AN TON I N A M ARI A DE M ELO DOS SAN TOS
AO LARGO DA COSTA PORTUGUESA
AO LARGO DA COSTA PORTUGUESA
LI SBOA
1 9 9 9
LARVAS DE CRUSTÁCEOS DECÁPODES AO LARGO DA COSTA PORTUGUESA
Agora que necessito de agradecer publicam ente a todos os que, de um a m aneira ou de outra, m e aj udaram , faltam - m e as palavras. Antes de o fazer, quero assim , explicar que a m inha falta de eloquência não tem a ver com a qualidade das m últiplas aj udas ao longo deste trabalho, m as sim , com a m inha inaptidão para o exprim ir.
Com eço por prestar o m eu grande reconhecim ento ao I PI MAR ( I nstituto de I nvestigação das Pescas e do Mar) , na pessoa dos seus antigos Presidentes, Dr. Carlos Reis e Dra. Carm en Lim a, e do presidente actual Dr. Marcelo Vasconcelos, durante a duração do trabalho, e a todo o pessoal que aí trabalha. Enquanto instituição de acolhim ent o, eu não poderia ter desej ado m elhor, tanto em am biente de trabalho, com o em disponibilidade e facilidades técnicas postas à disposição. No entanto, de todo o pessoal que trabalha ou trabalhou no I PI MAR, gostaria de poder realçar alguns para os quais a m inha dívida de gratidão é m aior.
Com eço por agradecer à Prof. Doutora Maria Helena Barahona Fernandes ( Faculdade de Ciências de Lisboa) a sua confiança, entusiasm o, apoio e com preensão, desde o início.
Tam bém um agradecim ento m uito especial à Dra. Aura Cascalho, que foi quem de início m e apoiou, incent ivou e resolveu t odos os m eus problem as burocráticos no I PI MAR. Tam bém a ela devo os m eus conhecim entos da biologia, ecologia e sistem ática dos adultos, pois que não se podem estudar os “ filhos” sem se conhecerem os seus fam iliares m ais próxim os.
Seguidam ente, agradeço m uito especialm ente à Dra. Maria Em ília Cunha, que foi quem m e m ostrou a im portância do estudo das larvas de crustáceos decápodes e que ao longo do trabalho não teve receio de com igo com partilhar da sua enorm e experiência em estudos de plâncton.
À Téc. Prof. Principal Maria de Fátim a Quintela quero agradecer todo o apoio logíst ico no laborat ório de Zooplânct on onde passam os horas infindáveis, eu identificando as m inhas larvas e ela os copépodes.
À Doutora Teresa Monteiro agradeço o seu interesse neste trabalho, as suas im port ant es sugest ões e a disponibilidade para ler t odo o trabalho m inuciosam ent e.
Às Dras. Teresa Moita e I vone Figueiredo tenho a agradecer o apoio m oral e am izade nas inúm eras e frutuosas discussões sobre o plâncton, os crustáceos e a sua distribuição. Aos Drs. João Pereira e I sabel Meneses agradeço pelas m últ iplas aj udas de inform ática sem as quais não m e teria sido possível fazer o “ scanning” dos cerca de 400 desenhos e que perm itiu a sua introdução no texto.
A todo o pessoal antigo e actual da Divisão de Docum entação e Apoio ao Utente do I PI MAR, especialm ente à Dra. Maria de Lourdes Monteiro, Dra. Lídia Nunes, Fernando Sousa, Alice Meca Bom bas, Luísa Correia e Erm elinda Martins, o m eu m uito obrigada.
O m eu obrigada a todo o pessoal Técnico dos Laboratórios de Fitoplâncton, Zooplâncton e I ctioplâncton do Departam ento de Am biente Marinho, pelas facilidades que m e concederam em apoio logístico.
Pelo apoio e resolução de vários problem as burocráticos aos Directores dos Departam ento de Am biente Marinho, Eng. Joaquim Pissarra e Departam ento de Recursos Marinhos, Dra. Graça Pestana, assim com o às funcionárias que prestam apoio às Secretarias dos Departam entos, Adélia Mart ins, j á aposent ada, Manuela Cordeiro e Am élia Nait a o m eu m uito obrigada.
Finalm ente, m erecem especial referência a Dra. Maria José de Figueiredo pelo int eresse que m ost rou pelo t rabalho, assim com o, pelas interessantes discussões sobre a biologia e ecologia do lagostim , a Doutora Maria de Lourdes Marecos, por m e ter em prestado o seu com putador onde fiz o tratam ento dos dados e a toda a equipa do Sector de Oceanografia Física e Teledetecção pelos ensinam entos sobre a circulação e as m assas de água na costa portuguesa.
Ao Prof. Doutor José Paula ( Laboratório Marít im o da Guia da Faculdade de Ciências de Lisboa) , o m eu profundo agradecim ento pela sua aj uda inicial no arranque do trabalho com bibliografia, explicações e m esm o a cedência de alguns estádios larvares de crustáceos decápodes por ele cult ivados em condições de laboratório e que m e perm it iram a sua com paração com os provenientes do plânct on.
Ao Dout or Alist air Lindley ( Natural Environm ent Research Council do Plym outh Marine Laboratory, Grã- Bretanha) , devo o seu interesse e estím ulo por este trabalho que se m anifestou nas sugest ões e correcções do m esm o.
Tam bém agradeço ao Doutor Marco Mura ( Universit à Cagliari, I t ália) e ao Dr. José António Cuesta ( Universidad de Sevilla, Espanha) , a am izade e a cedência de exem plares de estádios larvares, cultivados por eles em condições de laboratório e que m e proporcionaram a sua com paração com as larvas provenientes do plânct on.
À Prof. Doutora Maria Thessalou- Legaki ( Universidade de Atenas, Grécia) , agradeço as úteis discussões, ensinam entos sobre os Thalassinidea, leitura crítica da parte respeitante a este grupo e a ainda a sua am izade.
I nstituto de Ciencias Marinas de Andalucia, Espanha) , agradeço a am izade e bem com o t erem disponibilizado est ádios larvares provenient es de cult ivos laborat oriais, bem com o cópias dos seus trabalhos científicos antes m esm o da sua publicação.
À Junta Nacional de I nvestigação Científica e Tecnológica ( JNI CT) agradeço o financiam ento concedido através das Bolsas de estudo BD/ 1720/ 91- I G do program a CI ÊNCI A e BD/ 2617/ 94 do program a PRAXI S XXI .
CRUSTACEA DECAPODA LARVAE OFF THE PORTUGUESE COAST
The m ain obj ectives of this work were the identification of larval st ages of decapod crustaceans occurring off the Port uguese coast and the determ ination of occurrence pat t erns ( spat io- t em poral distribution and abundance) of the larval form s in this group along the Portuguese coast. About 700 sam ples from 35 scientific cam paigns carried out by I PI MAR ( I nst it ut o de I nvestigação das Pescas e do Mar) were analysed, corresponding t o a t ot al sam pling period from June 1986 t o March 1994.
I n order t o est ablish guidelines for t he descript ion of t he m orphology of larval st ages, t he nam es usually applicable for each phase and larval stage were standardised and each stage was defined. Also, a key for the ident ificat ion of larval developm ental phases within the Decapoda is presented.
The m orphological characterisation for the different larval stages for all taxa ident ified in t he sam ples, in part icular new larval form s is also present ed.
This work provides the first revision of the Decapod Crustacean larvae, which occur in t he coast al plankt on off t he Port uguese coast. Furtherm ore, for the first t im e in Port uguese, t he keys t o t he identification of the zoeal Decapoda genera, protozoa and zoea of Dendrobranchiat a were present ed. These keys will be useful in fut ure works on the ecology, biogeography and evaluation of stocks for sustainable fisheries.
I n order to correctly identify each taxon, useful inform at ion based on actual knowledge of each larval stage is presented. For the fist tim e 25 new larval form s have been described, and from a total of 168 taxa, 119 were identified to species level. This work present s larval form s from a t ot al of 50 decapod fam ilies.
Special at t ent ion was m ade t o t he larval st ages belonging t o t he species of high com m ercial value. These are: Arist eus ant ennat us ( red shrim p) , Arist aeom orpha foliacea ( giant red shrim p) , Parapenaeus longirost ris ( deep- water rose shrim p) , Nephrops norvegicus ( Norway lobst er) , Palinurus elephas ( com m on spiny lobst er) and Maj a squinado ( spinous spider crab) .
The pattern of occurrence for these species showed that the larvae of three of them are present in the plankton during the winter m onths. They are: A. foliacea in January, N. norvegicus from January to April and P. elephas from February t o March. On the ot her hand, only A. ant ennat us larvae collected appeared in August. Maj a squinado was present over a long period, from March to Septem ber and P. longirost ris has two reproduction periods during the year, one in the winter ( Novem ber- Decem ber) and the other in the sum m er ( August) .
throughout the year were: Solenocera m em branacea, Alpheus glaber, Thoralus cranchii, Eualus occult us, Processa m odica, P. nouveli, Processa EFSL11, Pandalina brevirost ris, Philocheras bispinosus, P. t rispinosus, Upogebia delt aura, Pisidia longicornis, Diogenes pugilat or, Anapagurus spp., Pagurus spp., Galat hea dispersa, G. int erm edia, Polybiinae, Coryst es cassivelaunus, At elecyclus rot undat us, Carcinus m aenas, Goneplax rhom boides, Ebalia spp. and Pilum nus spp.
