UNIVERSIDADE FEDERAL DO CEARÁ CENTRO DE CIÊNCIAS AGRÁRIAS DEPARTAMENTO DE ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ZOOTECNIA
ANTÔNIO NÉLSON LIMA DA COSTA
SINCRONIZAÇÃO DO ESTRO E DA OVULAÇÃO EM NOVILHAS GIROLANDAS: COMPARAÇÃO ENTRE DOIS PROTOCOLOS HORMONAIS
“CIDR-B” E “OVSYNCH”
ANTÔNIO NÉLSON LIMA DA COSTA
SINCRONIZAÇÃO DO ESTRO E DA OVULAÇÃO EM NOVILHAS GIROLANDAS: COMPARAÇÃO ENTRE OS PROTOCOLOS HORMONAIS
“CIDR-B” E “OVSYNCH”
Dissertação submetida à Coordenação do Curso de Pós-Graduação em Zootecnia, da Universidade Federal do Ceará, como requisito parcial para a obtenção do Título de Mestre em Zootecnia – Área de Concentração: Reprodução de Bovinos.
Orientador: Prof. Dr. Aírton Alencar de Araújo
ANTÔNIO NÉLSON LIMA DA COSTA
SINCRONIZAÇÃO DO ESTRO E DA OVULAÇÃO EM NOVILHAS GIROLANDAS: COMPARAÇÃO ENTRE OS PROTOCOLOS HORMONAIS
“CIDR-B” E “OVSYNCH”
Dissertação submetida à Coordenação do Curso de Pós-Graduação em Zootecnia, da Universidade Federal do Ceará, como requisito parcial para a obtenção do Título de Mestre em Zootecnia – Área de Concentração: Reprodução de Bovinos.
Aprovada em ____/____/_______
BANCA EXAMINADORA
_________________________________________________ Prof. Dr. Aírton Alencar de Araújo (Orientador)
Universidade Federal do Ceará – UFC
_______________________________________________ Profa. Dra. Ana Cláudia Nascimento Campos (Conselheira UFC)
Universidade Federal do Ceará – UFC
_____________________________________________ Prof. Dr. Davide Rondina (Conselheiro UECE)
Às minhas queridas Tia Cícera e Avó Adaltiva (in memorian).
OFEREÇO
Aos meus pais, meus irmãos, à Mirna e aos animais.
AGRADECIMENTOS
Agradeço, em primeiro lugar, ao nosso Pai Santo Deus Todo Poderoso, pois sem Ele nada seríamos e a todos os Santos que me auxiliaram em mais esta conquista.
Agradeço aos meus queridos pais, Manoel Nélson da Costa e Maria do Socorro Lima da Costa, por terem me dado inúmeras oportunidades na vida e por estarem sempre ao meu lado em todos os momentos.
Agradeço ao meu irmão Eduardo, sempre disposto a me ajudar nos momentos mais difíceis de minha vida, e às minhas irmãs Cristiane e Antônia, grandes companheiras.
Agradeço a todos os meus familiares que me apoiaram, em especial ao meu avô Antônio Ferreira e ao meu tio Cícero Costa, que são pessoas humanas de verdade.
Agradeço à minha namorada, Mirna Fontenele, por toda paciência, dedicação e carinho que tem por mim, sempre estando disposta a me ajudar sem medir esforços.
Agradeço ao meu orientador Professor Dr. Aírton Alencar por todo o apoio prestado na execução deste trabalho, pelos ensinamentos técnicos e de vida, os quais levarei comigo para sempre, além de sua paciência e generosidade.
Agradeço ao amigo Dr. Péricles Montezuma, o qual foi de fundamental importância na realização deste trabalho, se empenhando para conseguir os animais e cedendo parte de seu tempo tão restrito para nos ajudar nos trabalhos de campo.
Agradeço aos membros da banca examinadora, Professora Dra. Ana Cláudia e Professor Dr. Davide Rondina, pela colaboração e disponibilidade.
Agradeço a CIALNE, na pessoa do Sr Rafael Carneiro, pela concessão dos animais e das instalações para a execução do experimento.
Agradeço aos funcionários da Fazenda Recife (CIALNE XVIII) pela ajuda nos trabalhos de campo, em especial ao Sr Juscelino, ao Pedro, ao José Augusto, ao Mainha, ao Chiquinho, ao Jorge e ao Jessé.
Agradeço a todos os professores da Pós-Graduação da UFC pelos ensinamentos prestados durante todo o curso, em especial a professora Sônia pela sua boa vontade em ajudar.
Agradeço a todos os colegas de Mestrado pela ótima convivência durante o curso, em especial meu companheiro de área Jorge André, ao grande Ednir, ao Társio, a Irani, ao Roberto, a Thaísa, a Eva, a Rossana, ao Josemir, ao Bruno, ao Severino, ao Eduardo e Alethéia, a Ludmila e ao Luciano.
Agradeço ao Dr Arlindo Moura, que mesmo não estando presente, ajudou no preparo deste trabalho.
Agradeço aos amigos de todas as horas Adílio e Adson e todos os outros que estão sempre nos ajudando nas horas mais complicadas, como o Dr. João Alves.
Agradeço a DISVET, na pessoa do Sr Glaudêmio, pela ajuda nos hormônios do laboratório INTERVET e pela grande boa vontade e paciência que o mesmo nos demonstrou.
Agradeço ao laboratório PFIZER, divisão de saúde animal, nas pessoas do Sr Alexandre, Armando e Oclydes, pelo apoio incondicional, tanto na doação dos dispositivos “CIDR-B”, quanto no apoio prestado nos congressos.
Estudar, trabalhar e conquistar, sempre! Perder a humildade, nunca!
RESUMO
ABSTRACT
Two oestrus and ovulation synchronization protocols (CIDR-B and OVSYNCH) were tested on 90 acyclical Girolanda heifers. The animals were distributed in three groups. Group I (n=30) was submitted to the OVSYNCH protocol, consisting of: intramuscular (IM) application of 1.0 ml gonadotropin-releasing hormone (GnRH), 2.0 ml prostaglandin F2α (PGF2α) after 7 days, 1.0ml GnRH 48h later, then timed artificial insemination (TAI) 16h after the second application of GnRH. Group II (n=30) was submitted to the CIDR-B protocol, consisting of: introduction of intravaginal CIDR-B implant and application of 2.0 ml estradiol benzoate (EB) IM, implant removal after 8 days, application of 500 UI pregnant mare serum gonadotropin (PMSG) and 2.0 ml PGF2α, a second IM application of BE 24h subsequently, then TAI 30h later (i.e. 54h following implant removal). Group III (n=30; control group) received no hormonal treatment. Gestation was verified ultrasonographically 35 days after the first, second and third AI. The cost-benefit ratio of each protocol was also determined: The CIDR-B protocol was most efficient in the first AI in which it was comparable to the control group in providing a near-natural oestrus quality (conception and pregnancy rates of 60% and 50%, respectively; p<0.05). In the second AI, the two treatments
LISTAS DE ABREVIATURAS E SÍMBOLOS
Cds – Custo da Dose do Sêmen
CE – Ciclo Estral
CIDR-B – Controlled Internal Drug Releasing-Bovine
CL – Corpo Lúteo
Ct – Custo do Tratamento
E2 – Estrógeno
ECC – Escore de Condição Corporal
eCG – Gonadotrofina Coriônica Eqüina
FD – Folículo Dominante
FDA – Food and Drug Administration
Fig. - Figura
FO – Folículo Ovulatório
FS – Folículo Subordinado
FSH – Hormônio Folículo Estimulante
FUNCEME – Fundação Cearense de Meteorologia
GnRH – Hormônio Liberador de Gonadotrofinas
h – Hora
hCG – Gonadotrofina Coriônica Humana
IA – Inseminação Artificial
IGF-I – Fator de Crescimento da Insulina – 1
IIP – Intervalo IA até o Parto
IM – Intra-Muscular
IPPP – Idade Provável ao Primeiro Parto
kg - Quilograma
LH – Hormônio Luteinizante
MGA – Acetato de Melengestrol
ml – Mililitros
mm - Milímetros
n – Número de animais inseminados
N – Número de animais submetidos ao tratamento
Nia – Número de Inseminações Artificiais
Np – Número de Fêmeas Prenhes ao Diagnóstico de Gestação
P4 - Progesterona
PB – Proteína Bruta
PGF2α - Prostaglandina F2α
pg/ml – Picograma por ml
PMSG – Gonadotrofina Sérica da Égua Prenhe
PPP14 – Probabilidade de Prenhez de Novilhas aos 14 meses
Tab - Tabela
Tx - Taxa
LISTAS DE FIGURAS, TABELAS E ANEXOS
Página Fig. 1. Esquema das trocas hormonais no eixo
Hipotálamo-Hipófise-Ovários-Útero...16
Fig. 2. Diagrama de crescimento folicular durante o ciclo estral de 21 dias. Animal com 2 ondas de crescimento folicular...19
Fig. 3. Concentrações hormonais durante o crescimento folicular e ovulação no CE de uma fêmea bovina...23
