U N IV E R S ID A D E
D E S A O P A U L O
IN S T IT U T O
D E F is IC A
D E
5 A O
C A R L O S
E s t u d o s c r is t a lo g r a f ic o s
e m m a c r o m o le c u la s
b io l6 g ic a s :
A p lic a 9 0 e s
e m C a lg r a n u lin a
C d e g r a n u l6 c it o s p o r c in o s ,
T r ip a n o t io n a
r e d u t a s e
d e
Trypanosoma cruzi
e F o s f o lip a s e
A
2e x t r a id a
d o v e n e n o
d a s e r p e n t e
Bothrops moojeni
LISP
JIFQSC !S~!
~IIIIIIII ~111111111111IIII ~IIIIII
~Iillill
3-2-001"163
Tese apresentada ao Instituto de
Fisica de Sao Carlos, Universidade de
Sao Paulo, para obtenc;ao do titulo de
Doutor em Ciencias, area de
concentrac;ao Fisica Aplicada.
S a o C a rlo s
-1997-" Estudos cristalograficos em macromoleculas biol6gicas: Calgranulina C de granul6citos porcinos, Tripanotiona redutase de T r y p a n o s s o m a c r u z i e
fosfolipase A2 extraida do veneno da serpente B o t h r o p s
m o o j e n i " / Maria Cristina Nonato - Sao Carlos, 1997
ill ~
Il~
rl=\;
=
UNIV~RSIDADE
~
u .r-~ _ J_ :J
D E S A O P A U L OIn s t it u t o d e F 1 s ic a d e S a o C a r lo s
Av. Dr. Carlos Botelho, 1465 CEP 13560-250 - Sao Carlos - SF Brasil
Fone (016) 272-6222 Fax (016) 272-2218
MEMBROS DA COMISSAO JULGADORA DA TESE DE DOUTORADO DE M A R IA
C R IS T IN A N O N A T O APRESENTADA AO INSTITUTO DE FfslCA DE SAO CARLOS,
UNIVERSIDADE DE SAO PAULO, EM 21/03/1997.
,r>"
( £ > /) ' / ..'- ~ )
"-"/' /1,\' ....•Jv.~, ...\ 1 . / /7 1 >-~l".I\,., y"\./-" .'
\...'-
--~---~---ProY:' Dr. Glaucius Oliva (IFSC-USP) ---~-~----~--~---Profa. Ora. Leila Maria Be amini (IFSC-USP)Prof. Dr. Aida ~IF-USP)
---~~---Prof. Dr. Carlos Frederico Martins Menck (ICB-USP)
, ~
" E
p r e c i s o a m a ras
p e s s o a s c o m oA 6 R A D £ C IM E N T O S
• Ao Prof. Dr. Willian N. Hunter pela oportunidade, paci€mcia e pela importante orientayao durante meu trabalho na Inglaterra;
• Ao Prof. Dr. Richard C. Garratt pela indescritivel ajuda no desenvolvimento deste trabalho e pel a amizade;
• A
Prof- D~ Yvonne P. Mascarenhas pelo apoio ao longo destes anos e pela amizade.•
A
Prof- Dr" Heloisa Araujo por toda a ajuda com os estudos sobre fosfolipases e pela amizade;• Ao pesquisador e amigo Christian Schleicher pela colaborayao com os estudos sobre Calgranulina C
• Aos meus amigos do laborat6rio em Manchester: Rachel, Mark, Stefano e Charlie pelas discussoes, ajuda e amizade;
• Aos meus amigos do laborat6rio de Sao Carlos: Paulao, Zac, Portezani, Bia, Bianca, pH, Diet, Dulce, Jorge, Teresa, Pavao, Juan, Gerard. Rodrigo, Frank, Paula Kuser, Valma, Gisele, Carlos, Fred, Silvia, Fontes e Delboni, pelas importantes discussoes, pelo clima agradavel de trabalho e pela amizade;
• Aos meus queridos amigos do outro lade do oceano, Leo, Miguel, Sylvie e Nick por tornarem meus dias na Inglaterra tao especiais;
S U M A R IO
C A P I T U L O
I
- I N T R O D U V A O
C A P IT U L O
II - C A L G R A N U U N A
C D £
G R A N U L O C IT O S
II. 1. Introduyao 10
11.2. Purificayao e cristalizayao 16
11.3. Coleta e processamento de dados 17
11.4. Simetria nao cristalografica 19
II.5. T entativas para a determinayao da estrutura 20
11.5.1Substituiyao Molecular (MR) 20
11.5.2Substituiyao isom6rfica (SIR/MIR) 22
11.5.3Dispersao anomala a multiplos comprimentos de
onda (MAD) 23
C A P IT U L O
I I I
T R I P A N O T I O N A
R £ D U T A S £
D E
TRYPANOSOMA CRUZI
111.1.Introducao 111.2.Cristalizacao
111.3.Coleta e processamento de dados
iliA. Determinacao da estrutura e refinamento 111.5.Resultados cristalograficos
111.6.Discuss6es e conclus6es 111.7.- Referencias
C A P IT U L O
n l -
F O S F O U P A S E
A 2
E X T R A i D A
D O
V E N E N O
D A S E R P E N T E
BOTHROPS
MOOJENI
IV.1. Introducao 63
IV.2. Cristaliza<;ao 66
IV.3. Coleta e processamento de dados 67
IVA. Simetria nao cristalografica (NCS) 68
IV.5. Determina<;ao da estrutura 69
IV.5.1 Substitui<;ao molecular (MR) 69
IV.5.2 Analise do empacotamento 71
IV.5. Refinamento 72
IV.7.1. Purifica980
IV.7.2 Novos ensaios de cristaliza980 IV.7.3. Sequenciamento N-terminal IV.7.4 Refinamento
IV.8. Discussoes e conclusoes IV.9. ReferE3ncias
7 6
7 9
81 81
82
87
C A P iT U L O
V -
COMENTAmos
F IN A lS
APENDICEA
APENDICEB
A P E N D IC E C
APENDICED
APENDICEE
A P E N D IC E f
APENDICE6
L lS T A
D E T A B E L A S
Ponto de precipita~o para a Calgranulina C para diferentes agentes precipitantes. 0 teste foi realizado com uma solu~o de proteina a
1 0 m g / m l em 1mM de sulfato de zinco ( 2nS04) e Hepes (CaH,aN204S)
como soluyao tampao a pH 7.0
Coleta de dados para os cristais na ausencia (cristaI1) e presen<;a (cristal 2) de calcio.
Estatistica da coleta de dados para cristais de Calgranulina C a 2.6A de resolu~o. Cristais #1 e #2 foram crescidos na ausencia e na presen<;a dos ions Ca2+ , respectivamente.
Lista de proteinas extraidas do banco de dados de coordenadas de proteinas (PDB) que apresentam a motivo "EF-hand"( helice-loop-helice).
Comportamento dos cristais diante das solu90es de atomos pesados testadas.
Estatisticas relevantes a coleta de dados. Rs"rr, = 1:
I
I(k)-(I) 11I:1(k) onde I(k) e (I) representam a intensidade de uma dada reflexao e a media das intensidades respectivamente ecr a desvio padrao.Valores parciais para as parametros termicos Isotr6picos para C53S TR + Gspd. NA e equivalente ao numero de atomos e S a , ao fator de
Uma lista das possiveis liga¢es de hidrogemio formadas entre 0 Iigante N1
-glutationil-espermidina, tripanotiona redutase de T C r u z ie moleculas de solvente. Somente contatos ate 3.5A foram considerados. A and 8 correspondem as diferentes subunidades.
