
