Fig. 4. Esquema do protocolo “OVSYNCH”...35
Fig. 5. Esquema do protocolo “CIDR-B”...36
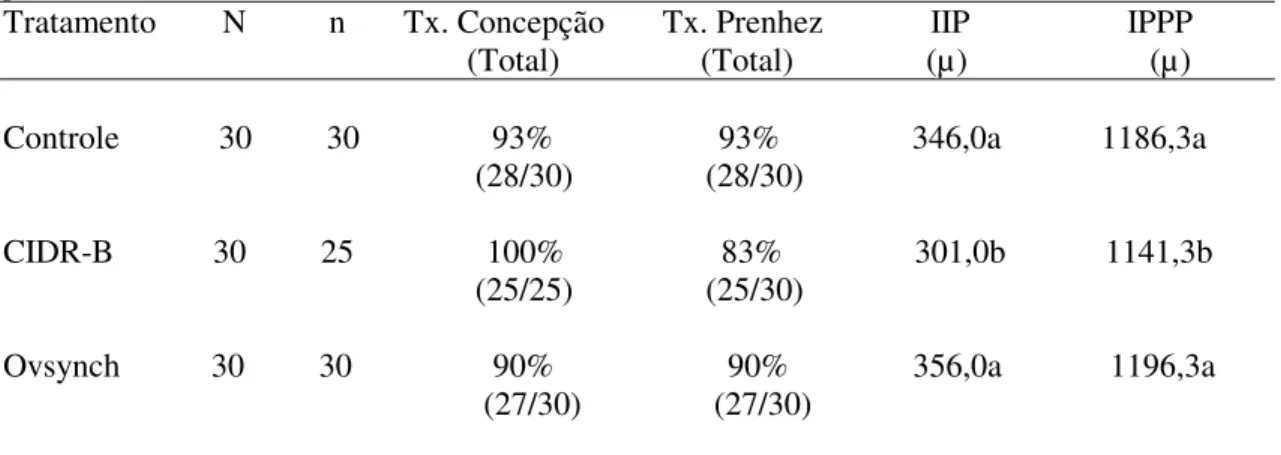
Tab. 1. Taxas de concepção, prenhez, e idades prováveis ao primeiro parto (IPPP) médias das novilhas que emprenharam na 1ªIA nos três tratamentos...40
Tab. 2. Taxas de concepção e prenhez totais, intervalo IA até o parto (IIP) e idade provável ao primeiro parto (IPPP) em médias totais...41
Tab. 3. Número de IAs necessárias para emprenhar uma novilha...41
Anexo-1. Novilhas Girolandas (3/8 Holandês X 5/8 Gir) utilizadas no experimento...65
Anexo-2. Avaliação ginecológica dos animais por palpação retal...65
Anexo-3. Preparação do implante “CIDR-B”...66
Anexo-4. Implante “CIDR-B” dentro do aplicador pronto para a introdução na vagina...66
Anexo-5. Implante “CIDR-B” dentro da vagina e fio de remoção exteriorizado na vulva..67
Anexo-7. Animais apresentando sinais de estro após o término dos tratamentos
hormonais...68
Anexo-8. Novilha apresentando muco vulvar pouco tempo antes da IATF...68
Anexo-9. Inseminação artificial em tempo fixo dos animais experimentais...69
Anexo-10. Diagnóstico de gestação por ultra-sonografia...69
Anexo-11. Equipe que participou do experimento...70
SUMÁRIO
Página
RESUMO...VII ABSTRACT...VIII LISTA DE ABREVIATURAS E SÍMBOLOS...IX LISTA DE FIGURAS, TABELAS E ANEXOS...XI
1. INTRODUÇÃO...01
2. REVISÃO DE LITERATURA ...04
2.1. Puberdade...04
2.1.1. Conceito...04
2.1.2. Determinação da puberdade...05
2.1.3. Fatores que influem na idade a puberdade...05
2.1.3.1. Fatores nutricionais...06
2.1.3.2. Fatores genéticos...07
2.1.3.3. Fatores sazonais...10
2.1.3.4. Efeito da presença do macho...11
2.1.4. Estratégias para reduzir a idade à puberdade...12
2.1.5. A puberdade em rebanhos mestiços (Bos indicus X Bos taurus)...12
2.2. Fisiologia reprodutiva da fêmea bovina...14
2.2.1. A regulação...14
2.2.2. Órgãos e sistemas liberadores de hormônios...14
2.2.2.1.Hipotálamo...14
2.2.2.2. Hipófise...15
2.2.2.3. Ovários...15
2.2.2.4. Útero...15
2.3. Crescimento folicular e ovulação...16
2.4. Hormônios relacionados com a onda de crescimento folicular...19
2.4.1. Hormônio liberador de gonadotrofinas (GnRH)...19
2.4.2. Hormônio folículo estimulante (FSH)...20
2.4.3. Hormônio luteinizante (LH)...21
2.5. Inseminação artificial em tempo fixo (IATF)...23
2.6. Os protocolos de IATF...25
2.6.1. Sincronização usando PGF2α...25
2.6.2. Sincronização usando GnRH-PGF2α (OVSYNCH)...26
2.6.3. Sincronização usando progestágenos (CIDR-B)...28
3. JUSTIFICATIVA...32
4. OBJETIVOS...33
4.1. Objetivo geral...33
4.2. Objetivos específicos...33
5. MATERIAL E MÉTODOS...34
5.1. Localização do experimento...34
5.2. Animais experimentais...34
5.3. Delineamento experimental...34
5.3.1. Protocolo “OVSYNCH”...35
5.3.2. Protocolo “CIDR-B”...35
5.4. Análise estatística...36
5.5. Análise econômica...36
6. RESULTADOS E DISCUSSÃO...38
7. CONCLUSÕES...44
8. REFERÊNCIAS BIBLIOGRÁFICAS...45
1. INTRODUÇÃO
Por ser um país de dimensões continentais e apresentar condições climáticas e edáficas favoráveis, o Brasil destaca-se mundialmente na criação bovina possuindo um rebanho com mais de 191 milhões de cabeças (MUSTEFAGA & NOGUEIRA, 2004). Sendo que isto só foi obtido após investimentos em áreas primordiais para o sucesso pecuário, às quais podem ser resumidas no trinômio: nutrição, genética e saúde animal. Com isso, as exportações de carne bovina tiveram um avanço considerável nos últimos anos, ou seja, em 1998 o país exportava cerca de 6% de sua produção, o que em 2004 chegou a quase 20% do total produzido (ANDRADE, 2004).
A pecuária tem apresentado ganhos satisfatórios nas características economicamente desejáveis, o que se deve principalmente à seleção e à difusão do uso de reprodutores geneticamente superiores através da inseminação artificial. A dificuldade de prever o momento do estro de vacas ou novilhas em um certo grupo pode inviabilizar o uso da inseminação artificial (IA), pois haverá um alto custo com mão de obra para detecção do estro. Como solução para melhorar os índices reprodutivos, temos os protocolos de sincronização do estro e ovulação, os quais incluem a sincronização do estro em fêmeas ciclando e a indução do estro acompanhado de ovulação em novilhas que ainda não atingiram a puberdade ou em vacas que não voltaram a apresentar estro após a parição (LAMB, 2003).
mais eficiente de sincronização de estro em bovinos. Assim, vários protocolos foram desenvolvidos com este propósito (GORDON, 1996).
Seguindo esta linha, o uso de um protocolo adequado que permita o uso racional da IA em tempo pré-fixado, pode trazer benefícios múltiplos para o produtor, tais como a melhoria da eficiência reprodutiva do rebanho e a redução da mão de obra, além de um maior controle do nascimento dos bezerros que irá facilitar a obtenção de uma homogeneidade da produção leiteira ao longo do ano e uma maior lucratividade para o produtor rural (GONSALVES, 2002).
Porém, para se obter sucesso e evitar problemas no uso da sincronização do estro algumas condições devem ser respeitadas nas fêmeas selecionadas para este propósito, às quais são: intervalo de tempo adequado parição-sincronização (mínimo de 40 dias pós-parto); vacas com escore corporal satisfatório (>3); ausência de problemas pós-parto anteriores e novilhas apresentando peso corporal de, no mínimo, 65% do peso adulto final (PATTERSON et al., 1999).
O uso de progestágenos objetivando a sincronização de estros em bovinos provém dos anos 50, sendo, de início, administrados por períodos de 11-14 dias. Com o passar do tempo, este intervalo diminuiu para 7-9 dias devido aos baixos índices de fertilização após a sincronização e, deste modo, obtiveram-se melhores índices de gestação (GORDON, 1996). A função básica dos progestágenos na sincronização é aumentar a vida útil do corpo lúteo e, conseqüente estro, em um mesmo período. Os métodos utilizados para a administração dos progestágenos são os seguintes: injeção em óleo; administração no alimento (MGA); implantes subcutâneos; esponjas intra-vaginais; dispositivo de espiral de plástico injetado para uso intra-vaginal (PRID) e aparato de plástico injetado em forma de “Y” para uso intra-vaginal (CIDR-B) (MORAES et al., 2002).
Outro hormônio muito utilizado em protocolos de sincronização é o GnRH, porém, inicialmente, era indicado para bovinos com cistos (SEGUIN et al., 1978).
Posteriormente, foi descoberta a valiosa utilidade deste hormônio para o controle da dinâmica folicular e luteal. TWAGIRAMUNGU et al. (1992) fizeram o relato de que
3*). WDPEém melhorou a taxa de sincronização em vacas leiteiras em lactação
(THATCHER et al., 1993).
As conseqüências do uso do GnRH em programas de reprodução foram mais bem esclarecidas após o desenvolvimento do protocolo de inseminação programada “OVSYNCH”, onde combinam-se tratamentos que visam o controle tanto da dinâmica folicular como luteal, fazendo com que o tempo de ovulação possa ser sincronizado com exatidão (PURSLEY et al., 1995). Após a sua criação, este protocolo vem sendo
utilizado largamente na indústria leiteira (WILTBANK & HAUGHIAN, 2003). Seu princípio consiste em provocar a ovulação e formação de um corpo lúteo ou luteinização do folículo dominante, que sofrerá posterior luteólise com a aplicação da
3*). $ VHJXQGD DSOLFDção de GnRH objetiva desencadear o processo ovulatório e,
deste modo, a ovulação sincronizada (MORAES et al., 2002).