Uma lista das possiveis contatos de Van der Waals formadas entre 0 ligante N1-glutationil-espermidina, Tripanotiona redutase de T C r u z ;e moleculas de solvente. Somente contatos ate 4.oA foram considerados. A and 8 correspondem as diferentes subunidades
Fator de discord6ancia Rrs entre 0 modele e a densidade eletronica para GSPd em ambos sitios ativos para 0 mapa 2Fo-Fc ao nivel de 1.0cr.
Desvios posicionais quanta a superposiyao entre as estruturas C58S TR +Gspd TR, TR nativa e TR complexada com (GSPdh
Estatlsticas relevantes
a
coleta de dados. Rsym=
2:I
l(k)-<I)/I:I(k) onde I(k) e (I) representam a intensidade de uma dada reflex80 e a media das intensidades, respectivamente e cr 0 desvio padrao.Soluyao da substituiyao molecular para dois dimeros. ex, ~ e y definem a rota<;80 e X, Y e Z a transla<;80 a ser aplicada no modele
U S T A
D £
F I6 U R A S
Diagrama esquematico do motivo liEF-hand" de ligayao de calcio (a) simbolizado pela mao direita (b).
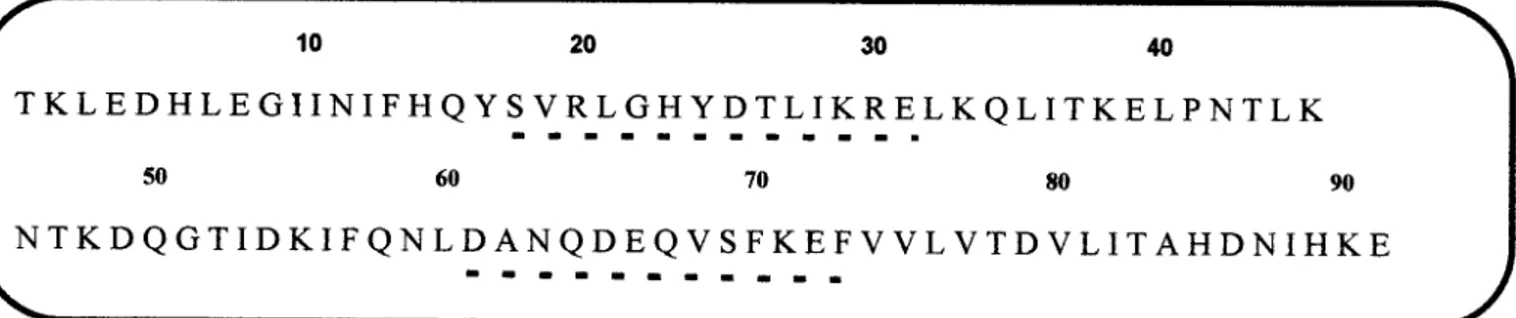
Estrutura primaria da calgranulina C. As linhas pontilhadas indicam os dois motivos liEF-hands" (HLH).
Alinhamento sequencial entre proteinas que ligam calcio do tipo S100 usando 0 sistema de Darwin (Gonnet, G.H., 1992). p - C A G C
-Calgranulina C porcina, h - C A G B - Calgranulina 8 humana, h C A G A
-Calgranulina A humana, p - C a B P 9 K - Calbindina-D9k porcina, b - S 1 0 0 / 3
-Cadeia ~ da proteina S100 bovina, h - C A C L - proteina que Iiga calcio
de placenta humana, b - S 1 0 0 L - S100L bovina, h - C A C Y - Calciclina
humana, b - S 1 0 0 u - Cadeia u da proteina S100 humana, h S 1 0 0 P
-SlOOP humana, p - C a [ 1 ] - Cadeia leve da calpactina I p o r c i n a , h - S 1 0 0 D
-S100D humana, h - S 1 0 0 E - S100E humana, p - S 1 0 0 C - S100C porcina.
Projeyao estereogratica ao Iongo do eixo c. seyao polar k=180°, da funyao de auto-rotayao calculadas com dados do cristal 1 no intervalo de 2.6-15A.
Diagrama esquematico para indicar a regra de prote~o biol6gica no metabolismo dos tripanossomatideos e onde a enzima tripanotiona esta envolvida.
Alinhamento sequencial entre TR de T r y p a n o s o m a c r u z i e GR
humana. Os residuos mais importantes para a catalise estao salientados em vermelho.
Vista estereografica da estrutura cristalografica do complexo mutante C58S TR +GSPd.
Distribuic;ao dos fatores de temperatura ao longo das cadeias principais e laterais para ambas subunidades.
Mapa de amissae (omissao (Fobs-Fcalc, cxcalc" mapa calculado excluindo as coordenadas da serina 58) ao nfvel de 3s.
Vista estereografica para GSPd e seu ajuste com 0 mapa de omissao Fobs-Fcalc ( calculo do mapa omitindo as coordendas do ligante no modele ao nivel 3 c r. a) subunidade A b) subunidade B.
Superposi~o entre as moleculas de GSPd para as duas subunidades. Em azul subunidade A e em vermelho subunidade B.
Distribui~o ao longo das subunidades para Rrs (espac;o real), isto e, 0 desvio entre a densidade eletronica e 0 modelo. Em rosa para a subunidade A, em azul para a subunidade B.
Fenilalanina 46 e seu correspondente mapa de amissae Fo - Fe ao nivel de 30-.a) subunidade A, b) subunidade B.
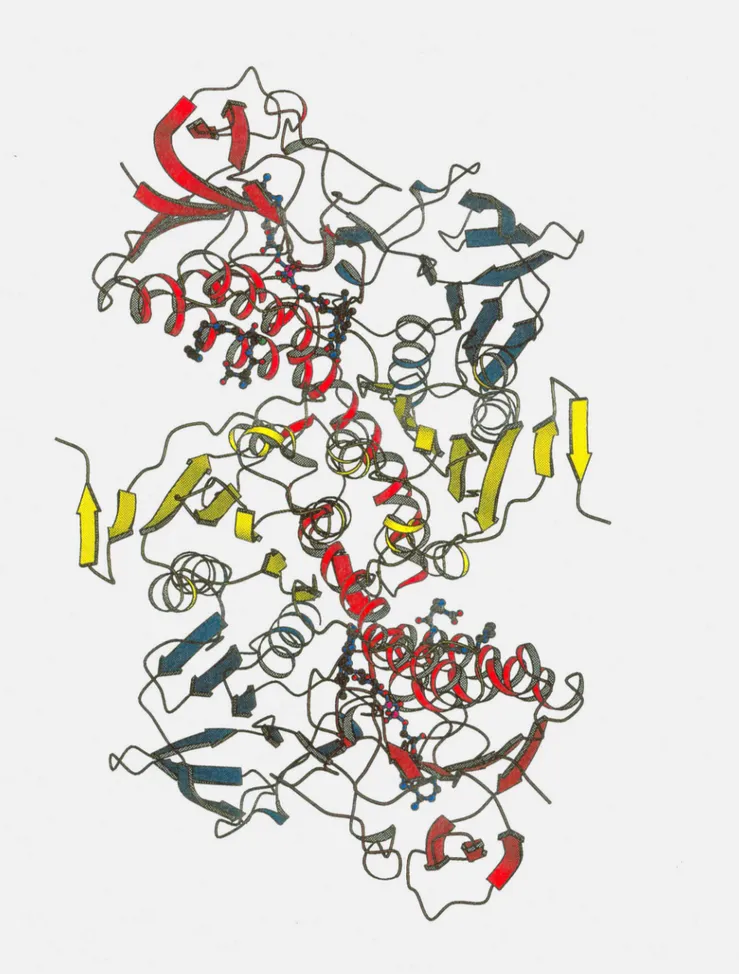
Estrutura tridimensional para C58S TR + Gspd. Em vermelho subunidade A, em azul subunidade B e em amarelo subunidade C.
