Scientific Note
Screening for detection of wild oat biotypes resistant to fenoxaprop-P-ethyl from Mexico
Hugo Cruz-Hipolito1*, José A. Domínguez-Valenzuela2, Fidel González-Torralva3 and Rafael De Prado4
1
Technical Representative at Bayer, Mexico, [email protected]; 2 Professor at University of Chapingo, Mexico,
[email protected]; 2 Post-graduate student at University of Cordoba, Spain, [email protected]; 4 Professor at University of Cordoba, Spain, [email protected].
*Author for correspondence
Avena is one of the world’s most important genera resistant to herbicides. Suspected resistant biotype and susceptible biotype of Avena fatua were tested by screening to confirm the resistance to fenoxaprop-P-ethyl from Mexico. Suspected-resistant population was collected in wheat fields after the herbicide failed to control wild oat and the susceptible one was collected from a field with no herbicide application. Petri dish experiments were performed using different fenoxaprop-P-ethyl concentrations (0, 1, 5, 10 and 40 M). Experiments were arranged in a completely randomized design with ten replicates. Plumule length in both biotypes decreased as fenoxaprop-P-ethyl concentration increased. However, there was a different response between the R and S biotypes. The EC50 for the resistant biotype was 3.3 M while for the susceptible biotype was 9.3 M, showing that the resistant biotype tolerated herbicide concentration approximately three times higher than the susceptible biotype. The results confirm the presence of Avena fatua resistant to fenoxaprop-P-ethyl in Mexico.
Keywords: Avena fatua, ACCase-inhibitor herbicide, EC50, herbicide resistance.
Introduction
Wheat is the most widely cultivated cereal in the world and occupies 17% of the world's cultivated land. It is the staple food of 35% of the world's population. However, approximately more than 90% of wheat fields have problems with weed infestation (Beckie et al. 2012). One of the most important weeds in wheat crop is Avena fatua, commonly known as wild oat, which is probably native to Central Asia and is cited in 55 countries invading more than 20 crops (Haile and Girma 2010). The weed species has great adaptability as its life cycle is synchronized with the wheat (Mortimer 1985). At present, the species is ranked the first important grass weed in wheat and barley fields from Mexico (Cruz-Hipolito et al. 2011). Wild oat is also considered as the second most important weed in terms of resistance to herbicides (Heap 2012).
Within the control strategies for wild oat management, chemical control is the most widely used due to the large areas infested. Herbicides recommended for control began to have trouble since the 1980's (Cruz-Hipolito et al. 2011). The aryloxy-phenoxypropionate herbicides are typically used for post emergence grass weed control in many crops. These herbicides act through inhibition of Acetyl-CoA carboxylase (ACCase), which catalyzes the first step in fatty acid biosynthesis. Consecutive use of different herbicides with the same mode of action in wheat fields of Mexico has led to select resistant wild oat plants and increasing their density in the population. The first documented report of resistance in wild oat was in 1998 in Mexico (Heap 2012). Since then, the incidence has increased dramatically and steadily. The resistance mechanism is due to target site (Cruz-Hipolito et al. 2011). Actually, these resistant populations have been established in some parts of
the cereal areas and if the current weed-crop management strategy does not change, increasing in the selection pressure will result in a wider dispersion of resistant populations.
The commonly used experiment for herbicide resistance detection is dose-response. Dose-response experiments are time and space consuming (Ribeiro et al. 2008). Although there is an advantage to using multiple doses to improve the ability to detect populations which vary in the extent of their resistance, it is often difficult to justify the extra work involved in routine screening (Moss et al.1998, Beckie et al. 2000, Ribeiro et al. 2008). A substitute experiment was the screening test. Screening tests are quick tests that provide information about the response of a biotype to herbicide application, showing some advantages such as: small space, inexpensive and early response (Perez-Jones et al. 2007, Ribeiro et al. 2008).
The objective of this work was to quickly corroborate the fenoxaprop-P-ethyl resistance of wild oat from Mexico.
Material and methods
Plant material and growing conditions. Wild oat seeds suspected as resistant to ACCase-inhibitor herbicides were collected in wheat fields from Mexico and one biotype was also collected from the same region that had never been treated with herbicides (susceptible). Caryopses were dehulled by hand, then sterilized with a solution of sodium hypochlorite at 5%, to be placed in Petri dishes containing two sheets of sterile filter paper, then add the herbicide solutions (treated plants) or distilled water (to check treatments). Seeds were placed in four rows with a fold down and the end of the embryo oriented in one direction, according to the protocol of Friesen (2000).
Communications in Plant Sciences (2237-4027) volume 2, issue 1, p.11-13, Jan-Mar, 2012 12
Experiments were conducted using 9-cm-diam Petri dishes containing two sheets of filter paper, with 16 seeds per biotype scattered over the surface of the filter paper. Petri dishes were maintained in a growth chamber at 28/18 C (day/night) in a 16-h photoperiod under 850 µmol m-2 s-1 photosynthetic photon-flux density delivered by a mixture of incandescent and fluorescent lights, with an 80% relative humidity.