Desta forma, este protocolo é bastante utilizado para a indução da ovulação ou luteinização de vacas em anestro. Nestas vacas, a exposição à progesterona atua de forma a evitar um ciclo curto após a segunda injeção de GnRH. Relatou-se também que a combinação de GnRH com progesterona, conhecido como “CIDR-SYNCH”, também melhora as taxas de prenhez de bovinos leiteiros que se encontram em anestro (PURSLEY et al., 2001).
O protocolo “CIDR-B” consiste em um dispositivo intra-vaginal com progesterona junto com outros hormônios (Benzoato de Estradiol e PMSG) em forma de “T”, que apresenta abas flexíveis dobráveis formando uma haste para ser inserida na vagina com o auxílio de aplicador, possuindo ainda na extremidade oposta às abas um cordão que facilita sua retirada. O suporte principal do “CIDR-B” é formado de nylon coberto por silicone impregnado com progesterona (1,9g), cujas concentrações sangüíneas aumentam rapidamente após a inserção, atingindo um pico máximo em uma hora (LAMB, 2003).
2. Revisão de Literatura
2.1. Puberdade:
2.1.1. Conceito:
É o momento em que o animal encontra-se apto à reprodução, ou seja, o início de sua vida reprodutiva (BLOOD & STUDDERT, 2002; CUNNINGHAM, 1999; REECE, 1996). Visto que, alguns parâmetros ainda precisam ser alcançados para que o animal torne-se púbere, tais como peso e idade que, nos bovinos, é bastante variável de acordo com a raça, sendo em torno dos 275kg e dos 8 a 12 meses de idade (CUNNINGHAM, 1999). RAWLINGS et al. (2003) caracterizam o início da puberdade
pela primeira ovulação e MORAN et al. (1989) relataram sua ocorrência entre os 6 e 24
meses de idade dependendo de fatores genéticos e ambientais. Com animais mestiços, PERRY et al. (1991) constataram o início da puberdade aos 11 meses de idade.
Fisiologicamente, a puberdade pode ser definida como o momento da manifestação do primeiro estro em conjunto com uma ovulação potencialmente fértil, desenvolvimento de corpo lúteo e uma fase luteal de duração normal (MARSON et al.,
2004). O animal só é púbere quando está apto a liberar seus gametas e exibir comportamento sexual, sendo o início da puberdade marcado pela maturação do eixo hipotalâmico-adeno-hipofisário (HAFEZ & HAFEZ, 2004). O declínio progressivo na resposta hipotalâmica ao estradiol permite um aumento na estimulação pituitária, à qual reflete em altos níveis de secreção de LH (DAY et al., 1987).
respectivamente. A maturidade sexual de novilhas envolve um aumento na circunferência ovariana, à qual é mais evidenciada por volta dos 13 meses de idade (LOBO et al., 2001). Além disso, tanto novilhas de raças zebuínas quanto européias,
mostram um aumento na secreção de LH enquanto a de FSH permanece inalterada (EVANS et al., 1994; NOGUEIRA et al., 2003).
2.1.2. Determinação da puberdade:
A idade à puberdade das fêmeas bovinas pode ser determinada de várias maneiras considerando os diversos fatores fisiológicos envolvidos, mas ROMANO (1997) e MARSON (2000) sugerem que a idade a puberdade e o ciclo estral subseqüente devem ser diagnosticados por meio de imagens ultrassonográficas ou por detecção dos níveis sanguíneos de progesterona (P4) através de radioimunoensaio (RIA).
RODRIGUES et al. (2002) caracterizaram a idade a puberdade como sendo a
idade da fêmea no dia em que um corpo lúteo foi observado pela primeira vez através de ultra-sonografia, aliado a sua funcionalidade comprovada pelas concentrações plasmáticas de P4 acima de 1ng/ml, mantidas durante 2-3 coletas de sangue consecutivas e com intervalos de 3-4 dias. Estas coletas devem obedecer uma periodicidade: serem iniciadas antes da fêmea apresentar o primeiro estro e, semanalmente serem realizadas até a detecção deste; a partir daí as mesmas deverão ser feitas a cada 3-4 dias até a manifestação do terceiro estro, caracterizando, assim, a maturidade sexual (MARSON et al., 2004).
No entanto, devido à dificuldade de se fazer esses procedimentos anteriores e do seu alto custo, muitos autores definem o momento de manifestação da puberdade como sendo a observação do primeiro estro (FERREL, 1982; ALENCAR et al., 1987).
Porém, na realidade, dificilmente os sistemas produtivos observam o primeiro estro das novilhas, principalmente quando estes são extensivos, aonde se tem apenas uma estimativa através da idade ao primeiro parto (MARSON et al., 2004; NOGUEIRA,
2.1.3. Fatores que influenciam na idade à puberdade:
A idade à puberdade é um importante aspecto produtivo em novilhas que são selecionadas para parir aos 2 anos de idade em bom estado corporal, melhorando sua vida reprodutiva, sem prejudicar sua longevidade ou ganho de peso quando comparadas às novilhas que parem aos 3 anos de idade (TRAN et al., 1988). A precocidade sexual
das fêmeas bovinas é avaliada através da idade à puberdade ou ao primeiro parto, sendo, desta maneira, um dos principais métodos para se mensurar e elevar a eficiência reprodutiva de um rebanho, pois este é provavelmente o melhor parâmetro de avaliação de fêmeas (MARSON et al., 2004).
Desta forma e, devido à alta herdabilidade (0,31-0,50) para esta característica, sua utilização na seleção de rebanhos é de fundamental importância (LASTER et al.,
1979; MARTIN et al., 1992). A idade à puberdade estimada para zebuínos nos trópicos
e sub-trópicos varia de 16-40 meses. Isto é atribuído a fatores genéticos e ambientais, destacando-se os nutricionais, sazonais e outros como a presença do macho (MUKASA MUGUERVA, 1989; SOUZA et al., 1995; PEREIRA; 2000; MARSON et al., 2004).
2.1.3.1. Fatores nutricionais:
Os fatores nutricionais estão inversamente relacionados com a idade à puberdade das novilhas, ou seja, quanto melhor o nível nutricional que uma novilha é submetida, menor será o número de dias para esta novilha tornar-se púbere (SCHILLO
et al., 1992). Melhorias na nutrição pós-desmame é um fator importante na diminuição
da idade à puberdade e ao primeiro parto (PATTERSON et al., 1992). Inicialmente
considerava-se uma novilha púbere com 40-50% do seu peso corporal adulto (SCHILLO et al., 1992), mas SPIRE (1997) propôs de 60-65% do peso corporal adulto
como sendo ideal à puberdade.
275kg e, consequentemente, elevou-se a taxa de concepção das novilhas no primeiro serviço de 5,8 para 45,5%, sendo refletido numa melhora na produção leiteira e confirmando que, a puberdade precoce é atingida tanto por um bom peso a desmama, quanto por um ganho significativo após esta.
Todavia BAGLEY (1993) adverte que ganhos subseqüentes de peso devem existir nos rebanhos para que se possa garantir, às novilhas púberes, a continuidade de ciclos estrais normais. Quando se têm condições ambientais desfavoráveis pós-desmame, deve ser feita uma diferenciação na alimentação, dando preferência às novilhas que se encontrem com escore mais baixo para que seja atingido um peso ideal à puberdade (BERETTA et al., 1996). Além disso, mesmo as novilhas que só obtiveram
melhores ganhos de peso nos últimos momentos da fase de desenvolvimento corporal, não serão prejudicadas nos seus desempenhos reprodutivos na primeira estação de monta (LYNCH et al., 1997).
Ao se promover um manejo nutricional adequado de novilhas, percebe-se melhoras tanto na eficiência reprodutiva, como um melhor funcionamento da endocrinologia reprodutiva das mesmas (MARSON et al., 2004). A manutenção da
elevada freqüência pulsátil de liberação de LH está diretamente relacionada à reserva energética do animal. Logo, se o animal sofrer restrição nutricional prolongada atrasará o início da puberdade devido à supressão da liberação do LH em pulsos com freqüência elevada, que são de fundamental importância para o crescimento folicular até o estágio pré-ovulatório (YELICH et al., 1996).
Além da diminuição na freqüência dos pulsos de LH, WILLIAMS et al. (2002)
relataram que a restrição nutricional pode provocar uma redução nas concentrações circulantes do hormônio leptina, que atua como regulador do metabolismo energético e no desenvolvimento do sistema reprodutivo, inclusive na puberdade, estando o peso corporal altamente correlacionado (R=0,85) com a leptina circulante. Os níveis de progesterona (P4) também são afetados quando o animal sofre restrição nutricional na fase de crescimento, mesmo se esta restrição for corrigida próximo à puberdade (PERON & FERNANDEZ, 1986).
reprodutivo de uma fêmea bovina (MARSON et al., 2004). FERREL (1982) e
MARSON (2000) concluíram que a condição corporal tem influência sobre o peso médio à puberdade e, desta forma, auxilia na previsão da idade e do peso da novilha no momento da puberdade.
2.1.3.2. Fatores genéticos:
De uma forma mais abrangente, os fatores genéticos interagem com os ambientais, mas destacam-se destes no que se refere a composição racial de uma fêmea, ou seja, a forma mais simplificada de se verificar a influência genética sobre a puberdade é comparando a idade média à puberdade entre as diversas raças de uma mesma espécie, ou melhor, as novilhas possuem uma idade mínima geneticamente pré-determinada em que conseguem atingir a puberdade, sendo esta uma fonte de variação altamente significativa (FERREL, 1982; HESS, 2002).
Em um rebanho de novilhas da raça nelore, NOGUEIRA et al. (2003) relataram
a presença de um fator genético envolvido na precocidade de 30% dos animais deste rebanho, que se tornaram gestantes aos 16 meses de idade, mesmo não existindo diferenças na idade ou no peso dos animais do grupo à primeira ovulação. O intervalo entre a primeira e a segunda ovulação, no entanto, foi maior para as novilhas precoces (12 dias) do que para as não precoces (9 dias), o que sugere um grau de maturidade diferente no hipotálamo nos dois tipos de animais.