Sequencia de aminoacidos para TR de T. c ru z i e a correspondente distribuic;ao quanta a estrutura secundaria.
Sitio ativo A para C58S TR + Gspd incluindo residuos fundamentais para a catalise eo ligante Gspd.
Superposic;ao entre os sitios ativos de TR de T. c ru z ; e Gr humana mostrando os residuos cataliticos para as duas enzima
Distribuic;ao eletrostatica para os sitios ativos do complexo mutante C58STR+GSPd de T.c ru z ie GR + GSSG humana.
Estrutura molecular para 0 FAD e 0 NADP+, nucleotideos envolvidos na ac;ao catalitica das enzimas Tripanotiona redutase e Glutationa redutase
Oiagrama esquematico da a9flo da fosfolipase A2 sobre 0 fosfolipideo. Extraido e adaptado de Voet & Voet (1995).
Empacotamento para as moleculas de BM-PLA2 na unidade assimetrica.
Alinhamento sequencial para 10 diferentes fosfolipases: B o t h r o p s a s p e r , pancreatica de porco, N a j a n a j a n a j a , A g k i s t r o d o n , C r o t a l u s a t r o x , Abelha, N a j a n a j a a t r a , B o t h r o p s J a r a r a c u s s u , pancreatica
bovina e humana.
Eletroforese em SOS-PAGE para 0 material enviado (coluna A) e as duas amostras finais depois do procedimento de puifica980 (B e C)
Perfil cromatografico ao longo das fra90es eluidas da coluna Superdex-75. Amostra inicial a 2 m g / m l
Porcentagem de atividade hemolitica para BM-PLA2 sendo que 100% corresponde
a
hem61ise total das celulas.Alinhamento parcial da sequencia N-terminal entre fosfolipases A2
C r o t a l u s A t r o x e B o t h r o p s m o o j e n i I Niveis de confian9a: A= alta,[6: =
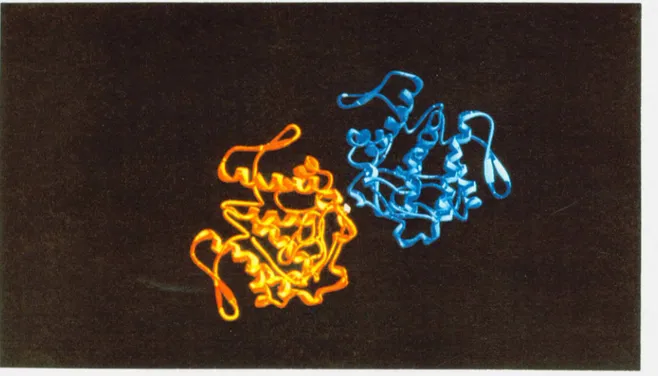
Dimero para fosfolipase A2 de B o t h r o p s m o o j e n i encontrada em sua forma cristalina.
Densidade eletronica para a ponte dissulfeto entre cisteinas 57 e 81. Mapa de densidade ao nivel 1 .0 c r
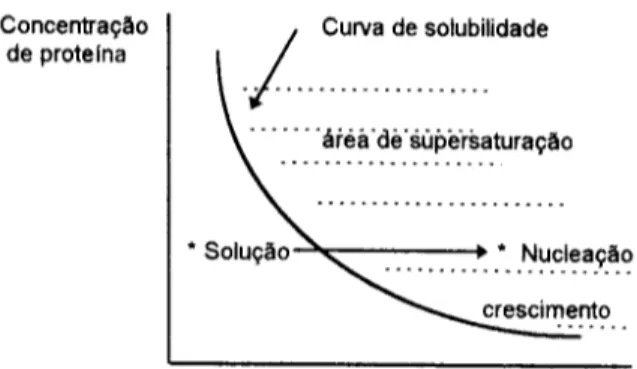
Tipica curva de solubilidade para uma proteina, como uma fun~o da concentra~o de sal ou qualquer outro parametro. Para melhores resultados a proteina deveria ser subrnetida a urn baixo nivel de supersatura~o que0 necessario para a forma~o do nucleo.
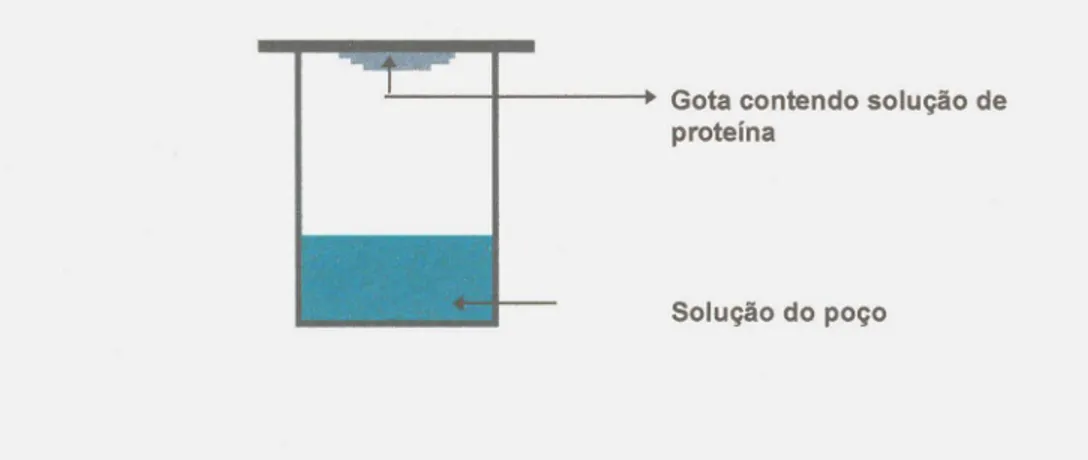
Gota suspensa ("hanging drop") .Tecnica de cristaliza~o de proteinas pelo metoda de difusao de vapor
Diagrama de Harker ilustrando a ambiguidade das fases para um unico derivado
Exemplo de correl;oes ao fator de espalhamento devido ao espalhamento anomalo ( curvas A e B). C corresponde a um oscilador harmonico simples.