All these taxa presented periods of great abundances followed by periods of scarcity. The great m aj ority are m ost abundant in the sum m er, for exam ple, At hanas nit escens, Alpheus glaber, Thoralus cranchii, Eualus occult us, Philocheras bispinosus neglect us and Diogenes pugilat or. Others presented a longer periods of great abundance, for exam ple, Processa canaliculat a, Pandalina brevirost ris and Philocheras sculpt us, which were predom inant during spring and sum m er, and Philocheras t rispinosus during autum n and winter. Som e species are m ost abundant in wint er, such as t he t axa belonging t o the sub- fam ily Polybiinae or t he t axa of genera Ebalia and Carcinus m aenas. Galat hea dispersa, G. int erm edia and Goneplax rhom boides present two periods of m axim um abundance during the year, t he first in spring ( March t o May) and t he second in aut um n ( Sept em ber t o Novem ber) .
Of the species with a lim ited period of occurrence of larvae in the plankton, the great m aj ority presented an enlarged period which includes two or even three seasons. Thus, Callianassa t yrrhena was present all the year except during the wint er, Pagurus bernhardus and Coryst es cassivelaunus, are also present during all t he seasons except in aut um n and finally Munida t enuim ana was not present during the sum m er m onths. Port um nus lat ipes, Pachygrapsus m arm orat us and the Xant ho species only appear in spring and sum m er. Processa EM5 and Callianassa t runcat a was present in plankt on only in t he sum m er and autum n. Som e species that occurred in only one season, were also not ed. Such as Processa EFSL11, Thia scut ellat a and Ast henognat hus at lant icus which only appeared in t he sum m er, Munida rugosa and At elecyclus rot undat us in spring and Gennadas elegans in wint er.
Este trabalho tem com o obj ectivo fazer a caracterização m orfológica dos diferentes estádios larvares das espécies de crustáceos decápodes da costa portuguesa, e a apresentação de chaves de identificação. Esta caracterização com preende não só as form as larvares j á descritas com o ainda novas form as larvares para a ciência e cuj a correspondência ao adulto é incerta e chaves de identificação para todas as espécies presentes nas am ostras e a determ inação dos padrões de ocorrência, da distribuição espaço- tem poral e da abundância dos estádios larvares ao longo de toda a costa continental portuguesa, de m odo a perm it ir um m elhor conhecim ento da sua ecologia e desta m aneira a fornecer um a base de trabalho para futuros trabalhos sobre estas larvas.
O cum prim ento dos obj ectivos foi feito com base na análise das larvas de Crustacea Decapoda provenientes de cerca de 700 am ostras de zooplâncton obtidas em 35 cam panhas de investigação realizadas pelo I PI MAR ( I nst it ut o de I nvestigação das Pescas e do Mar) , correspondendo a um período total de am ostragem de cruzeiros efectuados de Junho de 1986 a Março de 1994, e incluindo t oda a cost a cont inent al port uguesa.
Est e t rabalho const it ui um a prim eira revisão sobre as larvas de Crustáceos Decápodes que ocorrem no plâncton da costa continental portuguesa. Nele se apresenta tam bém pela prim eira vez chaves dicotóm icas para a identificação dos zoé dos géneros de decápodes, e t am bém para os estádios protozoé e estádios zoé de Dendrobranchiata que serão úteis em futuros trabalhos de ecologia, biogeografia e avaliação de recursos para efeitos de pesca sustentável.
Faz- se um a pequena descrição da m orfologia dos estádios larvares dos Decapoda e sugere- se um a uniform ização dos nom es em pregues em cada fase e estádio larvar e a definição de cada estádio larvar. Apresenta- se um a chave para a identificação das fases de desenvolvim ent o larvar.
Obteve- se a caracterização m orfológica dos diferentes estádios larvares das espécies de crustáceos decápodes da cost a port uguesa, incluindo novas form as larvares.
Este trabalho tam bém fornece indicações úteis sobre a identificação dos estádios larvares dos taxa analisados na observação das am ostras, utilizando o conhecim ento actual sobre cada form a larvar. Descreveram - se pela prim eira vez 25 taxa, conseguindo a identificação de 119 espécies. Descrevem - se e apresentam - se as figuras das form as larvares representantes de 50 fam ílias de Decápodes.
Foi dado um a especial atenção aos estádios larvares pertencentes a 6 espécies com grande valor com ercial: Arist eus ant ennat us ( cam arão verm elho) , Arist aeom orpha foliacea ( cam arão púrpura) , Parapenaeus longirost ris ( gam ba branca) , Nephrops norvegicus ( lagost im ) , Palinurus elephas ( lagosta) e Maj a squinado ( sant ola) .
Setem bro, para a presença de larvas no plâncton. P. longirost ris apresenta dois períodos de reprodução ao longo do ano, um no I nverno ( Novem bro- Dezem bro) e outro no Verão ( Agosto) .
Foram encontrados e identificados, por grupo taxonóm ico, 22 taxa de Dendrobrachiata, 2 taxa de Stenopodidea, 53 de Caridea, 1 de Astacidea (Nephrops norvegicus) , 10 de Thalassinidea, 4 de Palinura, 26 de Anom ura e 50 de Brachyura. O grupo dos Brachyura foi o m ais abundante.
Para os t axa m ais com uns faz- se a determ inação dos padrões de ocorrência, distribuição espaço- tem poral e abundância dos estádios larvares ao longo da costa portuguesa. Destes, os que tiveram as abundâncias m ais elevadas e que estiveram presentes no plâncton durante todo o ano, foram : Solenocera m em branacea, Alpheus glaber, Thoralus cranchii, Eualus occult us, Processa m odica, P. nouveli, Processa EFSL11, Pandalina brevirost ris, Philocheras bispinosus, P. t rispinosus, Upogebia delt aura, Pisidia longicornis, Diogenes pugilat or, Anapagurus spp., Pagurus spp., Galat hea dispersa, G. int erm edia, Polybiinae, Coryst es cassivelaunus, At elecyclus rot undat us, Carcinus m aenas, Goneplax rhom boides, Ebalia spp. e Pilum nus spp.
Todos estes taxa caracterizaram - se, no entanto, por terem picos de grande abundância seguidos de períodos de escassez. Na costa continental portuguesa a grande m aioria dos taxa tiveram a sua abundância m áxim a no Verão, com o foi o caso de At hanas nit escens, Alpheus glaber, Thoralus cranchii, Eualus occult us, Philocheras bispinosus neglect us e Diogenes pugilat or. Outras apresentaram um período m ais alargado de m aior abundância e que englobou a Prim avera- Verão ( por exem plo: Processa canaliculat a, Pandalina brevirost ris e Philocheras sculpt us) , ou Outono- I nverno ( por exem plo: Philocheras t rispinosus) . Algum as tiveram o m áxim o de abundância no I nverno, com o se verificou na generalidade dos t axa da sub-fam ília Polybiinae, igualm ent e para os t axa do género Ebalia e tam bém para Carcinus m aenas. De salientar, o facto das espécies Galat hea dispersa, G. int erm edia e Goneplax rhom boides apresentarem dois m áxim os de abundância ao longo do ano, o prim eiro na Prim avera ( Março a Maio) e o out ro no Outono ( Setem bro a Novem bro) .
Das espécies restritas a um a só estação do ano distinguiu- se: Processa EFSL11, Thia scut ellat a e Ast henognat hus at lant icus que ocorreram som ente no Verão, Munida rugosa e At elecyclus rot undat us de presença prim averis e Gennadas elegans a única espécie restrita ao I nverno.
Da representação espacial das larvas de decápodes para as am ostras m ensais das 4 secções da costa, no período de Outubro de 1986 a Janeiro de 1989, verificou- se a norte de Lisboa um m áxim o de abundância nas estações interm édias e valores m enores nas estações situadas nos extrem os das secções. Nas secções a sul de Lisboa, registou- se um a gradação da abundância larvar das estações m ais próxim as da cost a, para um m ínim o nas m ais afastadas da costa. Supõe- se que esta diferença estej a m uito provavelm ente relacionada com os fenóm enos de dispersão que afectam m uito a distribuição das larvas de crustáceos e que podem ser influenciados pela orografia do local e pelo sistem a de correntes e m assas de água da costa portuguesa.
Agradecim entos Abstract
Resum o Í ndice
I N TRODUÇÃO
I . I nt rodução e obj ect ivos
...