As diferenças raciais podem ser usadas para a obtenção de ótimos níveis de desempenho para diversas características produtivas (GREGORY & CUNDIFF, 1999). Assim, pode-se combinar estas diferenças objetivando seus produtos e se beneficiando dos efeitos positivos da heterose sobre várias características produtivas e reprodutivas (MARSON et al., 2004).
Com relação aos zebuínos, a interação genótipo-ambiente tem um efeito significante sobre o peso pós-desmama e, desta forma, para a ocorrência da primeira ovulação. Assim, o crescimento é restrito se a novilha não é bem alimentada e isto leva a uma elevação na idade à puberdade. Um aumento na produtividade pode ser alcançado pela introdução de um manejo nutricional adequado para redução da idade à puberdade em rebanhos que tenham altas taxas de parição (BERETTA et al., 2001;
FERREIRA et al., 2001).
Quando é feita a escolha de raças para a composição de um rebanho, deve-se lembrar que as diferenças apresentadas entre elas para a idade à puberdade das fêmeas não provêm da estrutura corporal adulta ou da taxa de crescimento, mas da aptidão para a qual são selecionadas (FERREL, 1982). Desta maneira, raças aptas para a produção de leite atingem a puberdade mais precocemente do que as de mesma estrutura corporal, mas que não são selecionadas para este tipo de produção (LASTER et al., 1979;
MARTIN et al., 1992). Raças de maior porte, como a charolesa e a chianina, tendem a
ser mais tardias e pesadas à puberdade do que as de menor porte, como angus e hereford (MARTIN et al., 1992; BAGLEY, 1993), sendo a correlação entre tamanho
corporal e idade à puberdade de 0,57 para as raças européias e 0,25 para as indianas (MARTIN et al., 1992).
Segundo MARSON et al. (2004) a importância dos fatores genéticos sobre a
manifestação da puberdade evidencia-se, de forma mais clara e objetiva, quando é determinada a herdabilidade para a característica, ou seja, a herdabilidade para a idade à puberdade varia de 0,31 a 0,50, significando que esta idade pode ser diminuída através da seleção genética, escolhendo-se animais mais precoces (LASTER et al., 1979;
MARTIN et al., 1992; SPLAN et al., 1998). No entanto, a caracterização dos critérios
de seleção genética para redução da idade à puberdade são de difícil mensuração, principalmente em criações extensivas, devido à dificuldade para a coleta de dados (ALBUQUERQUE & FRIES, 1997; BERGMANN, 1999).
características de órgãos reprodutivos como: tônus e tamanho uterino, tamanho dos ovários, crescimento folicular e presença de corpo lúteo; quando analisados em novilhas antes da estação de monta, são úteis na identificação das fêmeas com alta probabilidade de ovular (FERREIRA et al., 1999).
Por se mostrar um parâmetro de fácil medição, alta herdabilidade (de 0,46 a 0,48) e as correlações favoráveis com características seminais, reprodutivas das fêmeas e de crescimento, a circunferência escrotal tornou-se o mecanismo de seleção genética mais utilizado com o objetivo de melhorar a eficiência reprodutiva dos rebanhos (PEREIRA, 2001). No entanto, MARTINEZ-VELAZQUEZ et al. (2003) constataram
uma correlação genética de -0,15 entre circunferência escrotal e idade a puberdade das fêmeas demonstrando, assim, que a resposta genética em características reprodutivas de fêmeas através da seleção dos pais não é eficaz.
Uma forma de auxiliar esta mensuração foi a inclusão de uma medida das fêmeas, a PPP14, ou seja, a probabilidade de prenhez de novilhas aos 14 meses. Este parâmetro prioriza a obtenção de fêmeas que se tornem prenhes precocemente, sem que isto diminua seu ganho de peso e, desta maneira, não prejudicando o desenvolvimento corporal das matrizes (ELER et al., 2000).
A PPP14 vem sendo utilizada na raça nelore e seus resultados têm mostrado que a obtenção de fêmeas precoces é possível e isto eleva a credibilidade nos trabalhos científicos por parte do produtor (MARSON et al., 2004). Segundo BERGMANN
(1999), as características indicadoras de precocidade sexual com uma variabilidade genética adequada, de fácil mensuração e baixo custo, correlacionadas geneticamente com a idade à puberdade, devem ser utilizadas, valorizadas e associadas às técnicas moleculares para o estudo dos genes.
2.1.3.3. Fatores Sazonais:
SCHILLO et al. (1992), os efeitos sazonais estão relacionados à temperatura ambiental,
à estação de nascimento, ao fotoperíodo, dentre outros.
Um dos fatores sazonais implicados na manifestação da puberdade em novilhas zebu é o clima tropical (CHENOWETH, 1994). Embora estas novilhas sejam termicamente adaptadas, mudanças na dinâmica folicular expressas como redução na taxa de crescimento folicular e aumento na duração do crescimento folicular estão associadas com aumento na temperatura corporal durante o verão (GAMA FILHO et al., 2002). Vacas precisam ser eficientemente climatizadas durante o verão para se ter
uma melhoria na fertilidade (WOLFENSON et al., 2000). Altas temperaturas
influenciam na idade à puberdade ou por afetar a circulação de LH ou os níveis de prolactina e a taxa de crescimento (NOGUEIRA, 2004).
Embora os bovinos não sejam considerados animais sazonais, um estudo com
Bos taurus demonstrou que a estação de nascimento e a manifestação da puberdade
influenciam a idade à primeira ovulação em novilhas (SCHILLO et al., 1983). Em
clima subtropical, novilhas com uma grande proporção de genes zebu mostraram um efeito direto do fotoperíodo sobre a regulação da atividade ovulatória, mas este efeito inexistiu em animais Bos taurus (MEZZADRA et al., 1993). PETERS et al. (1978)
relataram que a exposição de novilhas a 16h de luz por dia durante o inverno, ocasionou a elevação da taxa de crescimento com redução da idade à puberdade. Assim como HANSEN et al. (1983) constataram este mesmo efeito em novilhas nascidas durante a
primavera e início de verão, quando expostas a fotoperíodos de 18h de luz durante o outono e o inverno.
Algumas outras variáveis dependentes de influências de aspecto sazonal, como a nutrição, no que se refere à disponibilidade de alimentos, podem interagir com a estação do ano modificando o momento de ocorrência da puberdade (MARSON et al., 2004).
LESMEIESTER et al. (1973) relataram os efeitos expressivos da época de nascimento
tardiamente na estação de monta ou nem se quer esta ocorreu (ORMAZABAL et al.,
1996).
O efeito da estação de nascimento em novilhas nascidas na primavera ou no outono sobre a idade e peso médio à puberdade não foi observado por HONARAMOOZ et al. (1999). Nas pesquisas coordenadas por ALENCAR et al.
(1987) com relação a menor disponibilidade de forragens durante a recria de novilhas, foi constatado que esta restrição prejudicou o desenvolvimento dos animais até os 24 meses de idade e elevou a idade à puberdade.
2.1.3.4. Efeito da presença do macho:
Vários sistemas produtivos expõem muitas de suas novilhas pré-púberes à estação reprodutiva e, alguns antecipam ainda mais este manejo expondo as fêmeas recém desmamadas a touros, tentando tirar proveito do “efeito macho” (MARSON et al., 2004). Este efeito se traduz em estímulo sobre o eixo hipotalâmico-hipofisário da
fêmea, atuando sobre a secreção de gonadotrofinas com conseqüente manifestação da puberdade (PATTERSON et al., 1992).
Novilhas submetidas a este tipo de manejo atingiram a puberdade em idade mais precoce e com menos peso corporal, porém foi necessário um tempo mínimo e efetivo de 2 a 3 meses de exposição das novilhas aos touros para que houvessem efeitos significativos sobre a puberdade (SPIRE, 1997).
WEHRMAN et al. (1996) e ORMAZABAL et al. (1996) não detectaram efeitos
positivos com relação à exposição de touros a novilhas para a antecipação da puberdade. Deste modo, MARSON et al. (2004) consideram os dados disponíveis sobre
2.1.4. Estratégias para reduzir a idade à puberdade:
Alguns procedimentos farmacológicos podem ocasionar um decréscimo na idade a primeira concepção (NOGUEIRA, 2004). O tratamento com norgestomet e PMSG pode reduzir a idade a primeira concepção em novilhas Bos indicus
(NARASIMHA RAO et al., 1986). O tratamento com ivermectina também acelerou a
puberdade independente do ganho de peso em novilhas de corte (LARSON et al.,
1995). O tratamento mensal com bST, dos 7 aos 12 meses de idade, em novilhas nelore, alterou o diâmetro da fibra muscular, mas não afetou o ganho de peso ou a precocidade sexual (MOREIRA et al., 2000).
2.1.5. A puberdade em rebanhos mestiços (Bos indicus X Bos taurus) :
Zebuínos são conhecidos por serem menos férteis e por apresentarem uma produção leiteira inferior à dos taurinos de um modo geral, mas sua melhor adaptação a condições ambientais diversas faz deles os mais aptos reprodutivamente nos trópicos (NOGUEIRA, 2004). Os mestiços incorporam a adaptação ambiental dos zebuínos e a produtividade dos taurinos apresentando um alto grau de heterose (NEGUSSIE et al.,
1999).
Bovinos Bos indicus foram importados na América do Norte para cruzamentos
com o objetivo de explorar suas características adaptativas e, alguns destes cruzamentos, foram mantidos para produzir um novo rebanho mestiço (CARTWRIGHT, 1980). Um bom exemplo do resultado deste processo é a raça Brahman que, embora seja considerada Zebu, é uma mistura da Guzerá, Nelore e Gir com uma pequena contribuição de raças européias que foram usadas no processo de cruzamento contínuo (SANDERS, 1980).