Diagrama de Argand mostrando as difrentes contribuil;oes para os fatores de estrutura dos espalhadores anomalos P, normais Q e suas
CAGC
C a B P s
HLH
CACY
CAPL
R M N
FPLC
P D B
S IR
M IR
M R
MAD
PLAz B M - P L Az
NCS
T R
G R
R B R
GSH
GSSG
T [ S ) z
T[SH]z
DNA
C58STR
+GSPd
NADPH
FAD
A B R E V I A T U R A S
- Calgranulina C
- Protefnas que Iigam calcio (Calcium Binding Proteins)
- Helice-"Loop"-Helice
- Calciclina
- Proteina que liga calcio de placenta humana
- Ressonancia magnetica nuclear
- Fast Performance Liquid Cromatography
- Banco de dados de protefnas (Protein Data Bank)
- Substituil;:aoisom6rfica simples (Simple Isomorphous Replacement)
- SUbstituil;:ao isom6rfica multipla (Multiple Isomorphous Replacement)
- Substituil;:ao molecular (Molecular Replacement)
- Dispersao anomala a multiplos comprimentos de onda (Multiple
Anomalous Dispersion)
- Fosfolipases do tipo A2
- Fosfolipases A2 extraida do veneno da serpente B o t h r o p s
m o o j e n i
- Simetria nao cristalografica ( Non Crystallographic Symmetry)
- Tripanotiona redutase
- Glutationa redutase
- Refinamento de corpo rlgido (Rigid Body Refinement)
- T r y p a n o s o m a c r u z i
- glutationa reduzida
- glutationa dissulfeto
- tripanotiona reduzida
- tripanotiona dissulfeto
- Acido desoxirribonucleico (deoxyribonucleic acid)
-Tripanotiona redutase com sua cisteina 58 substitulda por uma serina
complexada com glutationil-espermidina
- Nicotinamida adenosina dinucleotfdeo fosfato
RESUMO
A cristalografia de raios-X e urn metodo de singular importancia para a determinayao da
estrutura de macromoleculas. A importancia em resolver estruturas de proteinas
continua a crescer em campos variando desde a bioquimica e biofisica basicas ate 0
desenvolvimento farmaceutico e biotecnologia.
o
presente trabalho esta relacionado com os estudos cristalograficos de tres diferentesmoleculas biol6gicas.
Calgranulina C de granul6citos porcinos, uma proteina que liga calcio, supostamente
envolvida em processos celulares regulados, foi cristalizada com parametros de rede
a=b=54.35 e c=141.32A, grupo espacial P3121 ou seu enantiomorfo P3221. Varias
tentativas foram feitas no sentido de se determinar a estrutura por substituiyao
molecular, mas a alta flexibilidade entre os motivos "EF-hands" quando da ligayao ao
ion calcio podem ser responsaveis pelo insucesso dos resultados.
Tripanotiona redutase de T . c r u z i e urn exelente alvo para modelagem de inibidores e
potenciais drogas contra a doenya de Chagas. 0 complexo mutante TR + GSPD foi
cristalizado com parametros de rede a=b=92.7 e c=156.2A, grupo espacial P43 . Urn
conjunto preliminar de fases foi obtido por refinamento de corpo rigido contra as
coordenadas da TR nativa. 0 modelo foi refinado a 2.4A de resoluyao, Rtactor=19.8%.
Fosfolipase A2extraida do veneno da serpente B o th r o p s m o o je n i foi cristalizada
com parametres de rede a=63.1, b=90.5 e c=40.2A e f3=125.10 , grupo espacial C2. A
estrutura foi resolvida por substituiyao molecular usando0 dimero da fosfolipase A2 de
C r o ta lu s a tr o x como modelo. A analise de sua sequencia de aminoacidos e necessaria
A B S T R A C T
X-ray crystallography is a essential method for determination of the structure of
macromolecules. The importance of solVing protein structures continues to grow in
fields ranging from basic biochemistry and biophysics to pharmaceutical development
and biotechnology.
The current work is concerned to the crystallographic studies of three different biological
molecules.
Calgranulin C from pig granUlocytes, a calcium binding protein though to be involved in
regulation of cell process, was crystallized with cell parameters a=b=54.35 and
c=141.32A, space group P3121 or its enantiomorph P3221. Several attempts were made
in order to solve the structure, but the high flexibility between its EF-hands motives while
binding the ion calcium may be responsible for the lack of success in the results.
Trypanothione reductase (TR) from T . c r u z ; is a target enzyme for modeling an inhibitor
for Chagas' disease. The C58S TR + GSPD mutant complex was crystallized with
a=b=92.7 and c=156.2A, space group P43 .A preliminary set of phases was obtained by
rigid body refinement against the coordinates for the native TR. The model was refined
to 2.4A resolution, Rfactor=19.8%
Phospholipase A2 extracted from the venon of the snake B o th r o p s m o o je n ; was
crystallized with cell parameters a=63.1, b=90.5, c=40.2A and B = 1 2 5 .1 ° , space group
C2. The structure was solved by molecular replacement techniques using the dimer of
phospholipase A2 from C r o ta lu s a tr o x as a model. The aminoacid sequence analysis is
~
C A P iT U L O
I
C A P IT u L O I -lN T R O D U I;:A O 2
papel, interrompendo ciclos metab61icos essenciais que podem levar
a
morte do
parasita.
No caso de doen9as vir6ticas, um exemplo e
0mecanismo com que
0virus
I n f l u e n z a ,
responsavel pela gripe, ataca a celula-alvo. Conhecemos hoje em dia
como e quais proteinas virais (neuraminidase e hemaglutinina) reconhecem os
receptores (acido sialico) das celulas, no caso do sistema respirat6rio. Foi entao
modelado, a partir de sua estrutura, um inibidor especifico para a neuraminidase, de
forma que, esta agora, nao e mais capaz de reconhecer estas celulas. Este suposto
inibidor ja esta em fase de testes farmacol6gicos (von Itzstein,
e t a l . 1 9 9 3 )Mas como estaria a cristalografia auxiliando no entendimento destes sistemas
biol6gicos tao complexos?
A resposta e que estrutura e fun9ao estao intimamente correlacionados. Os
segredos dos mecanismos de reconhecimento e resposta das macromoleculas
biol6gicas dependem de suas estruturas quimicas, e com
0mesmo grau de
importancia, de suas estruturas tridimensionais.
Alem da importancia biol6gica,
0sucesso da tecnica tem crescido com
0desenvolvimento do poder computacional. 0 cristal6grafo conta atualmente com
computadores de alta qualidade grafica e de processamento.
A estrutura cristalogratica de uma molecula biol6gica pode ser obtida atraves
da analise do padrao de difra9ao produzida por seus cristais. Esta analise pode ser
sumarizada nos seguintes passos:
'" Purifica9ao e cristaliza9ao
'" Coleta e processamento dos dados
'" Determina9ao da estrutura
4
n e 2
12 0 = 10 2 2 4 (l
+
c a s 2 8 )a to m s
F (h )
=
I
fje x p (2 7 tih . Xj) jo n d e F(H)
e
0 f a t o r d e e s t r u t u r a d a r e f le x a o h = (h, k, 1), fje x
j s a c 0 f a t o r d e1 "
-p(x) = - L ..J F (h )e x p [-2 7 ti(h .x
+
ky+
lz )]v h
m e d id a s , l(H) ex IF(H)1 2
C A P iT u L O I -IN T R O D U ~ A O 5
se mostrado eficaz nao s6 par minimizar os danos causados pela exposi9ao aos
raios-X, mas tambem por tomar possivel
0estudo de cristais que nao sac estaveis
na presenl;a do Iiquido-mae (solul;ao na qual
0cristaI cresceu).