2MATERI AL E MÉTODOS I I . Mat erial e Mét odos ... 6
1. Material ... 6
2. Mét odos ... 8
2.1. Procedim ento laborat orial ... 8
2.2. Tratam ento dos dados ... 8
2.3. Medições ... 9
2.4. Figuras ... 10
2.5. Classificação dos Decapoda ... 10
2.6. I dentificação das form as larvares ... 10
MORFOLOGI A E N OMEN CLATURA DAS LARVAS I I I . Morfologia e nom enclat ura das larvas ... 13
1. .I ntrodução ... 13
2. .Morfologia geral das larvas ... 13
2.1. .. Cefalotórax, abdóm en e t elson ... 13
2.2. .. Apêndices ... 15
3. .Nom enclatura dos estádios larvares ... 17
3.1. .. Nom es dos estádios larvares ... 17
3.2. Núm ero de estádios larvares ... 18
3.3. .. Definição dos estádios larvares da fase Zoé ... 18
3.4. .. Chave para a identificação das fases de desenvolvim ento larvar ... 19
RESULTADOS E DI SCUSSÃO I V. Result ados e discussão ... 21
PARTE A- I dent ificação, dist ribuição e abundância dos est ádios larvares 21
2.2. .. Chave para a identificação dos estádios protozoé I I e I I I de
Dendrobranchiata da Cost a Portuguesa ... 43
2.3. .. Chave para a identificação dos estádios zoé de Dendrobranchiata da Costa Portuguesa ... 45
2.4. .. Chave de identificação para zoé I de Caridea ... 47
3. ... Diagnose, Referências, Dist ribuição e Abundância ... 51
Fam ília ARI STEI DAE Wood- Mason, 1891 ... 51
Fam ília BENTHESI SCYMI DAE Wood- Mason, 1891 ... 53
Fam ília PENAEI DAE Rafinesque, 1815 ... 61
Fam ília SI CYONI I DAE Ortm ann, 1898 ... 63
Fam ília SOLENOCERI DAE Wood- Mason e Alcock, 1891 ... 64
Fam ília SERGESTI DAE Dana, 1852 ... 65
Fam ília LUCI FERI DAE Dana, 1852 ... 72
Fam ília STENOPODI DAE Claus, 1872 ... 73
Fam ília NEMATOCARCI NI DAE Sm it h, 1884 ... 76
Fam ília OPLOPHORI DAE Dana, 1852 ... 77
Fam ília PASI PHAEI DAE Dana, 1852 ... 79
Fam ília RHYNCHOCI NETI DAE Ortm ann, 1890 ... 80
Fam ília PALAEMONI DAE Rafinesque, 1815 ... 81
Fam ília ALPHEI DAE Rafinesque, 1815 ... 88
Fam ília HI PPOLYTI DAE Dana, 1852 ... 93
Fam ília PROCESSI DAE Ortm ann, 1896 ... 99
Fam ília PANDALI DAE Hawort h, 1825 ... 107
Fam ília CRANGONI DAE Haworth, 1825 ... 116
Fam ília NEPHROPI DAE Dana, 1852 ... 127
Fam ília AXI I DAE Huxley, 1879 ... 129
3.1. Chave para a identificação das larvas de Callianassidae presentes nas águas da costa continent al port uguesa ... 131
Fam ília CALLI ANASSI DAE Dana, 1852 ... 131
Fam ília LAOMEDI I DAE Borradaile, 1903 ... 140
3.2. .. Chave para a identificação das larvas de Upogebia presentes nas águas da cost a cont inent al port uguesa ... 141
Fam ília UPOGEBI I DAE Borradaile, 1903 ... 141
Fam ília POLYCHELI DAE Wood- Mason, 1874 ... 145
Fam ília PALI NURI DAE Latreille, 1803 ... 147
3.3. Chave para a identificação das larvas dos géneros da fam ília Scyllaridae presentes na costa continent al port uguesa ... 148
Fam ília SCYLLARI DAE Latreille, 1825 ... 148
Fam ília PORCELLANI DAE Haworth, 1825 ... 175
Fam ília HOMOLI DAE De Haan, 1839 ... 178
Fam ília DORI PPI DAE MacLeay, 1838 ... 180
Fam ília CALAPPI DAE De Haan, 1833 ... 181
Fam ília LEUCOSI I DAE Sam ouelle, 1819 ... 183
Fam ília MAJI DAE Sam ouelle, 1819 ... 186
Fam ília PARTHENOPI DAE MacLeay, 1838 ... 197
Fam ília ATELECYCLI DAE Ortm ann, 1893 ... 199
Fam ília CORYSTI DAE Sam ouelle, 1819 ... 200
Fam ília PI RI MELI DAE Alcock, 1899 ... 201
Fam ília THI I DAE Dana, 1852 ... 204
Fam ília GERYONI DAE Colosi, 1923 ... 205
Fam ília PORTUNI DAE Rafinesque, 1815 ... 207
Fam ília XANTHI DAE MacLeay, 1838 ... 213
Fam ília ERI PHI I DAE MacLeay, 1838... 217
Fam ília PI LUMNI DAE Sam ouelle, 1819 ... 217
Fam ília GONEPLACI DAE MacLeay, 1838 ... 219
Fam ília ASTHENOGNATHI DAE St im pson, 1858 ... 222
Fam ília PI NNOTHERI DAE De Haan, 1833 ... 223
Fam ília GRAPSI DAE MacLeay, 1838 ... 225
Fam ília OCYPODI DAE Rafinesque, 1815 ... 232
PARTE B- Análise global das larvas de Crust áceos Decápodes ... 233
1. Com posição taxonóm ica na costa continental port uguesa ... 234
2. I m portância dos diversos grupos dent ro dos Decapoda ... 235
2.1. Dendrobranchiat a ... 236
2.2. Caridea ... 237
2.3. Thalassinidea ... 237
2.4. Anom ura ... 238
2.5. Brachyura ... 239
3. Padrões de distribuição das larvas ... 239
3.1. Tipos de sazonalidade ... 239
3.1.1. Espécies com larvas presentes todo o ano no plâncton ... 239
3.1.2. Espécies com época de reprodução lim it ada ... 240
Espécies de alt o valor com ercial ... 240
3.2. Dist ribuição espacial ... 241
4. Ciclo anual das variações ao longo da costa ... 243
4.1. Janeiro ... 245
4.2. Fevereiro ... 245
4.3. Março ... 246
4.7. Julho ... 250
4.8. Agosto ... 251
4.9. Setem bro ... 252
4.10. Out ubro ... 253
4.11. Novem bro ... 254
4.12. Dezem bro ... 255
CON CLUSÕES V. Conclusões ... 258
REFERÊN CI AS BI BLI OGRÁFI CAS VI . Referências bibliográficas ... 262
AN EXO
VI I . Anexo
O Universo é feit o essencialm ent e de coisa nenhum a. I ntervalos, distâncias, buracos, porosidade etérea. Espaço vazio, em sum a.
O resto, é a m atéria.
Daí, que este arrepio,
este cham á- lo e tê- lo, erguê- lo e defrontá- lo, est a frest a de nada aberta no vazio,
deve ser um intervalo.
I . I N TRODUÇÃO E OBJECTI VOS
A im portância dos crustáceos decápodes é elevada pelo interesse generalizado do Hom em no consum o de certas espécies que, por isso, se tornaram alvo de pescas dirigidas ( lagostas, lavagantes, cam arões e caranguej os) . Para além disto os crustáceos decápodes são um elo da teia alim entar de certas espécies de peixes dem ersais, com o, por exem plo, o bacalhau, o peixe- galo e a pescada ( Daan, 1973; Larrañeta, 1970; Pereda, Olaso e González, 1981; Silva, 1999) .
Em Portugal, o volum e das capturas de crustáceos no continente representa cerca de 0.85% do total das capturas de pescado; porém o seu valor económ ico atinge valores m édios de cerca de oito vezes superiores aos valores m édios de prim eira venda do total da pesca de peixes, crustáceos e m oluscos ( valores de 1998, Direcção- Geral das Pescas e Aquicultura, 1999) .
O I nstituto de I nvestigação das Pescas e do Mar ( I PI MAR) , no âm bito das suas atribuições, tem desenvolvido estudos sobre biologia e avaliação dos "stocks" de crustáceos decápodes ( Cascalho, 1988; Castro, 1988; Figueiredo, 1986) , sobre os quais assenta a gestão das pescarias respectivas. Os estudos de biologia realizados até agora apenas se referem à fase explorada da vida destes anim ais, isto é, estádios j uvenil e adulto.
Porém , o conhecim ento da biologia das espécies fica incom pleto sem o estudo da sua fase larvar. A vida larvar tem um a grande im portância dado que o "stock" explorável depende da sobrevivência dos estádios larvares.
No caso de espécies bentónicas m arinhas com larvas pelágicas, um bom conhecim ento da vida larvar é indispensável, j á que as larvas, de m orfologia e tipo de vida m uito diferente dos adultos, constituem um a grande parte do m eroplâncton e representam , a este nível, um a ligação essencial entre o plâncton e o bentos. De facto, a im portância da fase planctónica é considerável no ciclo vital das espécies ( Bourdillon- Casanova, 1960) .
No caso dos crustáceos decápodes da costa portuguesa, particularm ente as espécies de profundidade, o conhecim ento da vida larvar poderá esclarecer dúvidas ou confirm ar hipóteses form uladas sobre a distribuição destas espécies ( Cascalho, 1988) , nom eadam ente a sua relação com a deslocação da água m editerrânica int erm édia em profundidade, j unt o ao t alude cont inent al port uguês.
A ut ilização dest es est udos larvares pode t er valor com o “ indicadores biológicos” de m assas de água de origens diversas, que se sabe existirem ao longo da costa portuguesa e cuj o percurso e deslocações sazonais são alvo de investigações em curso ( Fiúza, et al., 1998; Leal, et al., 1999; Peliz e Fiúza, 1999) .
ut ilizar m odelos de previsão para a ocorrência e abundância das espécies. No que diz respeito às larvas de decápodes, este trabalho ainda não está feito na costa portuguesa, havendo alguns trabalhos em m eios lagunares, est uarinos ou j unt o à costa ( Gonçalves e Ribeiro, 1993; Morgado, 1993; Paula, 1987a; Paula, 1993) . Ao contrário de outras regiões da Europa, ainda não foi feit o um inventário de todas as form as larvares existentes na costa portuguesa.
A identificação de larvas de decápodes a partir de am ostras provenientes do plâncton é ainda hoj e um a tarefa difícil por existirem m uitas espécies para as quais não se conhece a sua fase larvar com o por exem plo, o caso da m aioria das espécies de profundidade. Na costa continental portuguesa este problem a torna- se m ais grave porque esta apresenta um a grande riqueza específica j á que é um a área de transição entre as águas do Atlântico Norte, águas de origem sub- tropical e o Mediterrâneo apresentando assim espécies de variadas proveniências.
Um a m aneira de resolver o problem a da ident ificação das larvas de decápodes é fazer o seu desenvolvim ento em laboratório desde a eclosão do ovo até ao prim eiro estádio j uvenil. Esta prática foi iniciada por M. V. Lebour em 1927 e generalizou- se m ais t arde, m uit o principalm ente, para espécies de Brachyura de zonas costeiras. No entanto, existem m uitas espécies que não são fáceis de cultivar por serem m uit o sensíveis ao t ipo de alim ent o fornecido ( por exem plo larvas de Grapsidae) ou por serem espécies de profundidade, por exem plo Aristeidae, torna-se m uit o difícil capturar as fêm eas vivas e m antê- las no laboratório.