O efeito aditivo dos cruzamentos pode ser visto na idade a primeira ovulação que diminui de 22 para 15 meses em bovinos mestiços Bos indicus X Bos taurus
genes de Bos taurus no DNA, mas é impossível dizer quando esta contribuição foi
introduzida (MEIRELLES et al., 1999).
Fêmeas mestiças 1/2 Europeu 1/2 Zebu mostram-se mais precoces e têm maior potencial de crescimento que animais 3/4 Zebu 1/4 Europeu (SILVA & PEREIRA, 1986). O aumento na proporção de genes Bos indicus no cruzamento foi acompanhado
por uma diminuição na produtividade em novilhas mestiças criadas em clima subtropical (ROCHA & LOBATO, 2002). Em um experimento conduzido na Argentina comparou-se gado puro com mestiço e foi observada puberdade antecipada nos mestiços (MEZZADRA et al., 1993).
Utilizando novilhas mestiças, GARCIA et al. (2002) constataram que o peso
corporal explicava a maior parte da variação associada ao inicio da puberdade seguida pela contribuição das concentrações de leptina circulante, às quais aumentam durante um período de 16 semanas antes da primeira ovulação.
Puberdade precoce em mestiços pode ser atribuída à heterose, mesmo aonde exista uma suplementação alimentar adequada pós-desmame (MEZZADRA et al.,
1993; MARSON et al., 2001). No entanto, a nutrição é um fator limitante porque
mestiços 5/8 Europeu 3/8 Zebu quando alimentados com forragens de baixa qualidade atingem a puberdade na mesma idade de zebuínos puros (ALENCAR et al., 1987).
Alguns cruzamentos (Red Angus e Simental X Nelore) apresentam maior eficiência de produção ao desmame que outros (Marchigiana ou Guzerá X Nelore) com o maior efeito aditivo ocorrendo quando genes de Bos indicus são combinados com os
de Bos taurus (PEROTTO et al., 2001). RESTLE et al. (1999) demonstraram uma
heterose de -12,8 para idade e peso à puberdade de mestiços Charoles X Nelore comparados a rebanhos puros.
2.2. Fisiologia Reprodutiva da Fêmea Bovina:
2.2.1. A regulação:
Dois sistemas regulatórios estão envolvidos no processo reprodutivo: o sistema endócrino e o sistema nervoso (PTASZYNSKA, 2003). Estes dois sistemas atuam de forma integrada e sinérgica, através de complexos mecanismos interligados do eixo hipotálamo-hipófise-gônadas, controlando assim as diversas funções reprodutivas (GRUNERT et al., 2005).
2.2.2. Órgãos e Sistemas Liberadores de Hormônios:
2.2.2.1. Hipotálamo:
Ocupa apenas uma porção cerebral, a porção anterior do diencéfalo (HAFEZ & HAFEZ, 2004; GRUNERT et al., 2005), formando conexões neurais com o lobo
posterior hipofisário, através do trato hipotálamo-hipofisário, e conexões vasculares com o lobo anterior da hipófise (HAFEZ & HAFEZ, 2004).
O sangue arterial chega a hipófise através das artérias hipofisárias inferior e superior, à qual forma alças capilares por onde o sangue flui para o sistema porta-hipotalâmico-hipofisário que começa e termina em capilares sem passagem pelo coração. Parte do fluxo venoso que sai da hipófise anterior forma um fluxo retrógrado, o qual expõe o hipotálamo a elevadas concentrações dos hormônios da hipófise anterior. Este fluxo possibilita a retro-alimentação negativa de regulação das funções hipotalâmicas e é conhecido como retro-alimentação de alça curta (HAFEZ & HAFEZ, 2004).
seretado de forma pulsátil e, desta forma, controla as secreções do FSH e do LH ao longo do ciclo estral (GRUNERT et al., 2005).
2.2.2.2.Hipófise:
Localiza-se na base cerebral e divide-se em três partes anatômicas distintas: lobos anterior, intermediário e posterior (HAFEZ & HAFEZ, 2004). Na hipófise anterior as células basófilas são responsáveis pela produção do FSH e do LH, os quais são conhecidos como gonadotrofinas hipofisárias (GRUNERT et al., 2005). Estas
gonadotrofinas estimulam a função gametocitogênica (crescimento, maturação e ovulação dos folículos) e estimulam a produção de estrógenos, andrógenos e progesterona pelas células das tecas foliculares, e inibina nas células da granulosa que atuará na regulação do FSH (DÖCKE, 1994).
2.2.2.3.Ovários:
Desempenham uma dupla função: a produção de células germinativas e a secreção de hormônios gonadais. As células da teca interna e as da granulosa do folículo ovulatório formam a fonte primária dos estrógenos circulantes e, após a ovulação, as células da granulosa e da teca são substituídas pelo corpo lúteo (CL), o qual secreta progesterona (HAFEZ & HAFEZ, 2004).
Os estrógenos promovem as modificações comportamentais durante o estro com a aceitação da monta pela fêmea e modificações em seu trato reprodutivo como, por exemplo, hipertrofia e hiperplasia uterina (GRUNERT et al., 2005).
2.2.2.4.Útero:
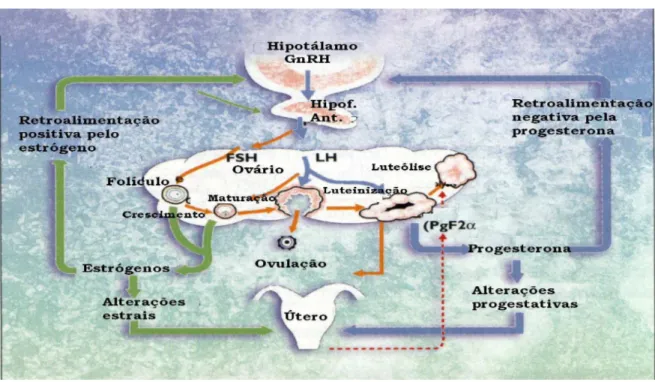
carúnculas. O endométrio varia em espessura e vascularização com as alterações hormonais ovarianas e com a prenhez, e participa efetivamente no controle da atividade do CL através da produção e liberação de PGF2α (prostaglandina F2α), um luteolítico natural que é liberado cerca de 14 dias após a ovulação no útero não gestante(Fig.01) (REECE, 1996).
Fig 1- Esquema das trocas hormonais no eixo Hipotálamo-Hipófise-Ovários-Útero.
2.3. Crescimento Folicular e Ovulação:
em baixa produção de andrógenos e, desta forma, baixa produção de estradiol, o que leva à atresia do folículo e degeneração do oócito (FORTUNE, 1994; FIENI et al.,
1995; MEREDITH, 1995; GINTHER et al., 2000).
O ovário possui milhares de folículos primordiais em estado inativo ao longo da vida e, alguns destes, por mecanismos desconhecidos, são recrutados para iniciar o desenvolvimento. Grupos de folículos de 5-8 mm emergem sobre a superfície ovariana e podem ser observados através de ultra-sonografia. A emergência de um grupo de folículos é precedida por uma elevação dos níveis de FSH. A partir deste grupo de folículos emergentes, um folículo é selecionado para continuar o desenvolvimento e tornar-se dominante (FD – folículo dominante). Esta seleção do FD ocorre em associação a um decréscimo nos níveis de FSH circulantes (FORTUNE, 1994; GINTHER et al., 1999).
O FD suprime o crescimento dos outros folículos, conhecidos como folículos subordinados (FS), e continua a se desenvolver mesmo diante da diminuição nas concentrações de FSH devido ao aumento na sua sensibilidade ao FSH, através da produção de fatores de crescimento locais, principalmente o IGF-I (fator de crescimento da insulina) (FORTUNE et al., 2001; RIVERA & FORTUNE, 2001).
O destino do FD é determinado pela freqüência do pulso de LH à qual o mesmo é exposto, ou seja, haverá ovulação em resposta a um pulso de LH a cada hora e, haverá atresia folicular quando a freqüência do pulso de LH for de um pulso a cada 3-8h (FIENI et al., 1995; GINTHER et al., 1999; FORTUNE et al., 2001).
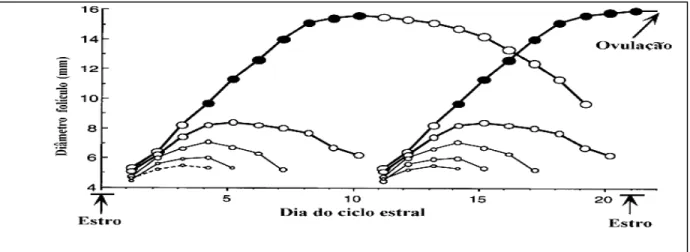
Este crescimento dos folículos ovarianos em bovinos é denominado de ondas de crescimento folicular e ocorre antes da puberdade, durante o ciclo estral, durante a prenhez e no pós-parto (STEVENSON & PURSLEY, 1994; BEAM & BUTLER, 1997). Com relação à dinâmica folicular de bovinos de raças européias, estes podem apresentar duas, três ou, até mesmo, quatro ondas de crescimento folicular (SIROIS & FORTUNE, 1988; GINTHER et al., 1989; MURPHY et al., 1990).
que 7mm de diâmetro na superfície, cem dias antes da primeira ovulação (ROBERSON, 1990).
Em um ciclo estral normal de uma fêmea bovina, um grupo de 10-50 folículos com 2-3mm emergem entre os dias 1-3 após o estro. Posteriormente, alguns crescem para 4-6mm e 2-5 folículos maiores continuam o crescimento enquanto os outros regridem. Daí, pelo menos um continua o crescimento e torna-se FD, o qual apresenta crescimento mais rápido que os outros, os FS (BODENSTEINER et al., 1996).