A fonte de radial;ao utilizada tambem e urn fator importante na coleta de
dados. Fontes de alta intensidade, como fontes de radia9ao sincrotron sac hoje
amplamente
utilizadas
em Cristalografia de
Raios-X,
0que tern
permitido
experimentos bastante avanl;ados. Devido
a
sua alta intensidade,
a radial;ao
sincrotron tern side utilizada para otimizar a coleta de dados e permitir estudos de
amostras (cristais pequenos
e / o ucela unitarias grandes) que nao poderiam ser
realizados em fontes convencionais.
Outra aplical;ao e a cristalografia resolvida no tempo, que utiliza a distribui9ao
espectral continua da radia9ao sincroton na analise da estrutura ao longo do tempo
atraves de experimentos de difral;ao de Laue. Nestes experimentos e possivel
acompanhar rea90es enzimaticas e processos fotoquimicos.
0terceiro maior usa da
radial;ao sincrotron em cristalografia de raios-X e a determina9ao de estruturas
cristalinas diretamente das medidas de difral;ao an6mala realizadas a diferentes
comprimentos de onda.
Em todas as fontes de radial;ao sincrotron espalhadas pelo mundo, ha hoje
linhas dedicadas exclusivamente
a
cristalografia de proteinas. No sincrotron
brasileiro, onde as primeiras linhas de pesquisa ja estao em fase de testes, esta
sendo construida uma linha destinada a coleta de dados em monocristais de
proteinas.
Varios sac os esforl;os para minimizar as erros durante a coleta de dados,
que podem levar a problemas posteriores na interpreta9ao dos resultados. No
CAPtruLO I -INTRODUc;itO
o m e t o d o d a s u b s t it u ic ; a o m o le c u la r (~7ft,NI)~qEIQ ). E s t e m e t o d o e b a s e a d o n o
C A P IT u L O I-lN T R O D U C ;:is.O
C A P IT u L O I-lN T R O D m ;;Js.O 8
descrito no apemdice E , a determinac;aoda posic;aodestes atomos pesados podem
levar a obtenc;aode um conjunto preliminar de fases.
o
terceiro e ultimo metoda e
0chamado metodo da dispersao anomala a
multiplos comprimentos de onda (MAD). Quando os eletrons se encontram em
camadas mais internas do atomo, isto e, mais pr6ximos do nucleo,
0tratamento
dado ao espalhamento devido a estes eletrons deve ser mais cuidadoso, uma vez
que, a forc;a de interac;ao entre esses e
0nucleo atomico nao pode mais ser
desprezada e
0eletron nao pode mais ser tratado como livre. Devido
a
forc;a de
interac;ao com
0nucleo, existe a introduc;ao de um termo imaginario ao fator de
espalhamento atomico, dependente do comprimento de onda da radiac;ao.
Um modelo inicial construido a partir de uma conjunto preliminar de fases
usualmente contem muitos erros. No sentido de se produzir um modelo com maior
acuracia, e necessario introduzir
0refinamento cristalogratico tao bem quanta a
reconstruc;ao do modelo atraves de manipulac;oes graticas. Estes passos sac
introduzidos em um processo ciclico de melhora gradual do modelo. Programas de
refinamento modificam
0modelo com
0objetivo de melhorar a concordancia entre
as amplitudes dos fatores de estrutura observados e calculados. Devido
a
limitada
resoluc;ao tipicamente obtida em cristalografia biomolecular, a relativa escassez de
dados esperimentais e suprida por informac;oes quimicas, como comprimentos de
ligac;ao e angulos. A reconstruc;ao do modelo manualmente e necessario para
remover erros que nao tem como ser corrigidos pelos programas de refinamento
(A P E N D lC E 6 .).
Quando
a estrutura
cristalografica e submetida
a
analise, residuos
importantes
para
a
catalise,
contatos
intermoleculares
responsaveis
pela
C A P IT u L O I -IN T R O D U C A O 9
podem ser identificadas, mas e importante ressaltar aqui, que a estrutura
cristalografica e uma "foto"da molecula em sua conformac;ao media, e portanto
cautela deve ser tomada para que nao haja uma interpretac;ao errenea dos
resultados.
o
presente trabalho visa 0 estudo cristalografico de tres diferentes moleculasbiol6gicas. No C A P iT U L O II sac apresentados os resultados dos estudos sobre a
calgranulina C de granul6citos porcinos e as tentativas no sentido de se determinar
a sua estrutura. 0 C A P iT U L O III apresenta 0 estudo cristalografico do complexo
mutante C58S Tripanotiona redutase + GSPd de
T.
c r u z i , trabalho este realizadodurante periodo de doutoramento sanduiche na Universidade de Manchester,
Inglaterra, sob a orientac;ao do Prof. Dr. Willian N. Hunter. A enzima Tripanotiona
redutase e um excelente alvo para modelagem racional de inibidores, na busca de
um farmaco contra a doenc;a de Chagas. 0 estudo deste complexo faz parte de um
projeto que visa uma caracterizac;ao meticulosa da enzima. No Q A P ,iT U lO IV
apresentamos os estudos cristalograticos da fosfolipase A2 extraida do veneno da
serpente B o t h r o p s m o o j e n i . Cristais de pequena dimensao foram obtidos, mas a
estrutura pede ser resolvida atraves do metodo da substituic;ao molecular. Os
sucesso da determinac;ao da estrutura e mais um novo resultado quando da
discussao do comportamento destas enzimas durante seu mecanisme de atividade.
Nos apendices (A -< il) se encontra em maior detalhe a teoria relativa
a
determinac;ao da estrutura por cristalografia de raios-X, como uma base para
!!/'
C A P i T u L O
I I
C A L 6 R A N U L IN A
C
D £ 6 R l\N U L O C IT O S
P O R C l N O S
1 1 .1 .IN T R O D U C A O
desde a biomineralizaC;8o em ossos e dentes ate um papel complexo como mensageiro
F i g u r a 1 1 - 1 D i a g r a m a e s q u e m a t i c o d o m o t i v o " E F - h a n d " d e l i g a ~ a o d e c a l c i o ( a ) s i m b o l i z a d o p e l a
m a o d i r e i t a ( b ) .
A f u n 9 8 0 d a c a l m o d u l i n a e m t r a n s d u 9 8 0 d e s i n a i s d e C a2+ t e m s i d e e x t e n s i v a m e n t e
q u e e s t a f o r m a d e i n t e r a 9 8 0 p o d e r e p r e s e n t a r u m m e c a n i s m e g e r a l p a r a a f u n 9 8 0 d e s t a s
p r o t e f n a s c o m o m e d i a d o r e s d e s i n a i s d e C a2+.
C A P i T u L o 1 1 - C A L 6 R . A N U L I N A C D E 6 R . A N U L O C r r O S P O R C I N O S 1 2
(Ernst et a I, 1990), proteina de aproximadamente 33kDa, e a grancalcina (Boyhan et a I,
1992) de 28kDa.
Alem destas, granul6citos tambem expressam calmodulina (Jones e t a I, 1982), e
varias CaBPs do tipo 8100, como calgranulinas A e B, supostamente associadas em um
heterocomplexo (Dell'Angelica e t a I, 1996) e a calgranulina C (Dell'Angelicae t a I, 1 9 9 4 ) .