Além disso, m uitas vezes as larvas cultivadas em laboratório apresentam pequenas diferenças m orfológicas relativam ente às larvas das m esm as espécies capturadas no plâncton o que dificulta a identificação das larvas capturadas do plânct on.
Assim , para a identificação de m uitas espécies ainda hoj e se recorre a toda a bibliografia exist ent e m esm o que as descrições de larvas sej am por vezes incom pletas e os autores dessas descrições m isturem várias espécies, porque na altura do estudo não tinham um conhecim ento com pleto do total de espécies existentes nas respectivas áreas.
O presente trabalho tem com o obj ectivos:
1. A caracterização m orfológica dos diferentes estádios larvares das espécies de crustáceos decápodes da costa portuguesa, com a apresentação de chaves de identificação, para as que não as possuíam . Esta caracterização com preende não só as form as larvares j á descritas com o ainda novas form as larvares para a ciência e cuj a correspondência a um dado adulto é incerta.
I I . MATERI AL E MÉTODOS
1 . MATERI AL
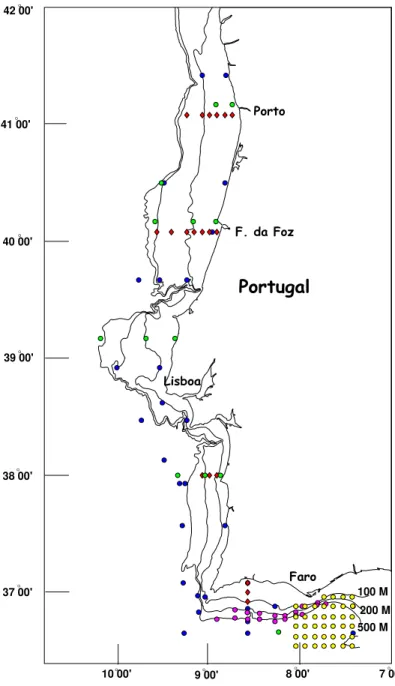
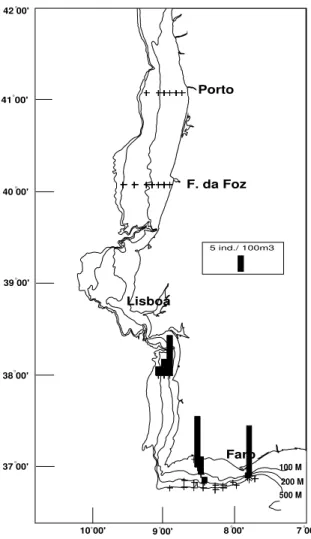
As larvas obj ecto deste trabalho são provenientes das cerca de 700 am ostras de zooplâncton obtidas em 35 cam panhas de investigação realizadas pelo I PI MAR ( I nstituto de I nvestigação das Pescas e do Mar) . O período total de am ostragem com preende cruzeiros efectuados de Junho de 1986 a Março de 1994, e inclui t oda a costa continental portuguesa ( ver figura 1 e tabela 1) .
100 M 200 M 500 M
Portugal
Lisboa
F. da Foz Porto
Faro 42 00'
41 00'
40 00'
39 00'
38 00'
37 00'
7 00' 8 00'
9 00' 10 00'
°
°
°
°
°
°
° ° ° °
Tabela 1 Apresentação das principais características das cam panhas de investigação utilizadas neste estudo.
Principais caract eríst icas das cam panhas de invest igação N .º da
Cam panha Sím bolo no m apa Período de t em po est ações N .º de Tipo de rede
0 2 1 1 1 0 8 6 ♦• 8- 26/ 10/ 1986 20 FAO de 1m
0 1 1 2 1 1 8 6 ♦ 12/ 11/ 1986 20 Bongo de 60cm
0 2 1 2 1 1 8 6 ♦ 7- 21/ 11/ 1986 20 Bongo de 60cm
0 2 1 3 1 2 8 6 ♦ 6- 20/ 12/ 1986 20 Bongo de 60cm
0 2 0 1 0 1 8 7 ♦ 16- 18/ 01/ 1987 20 Bongo de 60cm
0 2 0 2 0 2 8 7 ♦ 14- 16/ 02/ 1987 20 Bongo de 60cm
0 1 0 2 0 3 8 7 ♦ 19- 23/ 03/ 1987 20 Bongo de 60cm
0 2 0 4 0 4 8 7 ♦ 25/ 04- 10/ 05/ 1987 20 Bongo de 60cm
0 1 0 3 0 6 8 7 ♦ 12- 15/ 06/ 1987 20 Bongo de 60cm
0 1 0 5 0 7 8 7 ♦ 21- 23/ 07/ 1987 20 Bongo de 60cm
0 2 1 0 0 8 8 7 ♦ 16- 26/ 08/ 1987 20 Bongo de 60cm
0 1 1 0 0 9 8 7 ♦ 6- 22/ 09/ 1987 20 Bongo de 60cm
0 2 1 2 0 9 8 7 ♦ 22/ 09/ 1987 20 Bongo de 60cm
0 2 1 3 1 0 8 7 ♦• 12- 22/ 10/ 1987 20 Bongo de 60cm
0 2 1 4 1 0 8 7 ♦ 28/ 10- 15/ 11/ 1987 20 Bongo de 60cm
0 1 0 1 0 1 8 8 ♦ 9- 11/ 01/ 1988 20 Bongo de 60cm
0 2 0 3 0 2 8 8 ♦ 3- 29/ 03/ 1988 20 Bongo de 60cm
0 1 0 8 0 4 8 8 ♦ 5- 7/ 04/ 1988 20 Bongo de 60cm
0 1 1 2 0 5 8 8 ♦ 18- 20/ 05/ 1988 20 Bongo de 60cm
0 1 1 6 0 6 8 8 ♦ 25/ 06/ 1988 20 Bongo de 60cm
0 2 1 1 0 7 8 8 ♦ 22/ 07- 6/ 08/ 1988 20 Bongo de 60cm
0 1 2 0 0 8 8 8 ♦ 29- 30/ 08/ 1988 20 Bongo de 60cm
0 2 1 3 0 8 8 8 ♦ 08/ 09/ 1988 20 Bongo de 60cm
0 2 1 5 1 0 8 8 ♦• 5- 20/ 11/ 1988 20 Bongo de 60cm
0 1 2 9 1 2 8 8 ♦ 6- 8/ 12/ 1988 20 Bongo de 60cm
0 1 0 1 0 1 8 9 ♦ 11- 13/ 01/ 1989 20 Bongo de 60cm
0 2 0 8 0 6 8 6 • 9/ 06- 1/ 07/ 1986 30 FAO de 1m
0 2 0 9 0 7 8 7 • 28/ 07- 10/ 08/ 1987 30 Bongo de 60cm e FAO de 1m
0 2 0 4 0 7 9 0 • 5- 31/ 07/ 1990 13 Bongo de 60cm
0 1 1 7 0 8 9 3 • 5- 11/ 08/ 1993 18 Bongo de 60cm
0 1 0 2 0 2 9 0 • 7- 17/ 02/ 1990 40 Bongo de 60cm
0 1 0 1 0 1 9 1 • 22- 28/ 01/ 1991 40 Bongo de 60cm
0 1 0 1 0 1 9 4 • 11- 21/ 01/ 1994 39 FAO de 1m e I KMT
0 1 0 4 0 2 9 4 • 19- 24/ 02/ 1994 40 FAO de 1m e I KMT
0 1 0 7 0 3 9 4 • 25- 30/ 03/ 1994 40 FAO de 1m e I KMT
do fundo à superfície ( os fundos variando de 500 m a cerca de 20 m de profundidade) . As am ostras do sím bolo • foram colhidas em arrastos verticais de próxim o do fundo à superfície. Relat ivam ente às am ostras com o sím bolo • ut ilizaram - se várias est rat égias consoant e o ano em que se realizaram as cam panhas. Em 1990 fizeram - se dois arrastos por estação, um do fundo à superfície e out ro de 50m acim a do fundo até à superfície. Em 1991 t am bém se fizeram dois arrastos verticais por cada estação, um do fundo at é à superfície e o outro de cerca dos 450m de profundidade até à superfície. Finalm ente, em 1994 ut ilizaram - se dois t ipos de rede, um a rede de t ipo FAO de 1m de diâm et ro e um a rede I KMT ( I saacs Kidd Midwat er Trawl) . Nas est ações em que se ut ilizou a rede FAO fizeram - se dois arrastos verticais por cada estação, um do fundo at é à superfície e outro de cerca de 450m de profundidade até à superfície, no caso da I KMT fez- se um arrast o oblíquo do fundo à superfície.
Em t odas est as am ost ras foram ut ilizados fluxóm et ros Hidrobios para a determ inação do volum e de água filtrada por arrasto. No caso das redes FAO e I KMT a m alha da rede foi de 500 µm . A rede Bongo, pela sua própria constituição, tem duas redes, um a com 305 µm e a outra com 500 µm de vazio de m alha e nelas foram observadas som ente as am ostras provenientes da rede de 305 µm , tendo sido observadas as de 500 µm nos casos em que apareceram form as larvares raras. A escolha da rede com m alha m ais apertada teve em vista o obj ectivo de observar o m aior núm ero possível de est ádios iniciais ( prot ozoés) das larvas de espécies de valor económ ico, que são m uito pequenas e frágeis.