O tamanho do folículo ovulatório (FO) é altamente variável entre as fêmeas bovinas, ou seja, em novilhas da raça holandesa foram observados FO de 14,8 ± 0,2mm (SARTORI et al., 2000). GINTHER et al. (1989) observaram FO com tamanhos de
16,5 ± 0,4mm em novilhas com duas ondas foliculares, mas menores (13,9 ± 0,4mm) em novilhas com três ondas foliculares.
A emergência da onda folicular é o período em que os primeiros folículos atingiram tamanhos PP R TXH DFRQWHFH QR SLFR GR )6+ $SyV D HPHUJência, os folículos continuam a crescer e a concentração de FSH no sangue diminui até o momento da divergência folicular (WILTBANK et al., 2002). Esta foi definida como o
início da maior diferença nas taxas de crescimento entre o maior folículo, o FD, e o segundo maior folículo, o maior FS, quando o último atingiu o diâmetro máximo (GINTHER et al., 1996).
O FD desenvolve-se em três fases: crescimento, estática e regressão, aonde na primeira onda de crescimento folicular a fase de crescimento vai da emergência ao oitavo dia do ciclo estral; a fase estática do oitavo ao décimo e a fase de regressão ocorre após o décimo dia. A partir do décimo dia do ciclo estral, inicia-se a segunda onda de crescimento folicular e o reinicio do processo com o FD regredindo, no caso de três ondas, ou se tornando FO, no caso de duas ondas apenas (SILCOX et al., 1993).
A ocorrência de duas ou três ondas de crescimento folicular é determinada pela taxa de crescimento folicular e pela duração da fase lútea (KASTELIC et al., 1990). Em
progesterona retarda a secreção de PGF2α no útero até 16-18 dias após o estro, o que resultará na duração normal do ciclo estral (WILTBANK et al., 2002).
GINTHER et al. (1989) constataram que o intervalo médio entre as ovulações é
menor (± 20 dias) para ciclo com duas ondas e maior (± 23 dias) para ciclos com três ondas, relacionando uma terceira onda com uma fase luteal mais longa. PURSLEY et al. (1995) concluíram que animais com três ondas de crescimento folicular têm menor
resposta à primeira injeção de GnRH do protocolo “OVSYNCH”, o que reduz a taxa de sincronização, já que o intervalo entre ondas nesses animais é mais curto e a chance de se ter folículos capazes de ovular no momento da aplicação do GnRH é menor do que em animais com duas ondas (Fig.02).
2.4. Hormônios relacionados com a onda de crescimento folicular:
2.4.1. Hormônio liberador de gonadotrofinas (GnRH):
A descoberta dos pulsos de GnRH na circulação portal e a demonstração de que essa pulsatilidade era fundamental na manutenção da síntese e secreção das gonadotrofinas foram fundamentais para o entendimento da sua função reprodutiva (KNOBIL, 1974; 1980).
Diversos experimentos que avaliaram diretamente a liberação do GnRH em fêmeas bovinas, através de amostragem de fluido cerebrospinal, comprovaram a existência de uma associação dosagem resposta temporal muito próxima entre a secreção do GnRH a partir do hipotálamo e a secreção do LH a partir da hipófise (GAZAL et al., 1998;
YOSHIOKA et al., 2001).
A regulação tanto da freqüência como da amplitude dos pulsos de GnRH é fundamental para a transmissão do sinal correto para a secreção e a síntese de gonadotrofinas na hipófise. Em geral, pulsos de GnRH de maior freqüência e baixa amplitude favorecem a secreção de LH, enquanto os de baixa freqüência e alta amplitude favorecem a secreção do FSH (KAISER et al., 1995; VIZCARRA et al.,
1997;1999).
LINCOLN et al. (1984) propuseram um sistema de controle responsável pela
regulação dos pulsos de GnRH, o qual foi denominado de “gerador de pulsos do GnRH”. Vários fatores interferem na atividade deste gerador, porém os mais importantes em termos de ciclo estral são os esteróides gonadais, estrogênio e progesterona (WILTBANK et al., 2002).
GOODMAN & KARSCH (1980) descobriram que quando a progesterona é o hormônio predominante na circulação (fase luteal do CE), a secreção do LH se caracterizava por pulsos de freqüência muito baixa e amplitude alta. Quando as concentrações de estradiol na fase folicular foram mantidas, os pulsos do LH ocorreram em uma freqüência maior e amplitude menor. Posteriormente, foi confirmada que a secreção do GnRH corresponde ao padrão previsto pelos pulsos de LH (MOENTER et al., 1991).
Esses efeitos intensos da progesterona e do estradiol na pulsatilidade do GnRH (e do LH) são fundamentais para o desenvolvimento dos folículos, pois somente após uma queda no nível da progesterona é que o gerador de pulsos do GnRH irá produzir os pulsos de alta freqüência do LH necessários para estimular a produção do estradiol que resultará na ovulação do FD (WILTBANK et al., 2002).
As concentrações de FSH no sangue têm uma relação funcional importante com o padrão de crescimento folicular (fig.03), já que as ondas foliculares são sempre precedidas por aumento nos níveis de FSH (ADAMS et al., 1992). Próximo ao estro
ocorre dois picos de FSH difíceis de serem identificados por se passarem quase ao mesmo tempo. O primeiro pico corresponde ao pico de GnRH/LH que induz a ovulação e o segundo ocorre próximo a ovulação, sendo associado à emergência da primeira onda folicular, à qual é caracterizada como o período em que os primeiros folículos atingiram tamanho PP$HPHUJência da onda se dá no pico de FSH e, após esta, os folículos continuam o crescimento e o nível de FSH sangüíneo diminui até a divergência folicular (WILTBANK et al., 2002).
Um ponto relevante em todas as fases da reprodução de bovinos é a interação entre a secreção do FSH na hipófise e dos inibidores do mesmo nos ovários. Esse fenômeno foi denominado ligação bidirecional funcional entre o FSH e os folículos (GINTHER et al., 2000). GIBBONS et al. (1997) constataram que na ausência de
folículos após a ovariectomia em bovinos ocorre um rápido aumento do FSH no sangue que atinge o pico dentro de 24h com posterior secreção máxima contínua. Além disso, estes mesmos autores verificaram que na aspiração de todos os folículos com mais de 3mm houve um aumento semelhante nos níveis de FSH, indicando que a atividade inibidora deste hormônio deriva de folículos com mais de 3mm de diâmetro.
Os dois principais inibidores do FSH secretados pelo folículo são a inibina e o estradiol, sendo que a inibina parece ser secretada pelos folículos de todos os tamanhos e o estradiol tem seus níveis aumentados na corrente sangüínea somente após a seleção de um FD depois da divergência (KULICK et al., 1999).
2.4.3. Hormônio luteinizante (LH):
Tanto o crescimento folicular quanto a produção de estradiol nos folículos após a divergência parecem ser controlados pelos pulsos de LH (WILTBANK et al., 2002).
GONG et al. (1995) verificaram que os folículos não conseguiam atingir mais que 9mm
vacas tratadas com um antagonista de GnRH para redução dos pulsos de LH, observou-se que nas vacas tratadas nos dias 2-7 após o estro, durante a primeira onda folicular, o folículo maior cresceu até um diâmetro médio de 8mm e persistiu por apenas 6,3 dias em comparação ao diâmetro máximo de 11,8mm e persistência de 10,4 dias nas novilhas controle. Ainda foi observada uma redução na concentração de estradiol sangüínea após esta inibição nos pulsos de LH (FIKE et al., 1997).
GINTHER et al. (2001) em um outro estudo, constataram que a inibição dos
níveis de LH através de tratamento com porgesterona não alterou o momento nem as características de diâmetro na seleção folicular, mas a taxa de crescimento do FD em desenvolvimento caiu quando ele atingiu 10mm de diâmetro. Desta forma, o crescimento folicular após a divergência parece necessitar de pulsos de LH, sendo que o tamanho máximo do folículo pode ser reduzido quando são diminuídos os pulsos de LH (WILTBANK et al., 2002).
Foi observado que uma redução nas concentrações de progesterona sangüínea aumenta a quantidade dos pulsos de LH, prolonga o crescimento e aumenta o diâmetro máximo do FD (LUCY et al., 1992; BERGFELD et al., 1996; NE-161, 1996). Esse FD
persistente também pode ser induzido através de tratamento com pequenos pulsos de LH exógeno (TAFT et al., 1996). De grande importância fisiológica é o achado de que
um aumento transitório das concentrações médias de LH sangüíneas inclui o momento esperado da divergência folicular (KULICK et al., 1999), mesmo não sendo este
necessário para a divergência folicular (GINTHER et al., 2001).
2.4.4. Estradiol e Inibina:
O FD secreta mais que 80% do estradiol e também é responsável por 55% da inibina liberada na circulação sangüínea (CAMPBELL et al., 1991). BODESTEINER et al. (1996) verificaram que as concentrações de estradiol foram 10 vezes maiores no
maior folículo da onda (8,5mm de diâmetro) quando comparado às do segundo maior folículo(7,3mm de diâmetro). Além disso, KANEKO et al. (1992) constataram que a
Níveis máximos de FSH são observados na emergência de uma nova onda folicular (fig.03), provavelmente devido às baixas concentrações de inibina e estradiol no sangue. À medida que a onda folicular avança, os folículos maiores parecem aumentar a atividade inibitória do FSH, provavelmente a inibina, reduzindo o FSH no sangue. A concentração de estradiol no sangue aumenta de 0,2 para 1pg/mL próximo ao momento da seleção do folículo, o que causa a queda final nos níveis de FSH sangüíneo nesse momento (KULICK et al., 1999).