A familia 8100 compreende um grupo de proteinas ligantes de calcio de baixo peso
molecular (10-12kDa) (Kligman&Hilt,1988). Uma marcante caracteristica estrutural da
familia das 8100 € I a presen~ de dois sitios de ligac;aode calcio (Ca2+) na conformac;ao
dos classicos motivos liEF-hands", com 0 N-terminal mostrando um nao usual "loop" de
ligac;aode calcio contendo 14 residuos (Heizmann e Hunziker, 1991).
A func;ao destas CaBPs do tipo 8100 nao sac conhecidas. Algumas protefnas
como a CACY (calciclina) e a CAPL (proteina que liga calcio de placenta humana) foram
associadas ao desenvolvimento de tumores e
a
induc;aode metastase (Muraoe t a I, 1989),quando sua expressao esta desregulada.
Dois membros da familia 8100 tiveram suas estruturas tridimensionais
determinadas em soluc;ao por tecnicas de RMN (ressonancia magnetica nuclear): ,
calbindina, tambem conhecida como D9K (Kordel e t a I, 1993) e a calciclina, conhecida
como CACY ( Potts e t a I, 1995). Em 1986 a estrutura da D9K foi tambem determinada por
cristalografia de raios-X (8zebenyi e Moffat, 1986). Em todos os casos, as proteinas nao
tinham 0 ion calcio ligado. As proteinas se enovelam em um dominio globular unico
lembrando 0 dominio de ligayao de calcio de calmodulina e troponina C. Contudo, apesar
da alta identidade em termos de estrutura primaria e similaridades estruturais. existem
algumas diferen~s funcionais importantes entre os membros da familia 8100. Calbindina
D9K, por exemplo, e uma proteina 8100 atipica, ja que naG sofre variayoes
conformacionais significantes induzidas por calcio. Isto sugere que D9K pode funcionar
p-CAGC h- CAGB h-CAGA p-CABP9K b-S100~ h-S100D h-CAPL b-S100L h-S100E h-CACY b-S100a h-S100P p-Ca[l] p-S100C p-CAGC h- CAGB h-CAGA p-CABP9K b-S100~ h-S100D h-CAPL b-S100L h-S100E h-CACY b-S100a h-S100P p-Ca[l] p-S100C ... TKLEDHLEGIINIFHQYSVRLGHYDTLIKRELKQLITKELPNTLK ... MTCKMSQLERNIETIINTFHQYSVKLGHPDTLNQGEFKELVRKDLQNFLK ... MLTELEKALNSIIDVYHKYSLIKGNFHAVYRDDLKKLLETECPQYIR ... MSAQKSPAELKSIFEKYAAKEGDPNQLSKEELKQLIQAEFPSLLK ... SELEKAVVALIDVFHQYSGREGDKHKLKKSELKELINNELSHFL. MPAAWILWAHSHSELHTVMETPLEKALTTMVTTFHKYSGEEGSKLTLSRKELKELIKKEL ..CL. ...• MACPLEKALDVMVSTFHKYSGKEGDKFKLNKSELKELLTRELPSFL. ... SSPLEQALAVMVATFHKYSGQEGDKFKLSKGEMKELLHKELPSFV. ... MARPLEQAVAAIVSTFQEYAGRCGDKYKLCQAELKEKKQKELATWT.
... MACPLDQAIGLLVAIFHKYSGREGDKHYLSKKELKELIQKELTI ...
... GSELETAMETLINVFHAHSGKEGDKYKLSKKELKELLQTELSGFL.
... MTELETAMGMIIDVFSRYSGSEGSTQTLTKGELKVLMEKELPGFL.
... PSQMEHAMETMMFTFHKFAGDKGY ...LTKEDLRVLMEKEFPGFL.
... MAKRPTETERCIESLIAIFQKHAGRDGNNTKISKTEFLIFMNTELAAFT.
N.TKDQGTIDKIFQNLDANQDEQVSFKEFVVLVTDVLITAHDNIHKE .
KENKNEKVIEHIMEDLDTNADKQLSFEEFIMLMARLTWASHEKMHE-GDEGPGHHHKPGLGEGTP
... KKGADVWFKELDINTDGAVNFQEFLILVIKMGVAAHKKSHEESHKE .
GP RTLDDLFQELDKNGNGEVSFEEFQVLVKKISQ .
EEIKEQEVVDKVMETLDSDGDGECDFQEFMAFVAMITTACHEFFEHE . GE.MKESSIDDLMKSLDKNSDQEIDFKEYSVFLTMLCMAYNDFFLEDNK . GKRTDEAAFQKLMSNLDSNRDNEVDFQEYCVFLSCIAMMCNEFFEGFPDKQPRKK . GEKVDEEGLKKLMGDLDENSDQQVDFQEYAVFLALITIMGNDFFQGSPARS . PTEFRECDYNKFMSVLDTNKDCEVDFVEYVRSLACLCLYCHEYFKDCPSEPPCSQ . GSKLQDAEIARLMEDLDRNKDQEVNFQEYVTFLGALALIYNEALKG . DAQKDADAVDKVMKELDEDGDGEVDFQEYVVLVAALTVACNNFFWENS . QSGKDKDAVDKLLKDLDANGDAQVDFSEFIVFVAAITSACHKYFEKAGLK . ENQKDPLAVDKIMKDLDQCRDGKVGFQSFFSLIAGLTIACNDYFVVHMKQKGKK . QNQKDPGVLDRMMKKLDLDSDGQLDFQEFLNLIGGLAIACHDSFIKSTQK .
Figura 11-3.Alinhamento sequencial entre proteinas que ligam calcio do tipo 5100 usando 0 sistema
de Darwin (Gonnet, G.H., 1992).p - C A G C - Calgranulina C porcina, h - C A G B - Calgranulina B humana,
h-CAGA-ealgranulina A humana, p - C a B P 9 K - Calbindina-D9k porcina, b - S 1 0 0 P -Cadeia ~ da proteina
5100 bovina, h - C A C L - proteina que liga calcio de placenta humana, b - S 1 0 0 L - 5100L bovina, h
-C A -C Y - Calciclina humana,b - S 1 0 0 a .- Cadeia ada proteina 5100 humana,h - S 1 0 0 P - 5100P humana,p -C a [1 ]- Cadeia leve da calpactina I p o r c in a ,h - S 1 0 0 D -51000 humana, h - S 1 0 0 E - 5100E humana, p
-S 1 0 0 C - 5100C porcina.
Experimentos quanta as propriedades de liga<;ao ao Ca2
CAPITuLo 11- CAL6R.ANULINA C DE 6R.ANULOClTOS PORCINOS 1 5
propriedades da proteina ao se ligar ao ion calcio. Na ausElnciade Zn
2+ ,CAGC se liga a
um ion Ca
2+
por monamero, enquanto que na presenc;ade Zn
2+CAGC se liga a 2 ions
Ca
2+
por monamero, 0 que e consistente com a presenc;a dos dois motivos liEF-hands"
observados atraves da analise da sequencia de aminoacidos.
Calgranulina C tambem mostra alta afinidade por zinco. Como especula980
podemos notar que a regiao C-terminal (Figura 11-2)possui um motivo His-X-X-X-His
compreendendo os residuos 85 a 89. Estas duas histidinas fariam parte de uma helice em
uma conforma980 tal que favoreceria a liga980 com 0 zinco.
Varias sac as questoes que restam ser elucidadas.
/n v iv oCAGC se oligomeriza
como um dimero? A Iigayao ao ion Ca
2+da Calgranulina C e regulada pelo ion zinco?