2 . MÉTODOS
2 .1 . Procedim ent o Laborat orial
2 .2 . Trat am ent o dos dados
Os valores provenient es das cont agens foram depois transform ados em concentrações segundo a fórm ula:
X = ( n x f) / V,
onde:
X- abundância por m3;
n- núm ero de indivíduos cont ados;
f- valor do fraccionam ento quando realizado; V- volum e de água filt rado pela rede.
Com o os valores obtidos apresentavam para cada espécie quantidades m uito díspares, procedeu- se à sua logaritim ização, utilizando a transform ação considerada m ais válida por I banez ( 1971) para norm alizar a distribuição de abundância de organism os plânctónicos:
Log ( X + 1) .
Para o tratam ento dos dados de abundância e dist ribuição ut ilizou- se unicam ente os dados provenientes das cam panhas identificadas na Tabela 1 com o sím bolo ♦. Esta escolha deveu- se ao facto destas cam panhas terem m antido os m esm os m étodos e estações de recolha, com um a periodicidade m ensal de Outubro de 1986 a Janeiro de 1989. Este tratam ento no entanto só foi realizado para os taxa m uit o com uns ( presentes em todas as estações am ostradas) . Para os restantes taxa, e sem pre que tal se verificou necessário, fez- se referência a todas as estações am ostradas.
2 .3 . Medições
Sem pre que necessário foram efectuadas as seguintes m edições:
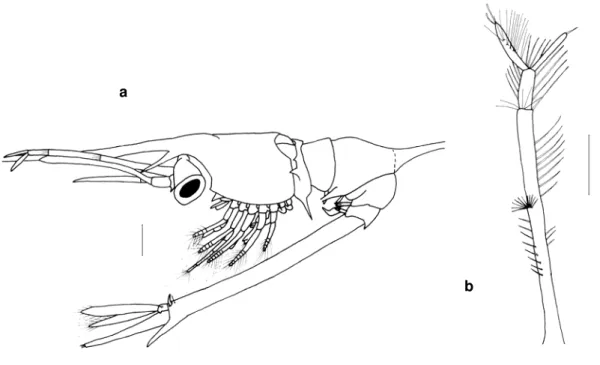
CT- Com prim ento total do exem plar, que vai da extrem idade do rostro até à m argem posterior do telson, sem considerar os espinhos deste. No caso das larvas de Brachyura este com prim ento foi m edido desde a m argem frontal, entre os olhos, até à ext rem idade da furca ( fig. 2) .
CC- Com prim ento do cefalotórax, que vai da m argem anterior à m argem posterior, sem rostro ( fig. 2) .
Todas estas m edições foram feitas com a lupa WI LD- M8 com ocular m icrom étrica.
CT CC
LC
Figura 2 Apresentação das m edições efectuadas nas larvas. CT- com prim ento total; CC- com prim ento da carapaça; LC- largura da carapaça.
2 .4 . Figuras
Todas as figuras foram realizadas pela autora e apresentam som ente as características m ais im portantes para a identificação dos exem plares. As que representam um a vista geral dos exem plares foram feitas com a lupa WI LD- M8 equipada de um a cam ara lucida. As figuras respeitantes aos apêndices foram feitas com o m icroscópio ZEI SS equipado com um a cam ara lucida.
2 .5 . Classificação dos Decapoda
2 .6 . I dent ificação das form as larvares
A ident ificação dos est ádios larvares foi feita com o recurso a todos os trabalhos disponíveis sobre cada taxa larvar. No entanto, ao trabalhar- se com form as larvares, acontece que por vezes se encontram exem plares para os quais não é possível est abelecer a sua correspondência com os adultos. Para estes exem plares terá de se estabelecer um a classificação tem porária que perm ita ordená- los até ser possível a sua nom eação posterior.
Para estes casos não existe nenhum conj unto de regras estabelecidas oficialm ent e. Decidiu- se, port ant o, seguir as regras estabelecidas por autores consagrados (e.g. William son, 1967 e, 1970; Rice e William son, 1977) , que consist e em est abelecer um a ligação de um determ inado exem plar a um género, sub- fam ília ou fam ília, seguido do prefixo E ( de Espinho) , F ( de Figueira da Foz) , S ( de Sines) , L ( de Lagos) ou A ( de Algarve) , relativo à área onde foi encontrado e finalm ente de um núm ero de ordem dentro do trabalho. Por exem plo: Rhynchocinet es A7 significa que as form as larvares com este nom e apresentam características que a colocam dent ro do género Rhynchocinet es, que as m esm as foram encontradas ao largo da costa algarvia e que, neste trabalho, é a sétim a descrição de um a nova form a larvar.
I I I . MORFOLOGI A E N OMEN CLATURA DAS LARVAS
1 . I N TRODUÇÃO
Nos prim eiros trabalhos efectuados sobre larvas de decápodes colhidas no plâncton foi- lhes dado um nom e específico, pois o autor pensou que se tratariam de adultos ( William son, 1982) . A causa disto foi não se saber ao tem po que o desenvolvim ento pós- em brionário dos decápodes inclui um a ou m ais m etam orfoses. Estas rápidas m udanças da m orfologia da larva durante o seu desenvolvim ento dificulta a sua correcta identificação. Tam bém o núm ero de fases larvares e consequentem ente o núm ero de m etam orfoses não é constante nos Decapoda. Sobre o núm ero e tipo de m etam orfoses nos Crustacea e particularm ente nos Decapoda William son ( 1982) j á fez um a análise detalhada.
Excepto as espécies de Decapoda que apresentam um desenvolvim ento directo ( passam do ovo para um indivíduo que é sim ilar ao adulto) , na grande m aioria dos casos o ovo eclode sob a form a de um a larva, m uito diferente do adulto, que apresenta um desenvolvim ento irregular, envolvendo pequenas e grandes variações na sua m orfologia. As grandes variações na m orfologia, aqui cham adas de m etam orfoses, fazem a passagem de um a fase larvar para outra e finalm ente para a form a j uvenil cuj o aspecto m orfológico é j á o de um adulto e im plicam um a m udança radical no aspecto geral da larva. As pequenas variações correspondem a um crescim ento gradual dentro de um a m esm a fase e que pode ser constituído por um ou m ais estádios de desenvolvim ento. Este crescim ento gradual não im plica um a m udança radical na form a do indivíduo m as sim pequenas m udanças relacionadas com o crescim ento e m ais visíveis ao nível dos apêndices.
Assim , de m aneira a facilit ar um a m aior com preensão dos t erm os em pregues no presente trabalho, faz- se de seguida um a pequena descrição da m orfologia dos estádios larvares dos Decapoda e dos nom es em geral em pregues em cada fase e estádio larvar.
2 . MORFOLOGI A GERAL DAS LARVAS
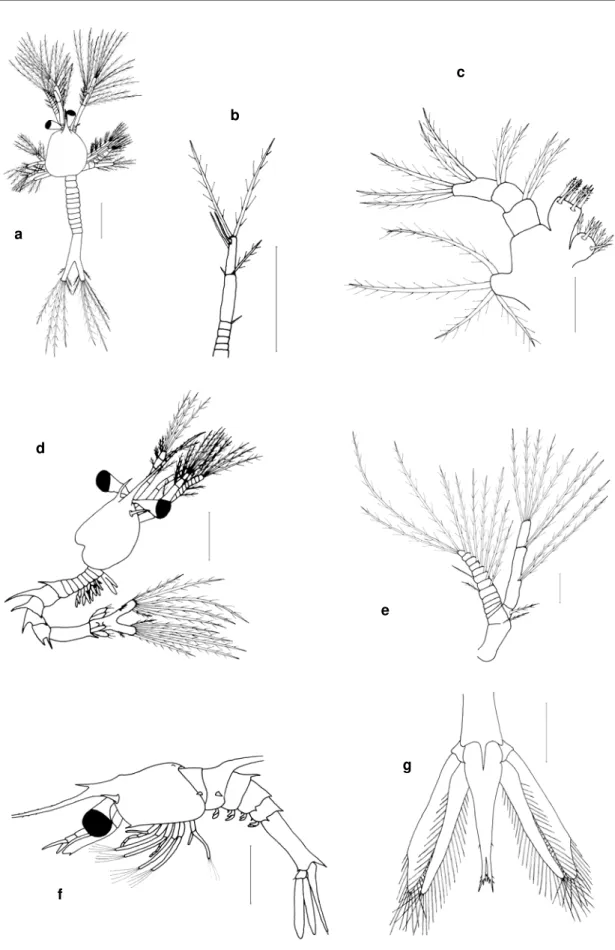
2 .1 . Cefalot órax, abdóm en e t elson ( fig. 3)
( m axilulas e m axilas) . Os t rês prim eiros segm entos torácicos apresentam três pares de m axilipedes e os rest ant es segm ent os t êm cinco pares de pereiópodes. A carapaça pode possuir rostro, carena m édio- dorsal, dentículos nas m argens anteriores e/ ou posteriores, espinho dorsal, espinhos laterais, espinhos supra- orbitais e pterigostom ianos e protuberâncias.
Cefalotórax Abdóm en od r A1 A2 d sa ex p pl e u t s en pr pe el sp ed f
Figura 3 Vista lateral de um a larva de Caridea (a) e do abdóm en de um a larva de Brachyura (b) : A1- antenula; A2- antena; d- dentículos; e- espinho; ed- espinho dorsal; el- espinho lateral; en- endopódito; ex- exopódito; f- furca; od- orgão dorsal; p- pereiópodes; pe- pêlos; pl- pleópodes; pr- processo; r- rostro; s- sedas; sa- segm ento abdom inal; sp- seda plum osa; t- telson; u- urópodes.