Experimental design and treatments. Four-milliliter aliquots of different fenoxaprop-P-ethyl concentrations (0, 1, 5, 10 and 40 M) were poured into the dishes, containing both seeds of susceptible and suspected-resistant plants, and transferred to the growth chamber. Experiments were arranged in a completely randomized design with ten replicates.
Evaluation. Seven days after treatment, seedling shoot length of plumule was measured for each biotype and expressed as the percentage of the untreated control. Measurement was performed in 16 seedlings, discarding the three highest and the three lower readings.
Statistical analysis. There was no interaction between treatments and experimental replicates detected by previous ANOVA test, so that all data were pooled across experimental replicates for further analysis and presentation. For each biotype, length of plumule data was fitted to a nonlinear, log-logistic regression model:
Y = c + {(d – c) / [1 + (x / g)b]}
where Y is the length of plumule expressed as a percentage of the untreated control, c and d are the coefficients corresponding to the lower and upper asymptotes, b is the slope of the line, g is the herbicide concentration at the point of inflection halfway between the upper and lower asymptotes, and x is the herbicide concentration used.
The herbicide concentration needed to inhibit length of plumule by 50% (EC50) compared to the untreated control can be determined, represented by the point of inflection of the regression equation above. The resistant factor (RF) was also computed as:
RF = EC50(R) / EC50(S)
Regression analysis was computed using Sigma Plot statistical software (Systat Software, version 10.0, USA).
Results and discussion
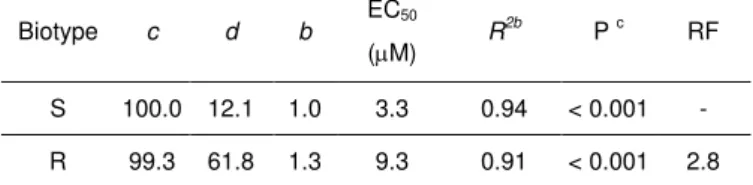
Length of plumule of susceptible (S) and suspect-resistant (R) seedlings of wild oat decreased when the fenoxaprop-P-ethyl concentration increased (Figure 1). However, there was a different response to herbicide concentration between the two wild oat biotypes. The EC50 for the S biotype was 3.3 M, while for the R biotype was 9.3 M (Table 1). Thus, the R biotype showed a resistance factor of 2.8, indicating that R biotype tolerated approximately three times more fenoxaprop-P-ethyl than S biotype.
In this research, the discriminatory concentration for fenoxaprop-P-ethyl screening of wild oat was 10 µM. Discriminating concentration of 28 µM
fenoxaprop-P-ethyl were recommended to detect R biotypes of
Alopecurus myosuroides while 21 µM fenoxaprop-P-ethyl for wild oat (Avena spp.) (Moss 2000). The screening bioassays for determining the effective dose are the basis to confirm the response of resistant or sensitive populations, which may take only from 7 up to 10 days (Ribeiro et al. 2008). The Petri dish assay has been used successfully to quantify herbicide resistance (Neve et al. 2004, Perez and Kogan 2003, Perez-Jones et al. 2007, Ribeiro et al. 2008). So, the Petri dish screening indicated that resistance to fenoxaprop-P-ethyl was present in the R biotype of wild oat from Mexico.
Figure 1. Plumule length reduction of fenoxaprop-P-ethyl-susceptible (S) and -resistant (R) Avena fatua biotypes as affected by fenoxaprop-P-ethyl concentration. Symbols and lines represent actual and predicted growth responses, respectively. Vertical bars represent ± standard errors of the mean. Cordoba, Spain, 2011.
By using Petri dish bioassays, some authors identified weed resistant biotypes to herbicides (Moss 2000, Neve et al. 2004, Perez and Kogan 2003, Perez-Jones et al. 2007, Ribeiro et al. 2008, Carvalho et al. 2011), also including cross resistance (Bourgeois et al. 1997), confirming the importance of Petri dish rapid tests. Bourgeois et al. (1997) identified three major types of cross-resistance in Avena species, namely (A) resistance to APP herbicides and little or no resistance to CHD herbicides; (B) low resistance to all herbicides; and (C) comparatively high resistance to all herbicides. None of the 82 Avena lines they studied had high CHD resistance and low APP resistance.
Table 1. Parameters of equationa
used to calculate the fenoxaprop-P-ethyl concentration required for 50% length of plumule reduction (EC50) in Avena fatua seedlings and the respective resistant factor (RF). Cordoba, Spain, 2011.