Qual 0 alva celular para CAGC? Como seria esta varia980 conformacional na presenya
dos ions calcio e zinco? E, qual seria a fun980 da Calgranulina C
em granul6citos
A estrutrura cristalografica tem papel fundamental para ajudar a elucidar estas
questoes.
Cristais de boa qualidade foram obtidos, mas ate 0 presente CAGC nao teve sua
estrutura determinada.
No decorrer do capitulo sera descrito todo 0 trabalho realizado na tentativa de se
determinar a estrutura cristalografica da Calgranulina C de granul6citos porcinos.
Os resultados obtidos foram submetidos e aceitos para publica980 (Nonato
e t a I,1 1 .2 .P U R IF IC A C A O
E C R IS T A L IZ A C A O
A purifica~o da CAGC (Dell'Angelica e t a I, 1994) 13 feita inicialmente isolando os
granul6citos do sangue fresco por sedimenta~o de Perxan seguida por centrifuga~o por
gradiente de Percoll (Schleicher e t a I, 1993). As celulas extrafdas sac rompidas e
fracionadas em uma coluna de filtra~o em gel Sephadex G-75. CAGC 13 a componente
mais abundante encontrada nas fra90es de 10 a 15 kDa, que ap6s combinadas, sac
levadas a uma cromatografia por troca ienica em uma coluna MonoQ (Pharmacia) em
FPLC (Pharmacia).
Para os experimentos iniciais de cristaliza9ao, amostras de CAGC purificada na
forma liofilizada foram-nos enviadas pelo Instituto de Qufmica y Fisicoqufmica da
Universidade de Buenos Aires, Argentina, atraves de uma colabora~o com 0 pesquisador
Christian Schleichler.
Como procedimento padrao, os primeiros experimentos de cristaliza~o foram
realizados no sentido de se encontrar condi90es de cristaliza9ao que estejam pr6ximas
das condi90es consideradas ideais, isto 13, condi90es que levem ao crescimento de cristais
adequados para os experimentos de difra9ao.
Para otimizar esta busca, inicialmente 13 feita uma estimativa do ponto de
precipita~o da protefna para diferentes agentes precipitantes, como sulfato de amenia e
polietilenoglicol, dentre outros. Uma vez que a cristaliza9ao, como descrito no APENDlCE.
A , 1 3um estado metaestavel entre a proteina em solu9ao e precipitada, as primeiras caixas
de cristaliza9ao sao montadas variando a concentra9ao do precipitante em tome dos
pontos de precipita9ao previamente estimados. Outro parametro variado 13 0 pH,
Tabela 11-1Ponto de precipita~Ao para a Calgranulina C para diferentes agentes precipitantes. 0 teste foi realizado com uma solu~Ao de proteina a 10mglml em 1mM (j.~.~LJlf~t()c;t~;ziflc:().(?f1~g~)~.ti~p~~. ( q 8 t i '1 I t - J : l ( ) ~ S ) .C:()I1l().~()Iuy~otCll1lpA()CI.pti7. 0
Agente precipitante Formula qufmica Ponto de precipitayao estimado
sulfato de amenio IN§14]?§94 69%
T a b e l a 1 1 - 2C o l e t a d e d a d o s p a r a o s c r i s t a i s n a a u s e n c i a ( c r i s t a I 1 ) e p r e s e n ~ a( c r i s t a l 2 ) d e c a l c i o .
C r is t a l F o t o s n u m e r o d e f o t o s V a r ia ~ a o a n g u la r e n t r e a s t e m p o d e e x p o s i~ o f o t o s
1 S tili 2 9 00
3 0 m in
O s c ila c a o 4 5 2 ° 5 0 m in
~ S till 2 9 00 1 5 m in
O s c ila c a o 6 0 1 .50 3 0 m in
T a b e l a 1 1 - 3- E s t a t i s t i c a d a c o l e t a d e d a d o s p a r a c r i s t a i s d e C a l g r a n u l i n a C a 2 . 6
A
d e r e s o lu ~ a o .C r i s t a i s# 1 e # 2 f o r a m c r e s c i d o s n a a u s e n c i a e n a p r e s e n ~ ad o s i o n s C a2+, r e s p e c t i v a m e n t e .
R e s o ~ o
( A )
R e fle x o e s
m e d . in d o
R m e r g e C o m p le te z a
1 > 3 0
R e fle x o e s
m e d . in d o
R m e < g e C o m p le te z a
1 > 3 0
8.00 1294 306 0.026 97.7 1103 297 0.046 98.3 0.193
5.70 2026 500 0.043 96.4 2288 525 0.042 98.3 0.025
4.67 2078 586 0.039 98.6 2542 616 0.042 98.7 0.031
4.05 2083 699 0.036 97.4 2768 732 0.040 97.8 0.038
3.62 1980 757 0.042 96.4 2911 806 0.042 98.6 0.039
3.31 2008 829 0.047 94.9 2993 888 0.045 98.0 0.040
3.06 2000 866 0.058 91. 4 3022 937 0.047 96.7 0.043
2.87 1865 857 0.078 86.7 2922 924 0.054 96.5 0.055
2.70 2068 940 0.097 83.3 3099 1027 0.058 94.8 0.064
2.60 2046 898 0.125 77 .9 3019 991 0.068 92.8 0.075
CAPITuLO I I •CAL6RANULlNA C D£ 6RANULOCITOS PORClNOS 19
P a r a m in im iz a r o s d a n o s c a u s a d o s p e la r a d ia l;8 o , a s c o le ta s fo r a m r e a liz a d a s
m a n te n d o o s c r is ta is s o b u m flu x o d e n itr o g e n io g a s o s o c o m te m p e r a tu r a c o n tr o la d a d e
4 ° C . D e s ta fo r m a , u m u n ic o c r is ta I d e c a d a d ife r e n te c o n d il;8 o d e c r is ta liz a l;8 o s e m o s tr o u
s u fic ie n te p a r a fo r n e c e r c o n ju n to s d e d a d o s d e b o a c o m p le te z a .
A s fo to s " s till" ( 0 e 9 0 ° , fo to s e m q u e 0 c r is ta l s e m a n te m p a r a d o ) fo r a m u tiliz a d a s
p a r a a c a r a c te r iz a l;8 o in ic ia l d o s c r is ta is . C r is ta is 1 e 2 s e m o s tr a r a m is o m 6 r fic o s .
A C A G C s e c r is ta liz a c o m p a r a m e tr o s d e r e d e a = b = 5 4 .3 5 ( 2 ) e c = 1 4 1 .3 2 ( 5 ) A . A
p r e s e n 9 8 d o lo n g o e ix o c fo i 0fa to r d e te r m in a n te p a r a a r e s o lu c ;a o d o s d a d o s c o le ta d o s , 2 .6 A .
E m b o r a o s c r is ta is d ifr a ta s s e m b e m , a m a is a lta r e s o lu c ;a o o s p ic o s s e s u p e r p u n h a m e
m e s m o u tiliz a n d o 0 m o d o m a is s e n s iv e l d e le itu r a d a s p la c a s d e im a g e m ( 1 0 0 J lm p o r p ix e l)
n a o e r a p a s s iv e l d ife r e n c ia - Io s .