Abdóm en: parte posterior do corpo constituída por seis segm entos articulados, nos estádios protozoé e nos dois prim eiros estádios zoé dos Pleocyem ata o sexto segm ento abdom inal está fundido com o telson. Nos estádios zoé m ais avançados os segm entos 1- 5 possuem um par de pleópodes e o segm ento seis tem um par de urópodes. Os segm entos abdom inais podem ter carena dorsal e/ ou espinhos m édio-dorsais, laterais, sedas, dentículos e protuberâncias dorso- laterais.
Telson: parte term inal do corpo sem apêndices e que na sua parte ventral apresenta a abertura anal. Na sua m argem posterior possui um conj unto variável de sedas plum osas que são contadas da m argem exterior para o centro, por vezes tem um a seda m édia. Os Stenopodidea, Thalassinidea e Anom ura têm , em vez da segunda seda plum osa, um a seda anom úrica que consiste num a seda plum osa m uit o fina. O telson pode tam bém estar ornam entado por espinhos e sedas lisas dorsais e lat erais.
2 .2 . Apêndices
Apêndices ( figs. 4- 7) : Todos os apêndices são suj eitos a um processo de segm entação do estádio m ais j ovem para os estádios m ais avançados.
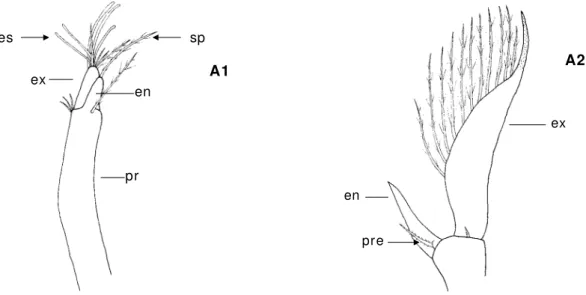
Antenula ( A1) - Um longo pedúnculo com posto por um exopódit o e um endopódito. O exopódito tem na sua extrem idade sedas lisas cham adas estetascos ( “ aesthetascs” ) ( quim ioreceptoras) e sedas plum osas ( m ecanoreceptoras) ( fig. 4) .
Antena ( A2) - Com posta por um pedúnculo curto com exopódito, endopódito e um processo espinhoso que pode estar m uit o desenvolvido na m aioria das espécies de Brachyura e que nas restantes espécies de decápodes apresenta- se com o um pequeno espinho. O exopódito na m aioria dos casos tem um a form a de escam a que pode estar segm entada e apresentar sedas plum osas. O endopódito é com posto por um flagelo ( fig. 4) .
pr ex
en
sp es
ex
en
pre
Figura 4 : A1- antenula ( ZI I de Callianassa t yrrhena) ; A2- antena ( ZI I de Galat hea FSL21) ;
en- endopódito; es- estetascos; ex- exopódito; pr- protopódito; pre- processo espinhoso; sp- seda plum osa.
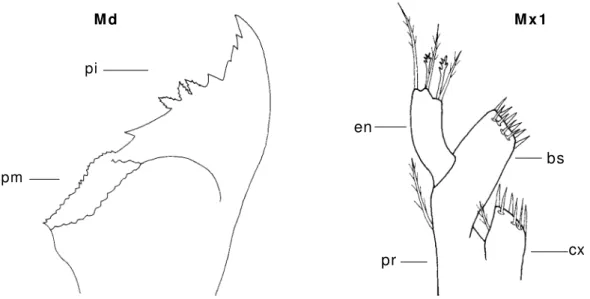
Mandíbula ( Md) - É com post a por um a base constituída por um a parte incisiva e por um a parte m olar ( fig. 5) .
pi
pm
pr en
cx bs
Figura 5 : Md- m andíbula ( ZV de Rhynchocinet es A7) ; Mx1- m axilula ( ZI V de Hippolyt e sp.) ;
bs- base; cx- coxa; en- endopódito; pi- parte incisiva; pm - parte m olar; pr- protopódito.
Maxilula ( Mx1) - É const it uída por um endopódito e um protopódito. Do protopódito erguem - se a base e a coxa ( fig. 5) .
Maxila ( Mx2) - A m axila dos estádios zoés é constituída por um endopódito, base, coxa e o escafognatite ( fig. 6) .
eg en
cx bs
sp
bs sp
en ex
Figura 6 :Mx2- m axila ( ZI de Maj inae ESL24) ; Mxp- 1º m axilipede ( ZI de Maj inae ESL24) ; bs- base; cx- coxa; eg- escafognatite; en- endopódito; ex- exopódito; sp- seda plum osa.
Maxilipedes ( Mxp1, Mxp2, Mxp3) - São apêndices birram osos em que o exopódito e o endopódito separam - se a partir da base. Os endopóditos são segm entados e os dois ram os possuem longas sedas plum osas ( fig. 6) .
Pereiópodes ( P1, P2, P3, P4, P5) - Est es podem ser birram osos ou unirram osos. O exopódito não é segm entado e o endopódito apresenta cinco segm entos: “ ischium ” ,
Md M x1
M x2
m ero, carpo, propódio e dáct ilo) . Os pereiópodes são constituídos de sedas e espinhos ( fig. 7) .
Pleópodes ( Pl1, Pl2, Pl3, Pl4, Pl5) - Só com eçam a aparecer nos estádios larvares m ais avançados com o pequenos gom os, no 1º estádio pós- larvar estão funcionais, são birram osos e possuem sedas plum osas.
Urópodes ( U) - São birram osos, tanto o endopódito com o o exopódito têm sedas plum osas e só aparecem quando o 6º segm ento abdom inal se separa do telson ( fig. 7) .
en ex
t
u
ex
en
Figura 7 : P- 1º pereiópode ( ZV de Philocheras SL14) ; U- urópodes ( ZI I I de Gennadas sp.B) ;
en- endopódito; ex- exopódito; t- telson; u- urópodes.
3 . N OM EN CLATURA DOS ESTÁDI OS LARVARES
3 .1 . N om es dos est ádios larvares
Não há um a clara uniform idade nos nom es dos diferentes estádios larvares, principalm ent e porque m uit os aut ores cont inuam a ut ilizar nom es de est ádios larvares que foram baptizados quando se pensava serem espécies novas. Quando se descobriu que eram sim plesm ente estádios larvares de espécies j á conhecidas, estes continuaram a ser descritos por esses nom es que acabaram por ser adoptados, até porque alguns dos nom es representavam form as larvares bizarras quando com paradas com a m aioria das restantes form as larvares ( por exem plo a larva de Jaxea noct urna que é m uit o conhecida com o “ t rachilifer” ) . Por isso nom es com o “ elaphocaris” , “ acanthosom a” e “ m astigopus” ( para as larvas de Sergestidae) , “ m ysis” ( para as larvas de Penaeoidea) , “ phyllosom a” ( para os estádios larvares de
Scyllaridae e Palinuridae) , “ t rachelifer” ( para as larvas do género Jaxea) , et c., cont inuam a ser ut ilizados ainda hoj e.
Sendo que os decápodes têm na sua m aioria um desenvolvim ento larvar com um , constituído por m udas, com um a ou duas m etam orfoses antes de atingirem o estádio j uvenil, William son ( 1969) , tentou uniform isar os nom es larvares com base no núm ero de m etam orfoses. Assim , tem os 3 fases larvares principais: nauplius, zoé e m egalopa ou decapodito. No caso dos Dendrobranchiata, o nom e protozoé é m uito utilizado para os 3 prim eiros estádios zoé ( Gurney, 1942 e William son, 1969) porque, nesta fase, a carapaça não cobre com pletam ente todos os segm entos torácicos. Por isso, est e t erm o será t am bém ut ilizado nest e t rabalho. Cada fase é com post a por um certo núm ero de estádios que se separam por m udas. A fase m egalopa nem sem pre se considera com o um a verdadeira larva pois j á apresenta em m uitos casos m uitas características das form as j uvenis ( por exem plo: Paguridae, Caridea, etc.) , sendo por isso cham ada de decapodito. Por isso m esm o, as m egalopas são raras em am ostras planctónicas, excepto m egalopas de Brachyura em épocas do ano bem definidas e não poderão ser abordadas em porm enor. A fase nauplius, que só existe nos Dendrobranchiata, tam bém não é tratada neste trabalho por não se encontrarem presentes nas am ostras, j á que os Dendrobranchiata que existem nas áreas prospectadas são indivíduos bentónicos ou pelágicos de profundidade e os nauplius destas espécies encontram - se em geral perto do fundo.
3 .2 . N úm ero de est ádios larvares
O núm ero de est ádios larvares t am bém varia, consoant e a fam ília, género ou m esm o a espécie, havendo m esm o espécies em que o desenvolvim ento é directo, quer dizer que o prim eiro indivíduo é j á um j uvenil. Considerando só os decápodes que têm ciclo de vida com fases larvares e as espécies das costas portuguesas, podem os afirm ar que os Dendrobranchiata têm em geral 3 estádios protozoé, 2- 4 estádios zoé, nos Pleocyem ata, os Stenopodidea e os Caridea têm entre 5- 9 estádios zoé, os Astacidea cerca de 3 estádios zoé, os Thalassinidea 2- 6 estádios zoé, os Palinura têm pelos m enos 8, podendo ter 15 ou m ais, os Anom ura e os Brachyura 2- 6 estádios zoé. Neste trabalho, os estádios zoé são indicados pela letra Z seguida de um núm ero rom ano correspondente ao estádio por ordem crescente, por exem plo: ZI significa prim eiro est ádio zoé.
3 .3 . Definição dos est ádios larvares da fase Zoé
Zoé I I ( Protozoé I I ) - Olhos m óveis, sem urópodes. No caso dos protozoé, e relativam ente ao estádio anterior, o abdóm en j á se apresenta segm entado. Zoé I I I ( Protozoé I I I ) - Urópodes presentes, 6º segm ento abdom inal j á separado do
t elson.
Zoé I V e seguintes- Apresentam m odificações adicionais que dependem da espécie.