Biotype c d b EC50 (M) R
2b
P c RF
S 100.0 12.1 1.0 3.3 0.94 < 0.001 -
R 99.3 61.8 1.3 9.3 0.91 < 0.001 2.8
a Y = c + {(d – c) / [1 + (x / g)b]}, where Y is the percent of length of plumule
reduction; x (independent variable) is the herbicide concentration; c and d are
the lower and upper, asymptote, respectively; b is the slope of the curve; and
EC50 is the effective herbicide concentration required for 50% of length of
plumule reduction. Data were pooled and fitted to a non-linear regression
model. b Approximate coefficient of determination of non-linear models with a
defined intercept calculated as pseudo R2 = 1 – (sums of squares of the
regression/corrected total sums of squares). c Probability level of significance
of the non-linear model.
fenoxaprop (mM)
0,1 1 10 100
Plu
m
u
le
le
n
g
th
(
%
Co
n
tro
l)
0 20 40 60 80 100
S R Predicted S Predicted R
Communications in Plant Sciences (2237-4027) volume 2, issue 1, p.11-13, Jan-Mar, 2012 13
It is essential to know whether or not a lack of weed control after herbicide application results from herbicide resistance or other reasons (Duke and Powles 2008; Powles 2008). Using the identification of herbicide-resistant weeds as a first step in resistance management demands an efficient and effective screening test; only after obtaining an accurate diagnosis can the nature, distribution and abundance of resistant weed populations be monitored (Perez and Kogan 2003).
In addition, understanding the factors influencing the evolution of herbicide resistance will help delay the appearance of resistance (Preston et al. 2009). For wild oat, the main factors can be cited as intensive use of fenoxaprop-P-ethyl and the lack of other weed management practices. Orchards and fallow situations are at high risk of herbicide resistance evolution (Carvalho et al. 2011). Thus, changing management practices to include other weed control tactics will help delay the evolution of herbicide resistance in this species, whereas the use of fenoxaprop-P-ethyl as the only control method will exacerbate herbicide resistance.
Conclusion
The present study confirms the presence of wild oat (Avena fatua) resistant biotype to fenoxaprop-P-ethyl in Mexico.
References
Beckie HJ, Heap IM, Smeda, RJ, Hall LM. 2000. Screening of herbicide resistance in weeds. Weed Sci 14:428-445.
Beckie HJ, Warwick SI, Sauder CA. 2012. Basis for herbicide resistance in Canadian populations of wild oat (Avena fatua). Weed Sci 60:10-18.
Bourgeois L, Kenkel NC, Morrison IN. 1997. Characterization of cross-resistance patterns in acetyl-CoA carboxylase inhibitor resistant wild oat (Avena fatua). Weed Sci 45:750-755
Carvalho LB, Cruz-Hipolito H, González-Torralva F, Alves PLCA, Christofolleti PJ, De Prado R. 2011. Detection of sourgrass (Digitaria insularis) biotypes resistant to glyphosate in Brazil. Weed Sci 59:171-176.
Cruz-Hipolito H, Osuna MD, Domínguez-Valenzuela J, Espinoza N, De Prado R. 2011. Mechanism of resistance to ACCase-inhibiting herbicides in wild oat
(Avena fatua) from Latin America. J Agric Food Chem 59:7261-7267.
Duke SO, Powles SB. 2008. Glyphosate: a once in a century herbicide. Pest Manag Sci 64:319-325. Friesen LF, Jones TL, Van Acker, RC Morrison IN.
2000. Identification of wild oat (Avena fatua) resistance to imazamethabenz, flamprop, and fenoxaprop-P. Weed Sci 48:532-540.
Haile D, Girma F. 2010. Integrated effect of seeding rate, herbicide dosage and application timing on durum wheat (Triticum turgidum L. var durum) yield, yield components and wild oat (Avena fatua L.) control in South Eastern Ethiopia. Mom Ethiop J Sci 2:12-26.
Heap IM. 2012. International survey of herbicide-resistant weeds. http://www.weedscience.org. Acce-ssed: June 6, 2010.
Mortimer AM. 1985. Intractable weeds: a failure to appreciate ecological principles in weed control? In Proceedings of British Crop Protection Conference, Weeds. London: BCP. pp.377-386.
Moss SR. 2000. The Rothamsted rapid resistance test for detecting herbicide-resistance in annual grass weeds. Weed Sci Soc Am Abstr 40:42-43.
Neve P, Sadler J, Powles SB. 2004. Multiple herbicide resistance in a glyphosate-resistant rigid ryegrass (Lolium rigidum) biotype. Weed Sci 52:920-928. Perez, A. Kogan M. 2003. Glyphosate-resistant Lolium
multiflorum in Chilean orchards. Weed Res 43:12-19. Perez-Jones A, Park KW, Polge N, Colquhoun J,
Mallory-SmithCA.2007.Investigatingthe mechanisms of glyphosate resistance in Lolium multiflorum. Planta 226:395-404.
Powles SB. 2008. Review. Evolved glyphosate-resistant weeds around the world: lessons to be learnt. Pest Manag Sci 64:360-365.
Preston C, Wakelin AM, Dolman FC, Bostamam Y, Boutsalis P. 2009. A decade of glyphosate-resistant Lolium around the world: mechanisms, genes, fitness, and agronomic management. Weed Sci 57:435-441.
Ribeiro D, Gil ND, Cruz-Hipolito HE, Ruiz-Santaella JP, Christoffoleti PJ, Vidal RA, De Prado R. 2008. Rapid assays for detection of glyphosate-resistant Lolium spp. J Plant Dis Prot 21:95-99.
Received on February 26, 2012. Accepted on March 27, 2012. Online published on March 30, 2012.