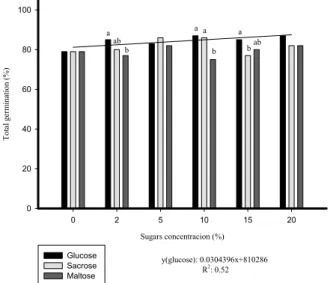
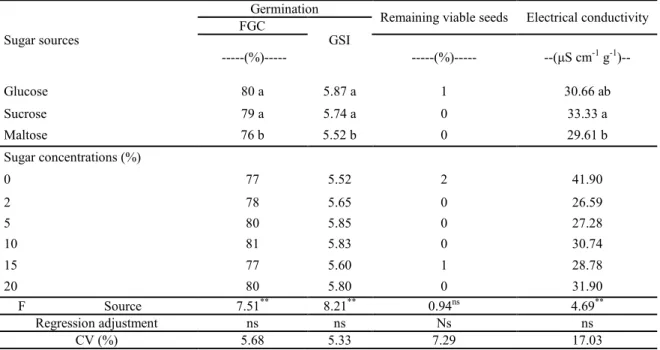
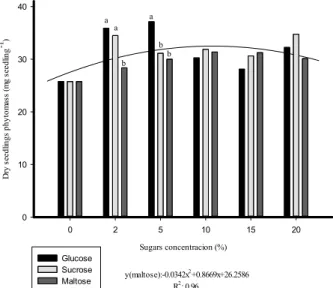
PRIMING OF BRACHIARIA SEEDS WITH DIFFERENT SUGAR SOURCES AND CONCENTRATIONS
Texto
Imagem
Documentos relacionados
Para Cassiolato e Lastres (2000) além de intensa importação de tecnologia, neste período as atividades de CT&I (assim como outros países da América Latina)
The aim of this study was to evaluate the effect of supplementation with different concentrations of reduced glutathione GSH (0; 5; 7.5; 10mM) in the extender
To evaluate the effect of preconditioning of maize seeds in ozonated water on seed physiological quality, germination test (GT) was conducted and the following variables were
The aim of this study was to evaluate the effect of foliar application of manganese, with different doses and stages of application, on the physiological quality of soybean seeds
Como o método utilizado foi o etnográfico e já foi referido anteriormente que se fez trabalho de campo, houve o deslocamento até a Ilha de São Vicente, onde se passou
Figura 20 - Aminograma do material coletado do lado seroso dos segmentos intestinais evertidos, evidenciando a absorção de S-metilcisteína proveniente do
O presente estudo teve como objetivo a caracterização morfológica e física de solos com horizontes subsuperficiais adensados, visando melhor entendimento sobre os processos
na Floresta Semidecídua do México, tornou-se fértil durante o período de estudo (31 meses), entretanto, duas plantas próximas a área de estudo (500 m) apresentaram