3 .4 . Chave para a ident ificação das fases de desenvolvim ent o larvar
Esta chave de identificação tem por obj ectivo perm itir a identificação rápida da fase larvar em que se encontra determ inado espécim en.
1. a) Os três prim eiros pares de apêndices cefálicos com sedas ... Nauplius b) Alguns ou todos os apêndices torácicos com sedas, pleópodes quando presentes
sem sedas ... 2 ( Zoé) c) Todos os apêndices torácicos com sedas, pleópodes com sedas e funcionais
... Megalopa
I V. RESULTADOS E DI SCUSSÃO
PARTE A
1. CLASSI FI CAÇÃO DAS FORMAS LARVARES OBJECTO DO ESTUDO
Ordem Decapoda Latreille, 1803
Subordem Dendrobranchiata Bate, 1888
Superfam ília Penaeoidea Rafinesque- Schm altz, 1815 Fam ília Aristeidae Wood- Mason, 1891
Género Arist eus Duvernoy, 1840
Arist eus ant ennat us ( Risso, 1816)
Género Arist aeom orpha Wood- Mason, 1891
Arist aeom or pha foliacea ( Risso, 1827)
Fam ília Benthesicym idae Wood- Mason, 1891 Género Bent hysicym us Bate, 1881
Bent hysicym us A1
Género Gennadas Bate, 1881
Gennadas elegans ( Sm ith, 1882)
Gennadas sp.B ( Calazans, 1992)
Fam ília Penaeidae Rafinesque- Schm altz, 1815 Penaeidae L2
Género Parapenaeus Sm ith, 1885
Parapenaeus longirost ris ( Lucas, 1846)
Fam ília Sicyoniidae Ortm ann, 1898 Género Sicyonia H. M. Edwards, 1830
Sicyonia carinat a ( Brünnich, 1768)
Fam ília Solenoceridae Wood- Mason, 1891 Género Solenocera Lucas, 1849
Solenocera m em branacea ( Risso, 1816)
Superfam ília Sergestoidea Dana, 1852 Fam ília Luciferidae De Haan, 1849
Género Lucifer Thom pson, 1829
Lucifer t ypus H. M. Edwards, 1837
Fam ília Sergestidae Dana, 1852
Género Sergest es H.- M. Edwards, 1830
Sergest es henseni ( Ortm ann, 1893)
Sergest es curvat us Crosnier e Forest, 1973
Sergest es ar ct icus Kröyer, 1855
Sergest es at lant icus H. M. Edwards, 1830
Sergest es sargassi Ortm ann, 1893
Sergest es cornut us Kröyer, 1855
Sergest es vigilax Stim pson, 1860
Sergest es S3
Sergest es SL5
Género Ser gia Stim pson, 1860
Sergia robust a ( Sm ith, 1882)
Sergia j aponica ( Bate, 1881)
Subordem Pleocyem ata Burkenroad, 1963 I nfraordem Stenopodidea Claus, 1872
Fam ília Stenopodidae Claus, 1872
St enopus species I I I ( Gurney, 1936)
Stenopodidae A6 I nfraordem Caridea Dana, 1852
Superfam ília Atyoidea De Haan, 1849 Fam ília Nem atocarcinidae Sm ith, 1884
Género Nem at ocarcinus A.- M. Edwards, 1881
Nem at ocarcinus sp.
Fam ília Oplophoridae Dana, 1852
Género Acant hephyra A.- M. Edwards, 1881
Acant hephyr a sp.
Género Syst ellaspis Bate, 1888
Syst ellaspis debilis ( A.- M. Edwards, 1881)
Superfam ília Pasiphaeoidea Dana, 1852 Fam ília Pasiphaeidae Dana, 1852
Género Pasiphaea Savigny, 1816
Pasiphaea sivado Risso, 1816
Superfam ília Rhynchocinetoidea Ortm ann, 1890 Fam ília Rhynchocinetidae Ortm ann, 1890
Rhynchocinet es A7
Superfam ília Palaeom onoidea Rafinesque, 1815 Fam ília Palaeom onidae Rafinesque, 1815
Género Palaem on Weber, 1795
Palaem on ser rat us ( Pennant, 1777)
Palaem on elegans Rathke, 1837
Palaem on longirost ris Zariquiey- Alvarez, 1968
Palaem on adspersus Rathke, 1837
Género Pont onia Latreille, 1829
Pont onia flavom aculat a Heller, 1864
Género Periclim enes O. G. Costa, 1844
Periclim enes sp. ( B.- Casanova, 1960)
Mesocaris F8
Superfam ília Alpheoidea Rafinesque, 1815 Fam ília Alpheidae Rafinesque, 1815
Género At hanas Leach, 1814
At hanas nit escens ( Leach, 1814)
Alpheus glaber ( Olivi, 1792)
Alpheidae L9
Género Synalpheus Bate, 1888
Synalpheus ESL10
Fam ília Hippolytidae Dana, 1852 Género Caridion Goës, 1863
Caridion gordoni ( Bate, 1859)
Caridion st ev eni Lebour, 1930
Género Thor alus Holthuis, 1947
Thoralus cranchii ( Leach, 1817)
Thoralus sp. ( Pike e William son, 1961)
Género Eualus Thallwitz, 1892
Eualus occult us ( Lebour, 1936)
Eualus pusiolus ( Kröyer, 1841)
Género Hippolyt e Leach, 1814
Hippolyt e sp.
Género Lysm at a Risso, 1816
Lysm at a set icaudat a ( Risso, 1816)
Fam ília Processidae Ortm ann, 1896 Género Processa Leach, 1815
Processa canaliculat a Leach, 1815
Processa eduliscrassipes Nouvel e Holthuis, 1957
Processa m odica m odica William son, 1979
Processa m odica carolii William son, 1979
Processa nouveli holt huisi Al- Adhub e William son, 1975
Processa nouveli nouveli Al- Adhub e William son, 1975
Processa EM5 ( William son, 1967)
Processa EM6 ( William son, 1967)
Processa EFSL11
Superfam ília Pandaloidea Haworth, 1825 Fam ília Pandalidae Haworth, 1825
Género Plesionika Bate, 1882
Plesionika sp. ( Kurian, 1956)
Plesionika FSL12
Género Dichelopandalus Caullery, 1896
Dichelopandalus bonnieri Caullery, 1896
Género St ylopandalus ?Borradaille, 1899?
St ylopandalus richardi ( Coutière, 1905)
Género Pandalina Calm an, 1899
Pandalina br evirost ris Rat hke, 1843
Pandalidae Sp.1 ( Barnich, 1996a) Pandalidae E13
Fam ília Crangonidae Haworth, 1825 Género Crangon Fabricius, 1798
Crangon crangon Linnaeus, 1758
Crangon allm ani Kinahan, 1857
Género Pont ocaris Bate, 1888
Pont ocaris cat aphract a ( Olivi, 1792)
Pont ocaris lacazei ( Gourret, 1887)
Género Pont ophilus Leach, 1817
Pont ophilus norvegicus ( Sars, 1861)
Pont ophilus spinosus ( Leach, 1815)
Género Philocheras Stebbing, 1900
Philocheras bispinosus bispinosus ( Hailstone, 1835)
Philocheras bispinosus neglect us G. O. Sars, 1886
Philocheras fasciat us ( Risso, 1816)
Philocheras sculpt us Bell, 1847
Philocheras t rispinosus ( Hailstone, 1835)
Philocheras echinulat us ( Sars, 1862)
Philocheras SL14
I nfraordem Astacidea Latreille, 1803
Superfam ília Nephropoidea Dana, 1852 Fam ília Nephropidae Dana, 1852
Género Nephrops Leach, 1814
Nephrops norvegicus ( Linnaeus, 1758)
I nfraordem Thalassinidea Lat reille, 1831
Superfam ília Thalassinoidea Latreille, 1831 Fam ília Axiidae Huxley, 1879
Género Axius Leach, 1815
Axius st irhynchus Leach, 1815
Axiidae E15
Fam ília Callianassidae Dana 1852 Género Callianassa Leach, 1814
?Callianassa t runcat a ( Giard e Bonnier, 1890)
Callianassidae SL16
?Callianassa t yrrhena ( Pet agna, 1792)
Callianassidae L17
Fam ília Laom ediidae Borradaile, 1903 Género Jaxea Nardo, 1847
Jaxea noct urna Nardo, 1947
Fam ília Upogebiidae Borradaile, 1903 Género Upogebia Leach, 1814
Upogebia delt aura ( Leach, 1815)
Upogebia EM13 ( William son, 1967)
I nfraordem Palinura Latreille, 1903
Superfam ília Eryonoidea De Haan, 1841 Fam ília Polychelidae Wood- Mason, 1874
Género Polycheles Heller, 1862
Polycheles A19
Superfam ília Palinuroidea Lat reille, 1803 Fam ília Palinuridae Latreille, 1803
Género Palinurus Weber, 1795
Palinurus elephas ( Fabricius, 1787)
Fam ília Scyllaridae Latreille, 1825 Género Scyllarus Fabricius, 1775
Scyllarus arct us ( Linnaeus, 1758)
Scyllarus A20
I nfraordem Anom ura H.- M. Edwards, 1832
Superfam ília Coenobitoidea Dana, 1851 Fam ília Diogenidae Ortm ann, 1892
Género Clibanarius Dana, 1852
Clibanarius eryt hropus ( Latreille, 1818)
Género Pagurist es Dana, 1851
Pagurist es er em it a ( Linnaeus, 1767)
Género Dardanus Paulson, 1875
Dardanus arrosor ( Herbst, 1796)
Género Diogenes Dana, 1851
Diogenes pugilat or ( Roux, 1829)
Género Calcinus Dana, 1851
Calcinus t ubularis ( Linnaeus, 1767)
Superfam ília Paguroidea Latreille, 1803 Fam ília Paguridae Latreille, 1803
Género Pagurus Fabricius, 1775
Pagurus bernhardus ( Linnaeus, 1758)
Pagurus alat us ( Fabricius, 1775)
Pagurus spp.