Em dois experimentos aonde novilhas foram tratadas com valerato de estradiol e HVWUDGLRORFUHVFLPHQWRIROLFXODUIRLVXSULPLGRHRFRUULDDHPHUJência de uma nova onda de crescimento folicular (BO et al., 1993; 1994). Porém, o estradiol sozinho é um
inibidor fraco da secreção do FSH, mas atua de forma sinérgica com a inibina para inibir intensamente essa secreção (WILTBANK et al., 2002).
KANEKO et al. (1995) verificaram uma relação inversa entre os níveis séricos
de inibina e FSH. A imunização contra a inibina resultou em aumento no número de folículos e na taxa de ovulação em bovinos, pelo aumento nos níveis de FSH sangüíneo (KANEKO et al., 1993; MORRIS et al., 1993; GLENCROSS et al., 1994). Tanto a
inibina quanto o estradiol são repressores potentes da síntese e da secreção do FSH no interior da hipófise (WILTBANK & HAUGHIAN, 2003).
2.5. Inseminação artificial em tempo fixo (IATF):
Com a atual situação econômica da pecuária mundial, exigi-se cada vez mais dos produtores a máxima eficiência produtiva para garantia de um retorno econômico satisfatório. Assim, elevados índices produtivos, associados à alta eficiência reprodutiva, devem ser objetivos que norteiam técnicos e produtores. Nesta concepção, a melhoria da eficiência reprodutiva é um dos principais fatores que contribuem para a melhora da performance produtiva e da lucratividade das criações bovinas (BARUSELLI et al., 2004).
A inseminação artificial (IA) consagrou-se mundialmente e provou ser uma técnica viável para acelerar o ganho genético e o retorno econômico da pecuária. Programas que empregam a IA em tempo fixo (IATF), sem a necessidade de detecção do estro, auxiliam de forma direta no emprego desta biotecnologia. Dentre os programas existentes, destacam-se os protocolos que usam GnRH, LH e hCG e prostaglandinas e/ou progesterona e estrógenos, todos sempre visando a IATF (BARUSELLI et al., 2004).
O Brasil possui mais de 73,5 milhões de fêmeas bovinas em idade reprodutiva criadas, na maioria, a pasto, aonde ocorrem comprometimentos na taxa de detecção de cio e na eficiência dos programas de IA. A perda de um cio retarda em mais ou menos 21 dias a IA e, aumenta a idade ao primeiro parto, o intervalo entre partos, diminui o número de bezerros nascidos e, conseqüentemente, a produção de leite (BARUSELLI et al., 2004).
Segundo DAY (2005) as novilhas são ótimas candidatas para a IA, tanto por razões práticas quanto por razões fisiológicas, ou seja, como não pariram anteriormente, não apresentam desafios como o anestro pós-parto, problemas induzidos por distocia e remoção de bezerro. Geralmente, as novilhas foram submetidas a um manejo que não apresenta grandes variações na condição corporal, facilitando o início de um programa de IA. Além disso, do ponto de vista genético, as novilhas representam as melhores fêmeas do rebanho, as mais avançadas geneticamente, como resultado de decisões anteriores relativas à reprodução.
Um fator limitante para o sucesso de programas de IA em novilhas é o momento da puberdade em relação ao inicio da estação de monta. BYERLEY et al. (1987)
demonstraram que há menor probabilidade das novilhas emprenharem em seu primeiro ciclo estral e que a fertilidade aumenta até o terceiro ciclo. Os produtores, muitas vezes, entendem que suas novilhas estão púberes antes do início da estação de IA devido à ocorrência do estro sem ovulação, ou seja, o estro não puberal (NELSEN et al., 1985;
RUTTER & RANDEL, 1986).
Segundo WEHRMAN et al. (1996) e ANDERSON & DAY (1998), o início
precoce do ciclo estral ocorre em um número significativo de novilhas. Porém, LUCY
et al. (2001), trabalhando com 724 novilhas em 5 locais nos EUA constataram que 43%
destas ainda se encontravam pré-púberes 7 dias antes da estação de IA com uma variação de 6 a 81% entre os rebanhos. Mas esta proporção de novilhas pré-púberes no início da primeira estação de monta pode ser amenizada através de manejo nutricional adequado, seleção de novilhas e uso de protocolos de sincronização (KINDER et al.,
1995). Desta maneira, os programas de sincronização devem ser ajustados para utilização tanto em animais cíclicos quanto em acíclicos (DAY, 2005).
2.6. Os protocolos de IATF:
2.6.1. Sincronização usando prostaglandina F2α (PGF2α):
A maneira mais tradicional de sincronização de estro tem sido o uso da PGF2α, que foi descoberta na década de 70 (MCCRACKEN, 1972) e que regula o ciclo estral da fêmea por provocar a regressão do CL. Uma injeção de prostaglandina mimetiza a liberação natural de PGF2α do útero para promover a regressão do CL. A regressão coordenada do CL ocasionará uma diminuição na progesterona, resultando numa apresentação síncrona de estro 2 a 5 dias depois em novilhas que estejam ciclando e que apresentem um CL responsivo a PGF2α (RATHBONE et al., 2001; DAY, 2005). Ainda foi determinado que a PGF2α não induz de forma constante a regressão lútea entre os dias 1-5 do CE, aumentando sua eficácia entre os dias 6-17 (LAUDERDALE et al., 1974).
KASTELIC & MAPLETOFT (1998) relataram que, quando o tratamento com
PGF2α é realizado no dia 5 do CE, momento em que o FD da primeira onda está em fase de crescimento, o intervalo aplicação-ovulação é de 3 dias. Se o tratamento for realizado no dia 12 do CE, quando o FD da segunda onda encontra-se no início da fase de crescimento, a ovulação ocorre 4,5 dias após a aplicação. Bovinos tratados no dia 8 do CE apresentam ovulação do FD da primeira onda 2 dias após a aplicação da PGF2α, mas pode ocorrer a ovulação do segundo FD 6 dias após o tratamento.
Segundo DAY (2005) a responsividade do CL à PGF2α foi menor em fêmeas tratadas entre os dias 5-9 (67%), intermediária para as tratadas entre os dias 9-12 (77%) e maior em fêmeas tratadas depois do dia 12 (91%). Após o dia 17 do CE, a PGF2α parece não influenciar o momento do estro, mas como estas fêmeas irão apresentar estro espontaneamente nos próximos 2-6 dias, estarão automaticamente sincronizadas.
As novilhas Bos indicus parecem ser menos responsivas a este tratamento,
independente do momento do CE em que a administração foi realizada (ORIHUELA et al., 1983; LANDIVAR et al., 1985; BRIDGES et al., 2000). Já bovinos de raças
JACKSON et al. (1979) desenvolveram protocolos com duas aplicações
seqüenciais, intervaladas de 11-14 dias, objetivando a melhoria da sincronização de cio com PGF2α. BRIDGES et al. (2000) observaram uma melhor resposta estral e taxa de prenhez em novilhas de sobreano mestiças quando a dose de PGF2α foi dividida e administrada com um intervalo de 24h. Isto se deveu a um aumento na regressão do CL por causa da administração fracionada de PGF2α.
Outro sistema proposto para melhorar a eficiência da PGF2α é a observação de
cio por 5 dias, com a aplicação da PGF2α no dia 6, para evitar tratar animais com CL não responsivo ao tratamento. Deste modo, é de fundamental importância a presença do CL funcional em fase específica do CE, ou seja, no diestro. Além disso, o estro e a ovulação não são precisamente sincronizados devido à variação do estágio de desenvolvimento folicular no momento da regressão do CL, já que alguns animais apresentam atraso no desenvolvimento e na maturação do FO. Logo, faz-se necessária a
detecção do cio após a administração de PGF2α para a realização da IA e para a obtenção de melhores taxas de concepção (STEVENSON et al., 1999).
A alta variabilidade das respostas ao tratamento com PGF2α e a característica de animais criados a pasto em condições tropicais apresentarem alta incidência de anestro, com ausência de CL, tem comprometido a eficiência deste tratamento. Deste modo, torna-se imprescindível o uso de meios que controlem o desenvolvimento luteínico e folicular para sincronizar o crescimento dos folículos e a sincronização da ovulação, permitindo o emprego da IATF sem observação de cio (BO et al., 2003).
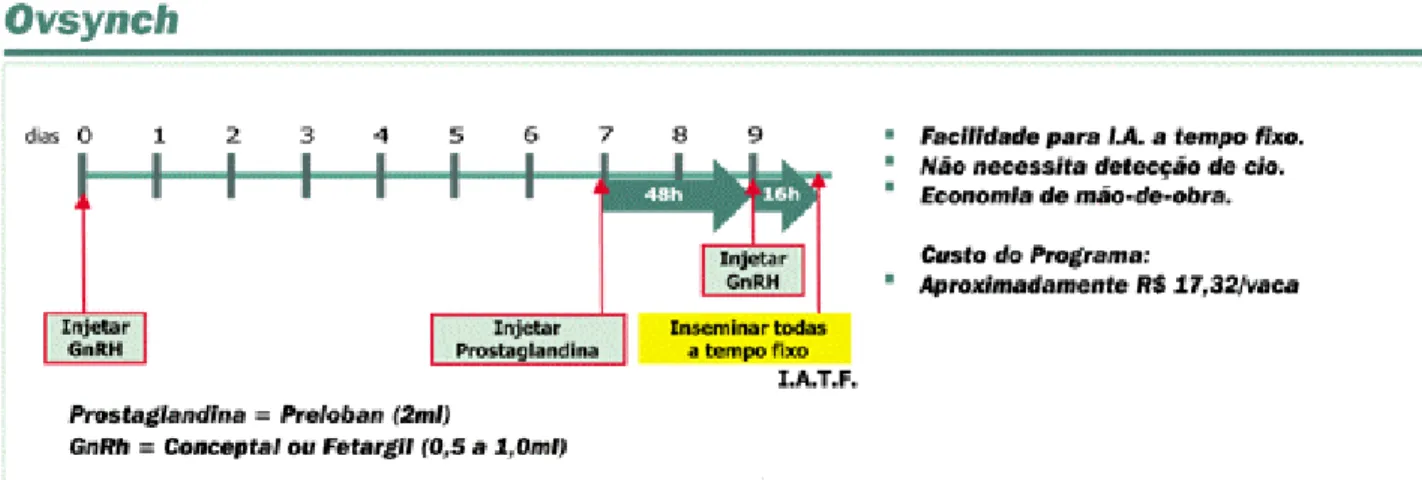
2.6.2. Sincronização usando GnRH- PGF2α (“OVSYNCH”):
secreção de estradiol e outros fatores responsáveis pelo bloqueio do FSH na hipófise (WILTBANK & HAUGHIAN, 2003).
