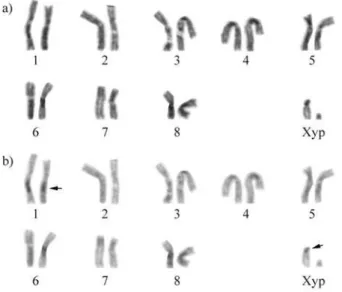
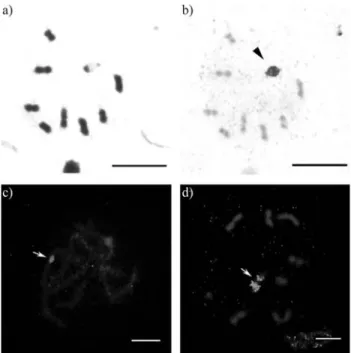
Cytogenetic analysis of Astylus antis (Perty, 1830) (Coleoptera, Melyridae): Karyotype, heterochromatin and location of ribosomal genes
Texto
Imagem
Documentos relacionados
Abstract: This paper proposes a systematic method for the simultaneous determination of adjusted ranks of sample observations and their sums and products adjusted for
This fact led us to select these profes- sionals for a cytogenetic biomonitoring of exposure to X-rays, using as parameters the analysis of the frequencies of chromosome aberration
Cytogenetic analysis of Trichomycterus areolatus , collected from the Tijeral and Huilma Rivers in southern Chile has shown a diploid chromosome number of 2n = 54, a fundamental
 ðåçóëüòàòå ïîëó÷àþò èíôîðìàöèþ î ìèòîòè÷åñêîì (ôàçû ìèòîçà, êîëè÷åñòâî ÿäåð íà êëåò- êó, ìèêðîÿäðà, ïàòîëîãèè ìèòîçà) è âèòàëüíîì (ìèòîòè÷åñêèé èíäåêñ, àïîïòîç)
The cytogenetic analysis of Characidium showed a conserved diploid number of 2n=50, with a predominance of metacentric and submetacentric chromosomes (Figs. In Characidium cf.
acrobata showed the same diploid number, fundamental number and chromosome morphology as Kerodon rupestris.. But its NOR location and heterochromatin distribution patterns
The cytogenetic characterization of the chromosome number, meiosis and karyotype were all performed with the São Borja population, and an analysis of pollen fertility was
This work aimed to evaluate the interaction between Astylus variegatus (Germar 1824) (Coleoptera: Melyridae) and the nasturtium flowers associated to the weeks of flourishing, in