Género Nem at opagurus A. M.- Edwards e Bouvier, 1892
Nem at opagurus longicornis A. M.- Edwards e Bouvier, 1892
Género Cest opagurus Bouvier, 1897
Cest opagurus t im idus ( Roux, 1830)
Género Spiropagurus Stim pson, 1858
Spiropagurus elegans Miers, 1881
Género Anapagurus Henderson, 1887
Anapagurus spp.
Superfam ília Galatheoidea Sam ouelle, 1819 Fam ília Galatheidae Sam ouelle, 1819
Galat hea dispersa Sp. Bate, 1859
Galat hea int erm edia Lillj eborg, 1851
Galat hea nex a Em bleton, 1834
Galat hea squam ifera Leach, 1814
Galat hea st rigosa Linnaeus, 1767
Galat hea FSL21
Galat hea S22
Galat hea L23
Género Munida Leach, 1820
Munida t enuim ana Sars, 1872
Munida rugosa ( Fabricius, 1775)
Munida sarsi Huus, 1935
Munida ASM9 ( Rice e William son, 1977)
Fam ília Porcellanidae Haworth, 1825 Género Pisidia Leach, 1820
Pisidia longicornis ( Linnaeus, 1767)
Género Porcellana Lam arck, 1801
Porcellana plat ycheles ( Pennant, 1777)
I nfraordem Brachyura Latreille, 1803 Secção Podot rem ata Guinot, 1977
Superfam ília Hom oloidea De Haan, 1839 Fam ília Hom olidae De Haan, 1839
Género Hom ola Leach, 1815
?Hom ola barbat a ( Fabricius, 1793) Secção Heterotrem ata Guinot, 1977
Superfam ília Dorippoidea MacLeay, 1838 Fam ília Dorippidae MacLeay, 1838
Género Et husa Roux, 1830
Et husa m ascarone ( Herbst, 1785)
Género Medorippe Manning e Holthuis, 1981
Medorippe lanat a ( Linnaeus, 1767)
Superfam ília Leucosioidea Sam ouelle, 1819 Fam ília Calappidae De Haan, 1833
Género Calappa Weber, 1795
Calappa granulat a ( Linnaeus, 1758)
Fam ília Leucosiidae Sam ouelle, 1819 Género Ebalia Leach, 1817
Ebalia t uberosa ( Pennant, 1777)
Ebalia spp.
Superfam ília Maj oidea Sam ouelle, 1819 Fam ília Maj idae Sam ouelle, 1819
Género Maj a Lam arck, 1801
Maj a golt ziana Oliveira, 1888
Maj a crispat a Risso, 1827
Género Pisa Leach, 1814
Pisa arm at a ( Lat reille, 1803)
Pisinae S9 ( Paula, 1987) Género Eurynom e Leach, 1814
Eurynom e aspera ( Pennant, 1777)
Eurynom e spinosa Hailstone, 1835
Género Achaeus Leach, 1817
Achaeus cranchii Leach, 1817
Género I nachus Weber, 1795
I nachus dorset t ensis ( Pennant, 1777)
I nachus phalangium ( Fabricius, 1775)
I nachus lept ochirus Leach, 1817
I nachus t horacicus ( Roux, 1830)
Género Macr opodia Leach, 1814
Macropodia spp.
Maj inae ESL24
Superfam ília Parthenopoidea MacLeay, 1838 Fam ília Parthenopidae MacLeay, 1838
Género Part henope Weber, 1795
Part henope m assena ( Roux, 1830)
Part henope S14 ( Paula, 1987)
Superfam ília Cancroidea Latreille, 1803 Fam ília Atelecyclidae Ortm ann, 1893
Género At elecyclus Leach, 1814
At elecyclus rot undat us ( Olivi, 1792)
Fam ília Corystidae Sam ouelle, 1819 Género Coryst es Bosc, 1801
Coryst es cassivelaunus ( Pennant, 1777)
Fam ília Pirim elidae Alcock, 1899 Género Pirim ela Leach, 1815
Pirim ela dent iculat a ( Mont agu, 1808)
Género Sir pus Gordon, 1953
Sirpus sp.
Fam ília Thiidae Dana, 1852 Género Thia Leach, 1815
Thia scut ellat a ( Fabricius, 1793)
Superfam ília Portunoidea Rafinesque, 1815 Fam ília Geryonidae Colosi, 1923
Género Gery on Kröyer, 1837
Geryon longipes A. M. Edwards, 1882
Género Carcinus Leach, 1814
Carcinus m aenas Linnaeus, 1758
Género Port um nus Leach, 1814
Port um nus lat ipes ( Pennant, 1777)
SubFam ília Polybiinae Ortm ann, 1893 Género Bat hynect es Stim pson, 1871
Bat hynect es m aravigna ( Prestandrea, 1839)
Género Liocarcinus Stim pson, 1870
Liocarcinus depurat or ( Linnaeus, 1758)
Género Necora Holthuis, 1987
Necora puber ( Linnaeus, 1767)
Género Polybius Leach 1820
Polybius henslowi Leach, 1820
Superfam ília Xanthoidea MacLeay, 1838 Fam ília Xanthidae MacLeay, 1838
Género Monodaeus Guinot, 1967
Monodaeus couchi ( Couch, 1851)
Género Nanocassiope Guinot, 1967
Nanocassiope m elanodact yla ( A. M. Edwards, 1867)
Género Xant ho Leach, 1815
Xant ho incisus ( Leach, 1814)
Xant ho spp.
Fam ília Eriphiidae MacLeay, 1838 Género Eriphia Lat reille, 1817
Eriphia verrucosa ( Forskäl, 1775)
Fam ília Pilum nidae Sam ouelle, 1819 Género Pilum nus Leach, 1815
Pilum nus spp.
Superfam ília Goneplacoidea MacLeay, 1838 Fam ília Goneplacidae MacLeay, 1838
Género Goneplax Leach, 1814
Goneplax rhom boides ( Linnaeus, 1758)
Goneplacidae L25
Superfam ília Pinnotheroidea De Haan, 1833 Fam ília Asthenognathidae Stim pson, 1858
Género Ast henognat hus Stim pson, 1858
Ast henognat hus at lant icus Monod, 1932
Fam ília Pinnotheridae De Haan, 1833 Género Pinnot heres Bosc, 1802
Pinnot heres pinnot heres ( Linnaeus, 1758)
Pinnot heres pisum ( Linnaeus, 1758)
Secção Thoracotrem ata Guinot, 1977
Fam ília Grapsidae MacLeay, 1838
Plagusinae ASM29 ( Rice e William son, 1977) Género Pachygrapsus Randall, 1840
Pachygrapsus m arm orat us ( Fabricius, 1787)
Género Brachynot us De Haan, 1833
Brachynot us sexdent at us ( Risso, 1827)
Superfam ília Ocypodoidea Rafinesque, 1815 Fam ília Ocypodidae Rafinesque, 1815
Género Uca Leach, 1814
2. CHAVES DE I DENTI FI CAÇÃO PARA AS LARVAS DE DECAPODA DA
COSTA PORTUGUESA
A identificação de larvas de crustáceos decápodes a partir de am ostras do plâncton não é tarefa fácil pois além de não se conhecer a m orfologia das larvas respeitante a todas as espécies, tam bém é frequente a sua alteração de um estádio larvar para out ro. Em geral as chaves de ident ificação exist ent es são parciais, feit as para um grupo específico de espécies: e.g. Paula ( 1996) para as larvas de Brachyura; para um determ inado estádio larvar e.g. Calazans ( 1993) para os vários estádios larvares dos Penaeidea, etc. As chaves de identificação que consideram todas as fam ílias de decápodes são as de William son ( 1957a) para latitudes de 40° - 80° N, o que não inclui toda a costa continental portuguesa e as de Barnich ( 1996a) para o Mediterrâneo.
Assim , com o obj ectivo de facilit ar o trabalho de identificação de larvas de crustáceos decápodes e de contribuir para o estím ulo do estudo deste im port ante grupo taxonóm ico, apresenta- se um a chave de identificação para os estádios zoé de todos os géneros observados nas am ostras, baseada nos estudados no presente trabalho e na chave de William son ( 1957) . Tam bém se incluem um a chave para os zoé I de Caridea, um a para os protozoé e outra para os zoé de Dendrobranchiata.
As chaves de identificação pretendem - se sim ples e baseadas em caracteres fáceis de distinguir com um a sim ples lupa, recorrendo- se excepcionalm ente ao m icroscópio. O texto é acom panhado por figuras para m elhor ilustrar as diferenças taxonóm icas que se pretende observar e para que a identificação se torne m ais fácil até para um principiante. A chave por si só não garante um a identificação sem pre correcta das espécies e é por vezes necessário recorrer a descrições com pletas e porm enorizadas das espécies alvo. Portanto, apresenta- se para cada espécie, inform ações relat ivas à m elhor font e de inform ação para a identificação correcta e com plem ent ar de cada espécie e, quando esta não existe, faz- se um a descrição dos estádios larvares observados, sem pre acom panhada de ilustrações. Além das inform ações relat ivas à taxonom ia e m orfologia larvar, tam bém se fornecem resultados sobre a sua distribuição e abundância na cost a continental portuguesa e, sem pre que possível, com param - se com a distribuição e abundância noutras regiões, onde determ inada espécie tam bém ocorre. Tam bém se ut ilizam dados referent es à distribuição e época de reprodução dos adultos.