Muitas pesquisas foram direcionadas para o desenvolvimento de sistemas que usem uma combinação de GnRH e PGF2α para sincronizar o estro em bovinos. Estes sistemas incluem a administração de GnRH ou um análogo, seguido 7 dias depois de PGF2α. A administração do GnRH tem por objetivo induzir a emergência síncrona de uma nova onda de crescimento folicular 1 a 2 dias depois, e a regressão do CL no dia 7 permite o início do proestro. Se a injeção inicial de GnRH for eficaz, os folículos estão em um estágio de desenvolvimento padrão e o FD maduro no momento da regressão lútea, aumentando-se a precisão do estro sincronizado (DAY, 2005).
Equipes americanas das universidades da Flórida (WOLFENSON et al., 1994)
e de Wisconsin (PURSLEY et al., 1995) desenvolveram o protocolo de sincronização
da ovulação “OVSYNCH”, o qual permite a sincronização da ovulação sem a
necessidade de observação de cio. Este tratamento consiste em uma injeção de PGF2α em fêmeas tratadas previamente (6-7 dias) com GnRH. Este induz uma descarga hipofisária de LH, seguida de ovulação ou regressão (atresia) de um possível FD, resultando na emergência de uma nova onda folicular nos dois ou três dias seguintes a injeção de GnRH (WOLFENSON et al., 1994; PURSLEY et al., 1995). A sincronia do
cio e da ovulação pode ser aumentada (mais de 90%) administrando-se uma segunda injeção de GnRH, 36-48h após a PGF2α. A fertilidade dos animais inseminados em tempo pré-fixado (16-24h após a segunda injeção de GnRH) assemelha-se à verificada em animais inseminados no cio observado (PURSLEY et al., 1995; 1997).
PURSLEY et al., 1995 relataram que o início da onda, após a ovulação do FD,
foi de 2,10±0,31 dias para vacas e de 1,50±0,47 dias para novilhas. BARUSELLI et al.
(2000) verificaram que bovinos tratados com este protocolo apresentaram maior taxa de ovulação (74,3%) que aqueles tratados com uma única dose de PGF2α (58,8%). A taxas de concepção de fêmeas bovinas de leite sincronizadas pelo “OVSYNCH” e inseminadas em tempo fixo variam de 26-55% (PURSLEY et al., 1995; 1997;
VASCONCELOS et al., 1999).
novilhas do que em vacas lactantes. Esta menor duração das ondas foliculares aumenta as chances de administração da primeira injeção de GnRH na ausência de um FD e/ou renovação de uma onda folicular antes que a ovulação possa ser induzida com a segunda injeção (PURSLEY et al., 1997).
Em experimento conduzido por PURSLEY et al., 1997 utilizando o protocolo
“OVSYNCH” em novilhas leiteiras, as taxas de prenhez (35%) foram inferiores às do tratamento controle com PGF2α (74,4%). Em outro experimento conduzido por STEVENSON et al. (2000) também com novilhas leiteiras, este protocolo mostrou-se
mais efetivo com taxa de concepção e de prenhez de 42%, mas ainda assim não se mostrando indicado para esta classe de fêmeas. RIBEIRO et al. (2001) trabalhando com
novilhas de duas raças de corte obteve taxas de concepção ainda menores: de 6,2% para a raça Santa Gertrudes e de 35,7% para a raça Pitangueiras.
Em Zebuínos, a resposta ao “OVSYNCH” foi avaliada em vacas lactantes (60-90 dias pós-parto) e em vacas não lactantes, aonde foram obtidas taxas de prenhez de 47% para as lactantes e 45% para as não lactantes (BARROS, 2000). Quando usado em vacas em anestro este protocolo revelou taxas bem mais baixas (14,9%), assim como em vacas de corte lactantes (15%). Indicando, desta forma, que o protocolo “OVSYNCH” não deve ser usado em animais em anestro,devendo ser utilizado somente em vacas que se apresentem ciclando (BARUSELLI et al., 2002).
MONTEZUMA JÚNIOR et al. (2004) obtiveram taxa de concepção mais
elevada (57%) utilizando este protocolo em vacas Girolandas no estado do Ceará que se apresentavam em anestro pós-parto. OLIVEIRA et al. (2003) obtiveram taxas de
concepção variando de 34,6-59% em primeira e segunda IAs utilizando animais em anestro pós-parto da raça Girolanda no estado do Ceará, com graus de sangue semelhantes aos dos animais experimentais utilizados por MONTEZUMA JÚNIOR et al. (2004).
2.6.3. Sincronização utilizando progestágenos (CIDR-B):
provavelmente aos períodos longos dos tratamentos: de 14 a 21 dias (WILTBANK et al., 1965). Atualmente, sabe-se que períodos prolongados de utilização de
progestágenos levam a formação de folículos persistentes. Além disso, os oócitos oriundos destas ovulações não se desenvolviam plenamente em razão da ativação precoce da meiose, à qual em condições normais ocorre pelas liberações pré-ovulatórias de LH (FORTUNE & RIVERA, 1999).
Talvez um dos principais conceitos surgidos recentemente tenha sido sobre os incrementos realizados para melhorar a taxa de concepção, que podem ser obtidos com a atresia do FD, no início do tratamento com progesteronas, impedindo-se, assim, a formação de folículos persistentes e permitindo o desenvolvimento de uma nova onda folicular que resultaria em um FD ovulatório pronto para a fertilização (MADUREIRA, 2000).
Dentre as formas de controle do crescimento folicular, o tratamento com progesterona e estradiol tem se mostrado eficaz na promoção da atresia dos folículos em desenvolvimento e na emergência de uma nova onda folicular 3 a 5 dias depois (BO
et al., 1994; 1995). Desta maneira, o benzoato de estradiol no início do tratamento com
progesterona promove a atresia dos FDs presentes e a emergência de uma nova onda folicular (BRAGANÇA et al., 2004). Ressaltando que pode ser também associado um
agente luteolítico (PGF2α), objetivando um melhor controle do estro e da ovulação (MARTINEZ et al., 2000).
Até 2002, o acetato de melengestrol (MGA) era a única progestina aprovada pelo FDA (“Food and Drug Administration”) para uso na sincronização do estro. Neste mesmo ano, uma nova progestina foi aprovada pelo FDA, à qual trata-se de um dispositivo intravaginal de liberação de progesterona, o “CIDR” (“Controlled Internal Releasing Device”) (LAMB, 2003).
O suporte principal do “CIDR-B” é de nylon coberto com silicone impregnado com progesterona (1,9g). Este suporte, fixado adequadamente ao aplicador, é liberado na região anterior da vagina. Após a inserção, as concentrações de progesterona sobem rápido, atingindo o máximo em uma hora, mantendo-se constantes nos 7-8 dias em que o dispositivo permanece na vagina e, sendo eliminados rapidamente após a retirada, o que ocasionará um aumento na secreção pulsátil de LH resultando, dentro de 1-3 dias, em cio e ovulação do FD presente (LAMB, 2003).
A ponta do dispositivo fica visível e aderida à vulva, sendo que, a possível perda do mesmo pode ser atribuída a uma aplicação errônea na porção vaginal e /ou à remoção pelos próprios animais do rebanho, embora esta perda seja insignificante (2%). Em alguns casos, observa-se irritação vaginal, com ocorrência de muco claro, opaco ou amarelo quando o “CIDR-B” é retirado, sendo os mesmos considerados normais e não afetando a eficácia do dispositivo (LAMB, 2003).
A aplicação de PGF2α e seus análogos sintéticos 2 dias antes ou no momento de retirada do “CIDR-B” tem sido recomendada para assegurar a luteólise, uma vez que o benzoato de estradiol não se mostrou efetivo na indução da luteólise (DISKIN et al.,
2002). Nos 7-8 dias em que o “CIDR-B” permanece na vagina, a propagação da P4 não afeta a luteólise espontânea, ou seja, supondo-se que todas as novilhas e vacas têm CE de 21 dias, haverá duas populações de fêmeas após 6 dias de tratamento com “CIDR-B”: fêmeas sem e com CL mais de 6 dias após a ovulação. Deste modo, todas as fêmeas apresentariam CL potencialmente responsivo à PGF2α (LAMB, 2003).
Embora a maioria das pesquisas indique que apenas cerca de 90% dos CL em novilhas e vacas com mais de 6 dias após ovulação regridem com uma injeção de PGF2α, somente cerca de 60% das fêmeas apresentam CL no momento da aplicação da mesma, supondo-se que a regressão espontânea começa cerca de 18 dias após a ovulação. Por conseguinte, cerca de 95% das fêmeas tratadas com “CIDR-B” / PGF2α são sincronizadas para exibir estro dentro de poucos dias após a retirada do dispositivo. Contudo, mais de 95% das fêmeas tratadas irão exibir estro se o mesmo for monitorado por 5 dias após a retirada do “CIDR-B” (LAMB, 2003).