A p r o m e d ia c ;a o d a s r e fle x 5 e s e q u iv a le n te s d e fin e u n ic a m e n te a s im e tr ia d e L a u e
c o m o 3 m 1 . A a n a lis e d a s e x tin g 6 e s s is te m a tic a s in d ic a a c o n d ic ;a o d e e x is te n c ia p a r a
r e fle x 5 e s d o tip o [= 3 n , 0 q u e e c o m p a tiv e l c o m 0 g r u p o e s p a c ia l P 312 1 o u s e u e n a n tio m o r fo
P 322 1 .
11.4.
S I M E T R I A N A O C R I S T A L O G R A F I C AA p a r tir d o s d a d o s c r is ta lo g r a fic o s fo i e n ta o p o s s iv e l fa z e r u m a e s tim a tiv a d o n u m e r o
d e m o n 6 m e r o s p o r c e la u n ita r ia e d o c o n te u d o d o s o lv e n te . ( A P E N D IC E B ) . A T a b e la 1 1 - 4
m o s tr a o s v a lo r e s e s tim a d o s p a r a a d e n s id a d e d o c r is ta l ( D c ) e a p o r c e n ta g e m d e s o lv e n te
(VvN) e m fu n c ;a o d o n u m e r o d e m o le c u la s d e p r o te fn a ( n )
T a b e la 1 1 - 4C a lc u lo d o n u m e r o d e m o h flc u la s d e C A G C p o r c e la u n ita r ia
D c ( g /c m3)
1 .0 7
1 .1 5
1 .2 3
VwN
7 8 .4
5 6 .7
3 5 .1
n ° d e M a th e w s ( J .h D a )
5.68
2 .8 4
1 .8 9
U m m o n 6 m e r o n a u n id a d e a s s im e tr ic a c o r r e s p o n d e r ia a u m a a lta p o r c e n ta g e m d e
s o lv e n te , 0 q u e e in c o m u m p a r a c r is ta is q u e d ifr a ta m b e m c o m o o s d e c a lg r a n u lin a C . O s
v a lo r e s e n c o n tr a d o s p a r a 0 d im e r o e tr im e r o s e e n c o n tr a m d e n tr o d e u m a fa ix a d e v a lo r e s
e le m e n to s d e s im e tr ia n a o c r is ta lo g r a fic a fo i e n ta o c a lc u la d a a fu n t;8 o d e a u to - r o ta t;8 o ( F ig u r a
1 1 - 4 ) u tiliz a n d o 0 p r o g r a m a A M o R e ( N a v a z a , 1 9 9 4 ; A p e n d ic e D )
F ig u r a 1 1-4 P r o je ~ A o e s te r e o g r a fic a a o lo n g o d o e ix o c , s e A o p o la r k = 1 8 0 ° , d a fu n ~ A o d e a u to - r o ta ~ a o
c a lc u la d a s c o m d a d o s d o c r is ta l1 n o in te r v a lo d e 2 .6 - 1 5 A.
11.5.
TENTATIVAS
PARA A DETERMINACAO
DA ESTRUTURA
o
m e to d a d a s u b s titu i9 a o m o le c u la r , im p le m e n ta d o n o p r o g r a m a A M o R e ( N a v a z a ,1 9 9 4 ) , fo i e x a u s tiv a m e n te u tiliz a d o n a te n ta tiv a d e s e d e te r m in a r a e s tr u tu r a d a
Tabela 11-5Lista de proteinas extraidas do banco de dados de coordenadas de proteinas (PDB) que apresentam 0 motivo "EF-hand"( helice-loop-he lice).
Protefnas que ligam calcio
calmodulina parvalbumina
modelos (Protein Data Bank)
1c11,1deg, 1trc, 10sa, 1clm, 1cln,2bbm, 3cln 1cdp,4cpv,5cpv,2pa1 ,4pa1, 1pa1,3pa1 ,3pat,5pa
1,1 rtp 1rro, 10md
4icb, 3icb,4bod, 2bca, 2bcb, 1cb1 1top,4tnc, 1cta, 1ctd, 5tnc
2sep,2sas 1rec 1scm Oncomodulina
Calbindina D9K troponina C
protefna que liga calcio do sarcoplasma recoverina
11.5.2Substituicao
isom6rfica
(SIRIMIR)
Tabelall:6e,:()m p0rtam ento dos cril;tail;~iarltedal;solu~oesdE!atom os pel;Cldostestadas.
Composto concentr8C;8o obserV8c;6es
Pt(NH3h C b 1 mM se dissolveu ap6s 24h de banho
(CH3COzhSm.xH20 1mM se dissolveu ap6s 24h de banho
(CzH30z)2Hg 1mM se dissolveu ap6s 24h de banho
U02(CzH30Z)2+2H20 1mM quebrou imediatamente ap6s ser mergulhado na soluyao
3CdS04.8H20 1mM se dissolveu ap6s 24h de banho
C A P I T u L O I I-C A L 6 R A N U U N A C D E 6 R A N U L O C r r O S P O R C I N O S 2 3
o
cristal banhado na solu980 de K2PtCI4 , unico que resistiu ao experimento, foilevado a coleta de dados. Os dados coletados para 0 possfvel derivado foram
promediados contra os dados da protefna nativa e mostraram uma pequena varia980 para
o valor do R m e r g e , de 5.6% para 7.4% , mudan98 esta muito pequena para a presen98 de
um derivado.
Nenhuma nova condi980 foi testada por falta de disponibilidade de amostra
purificada para cristaliza980
11.5.3Dispersae anomala a multiples cemprimentes de enda (MAD)
Outra tentativa foi a coleta de dados no sfncrotron em Hamburgo com 0 objetivo de
se determinar a estrutura pelo metodo de dispers80 anomala a mUltiplos comprimentos de
onda. 0 fon zinco foi utilizado nas condi90es de cristaliza980 e as fortes evidencias
bioqufmicas indicadas na introdu980 sugeriam que este pudesse estar ligado as moleculas
de protefna do crista!. Desta forma a aplica980 do metodo seria possfvel devido a
presen9a do zinco como espalhador anomalo.
Na tentativa de reduzir os danos causados por radia980, foram utilizadas tecnicas
em criocristalografia , onde os cristais sac banhados em uma solu98o criogenica e a coleta
para os varios cristais feita a aproximadamente -170°C.
Os cristais que haviam side transportados desde de 0 Laborat6rio de Cristalografia
de Proteinas do IFSC, se mostraram de ma qualidade comparados com os experimentos
realizados "em casal', isto
e,
perderam muito sua capacidade de difratar. N80 foi posslvelcoletar ao menos um unico conjunto completo de dados (aproxidamente 18 cristais foram
submetidos a coleta). Associamos este problema a varia980 de temperatura e a agitar;80
1 1 . 6 A N A L I S E D O S R E S U L T A D O S E P E R S P E C T I V A S F U T U R A S
Determinar a estrutura da Calgranulina C e sem duvida nenhuma muito importante
para
0entendimento do mecanisme de atuayao das proteinas que ligam calcio e sua
variayao conformacional na presenca do ion calcio. Os insucessos obtidos atraves do
metodo da substituiyao molecular e mais uma evidencia da liberdade conformacional
destas proteinas. A calgranulina C pode apresentar uma variayao conformacional quando
comparada com todas as proteinas que ligam calcio que ja tiveram suas estruturas
tridimensionais determinadas seja por cristalografia de raios-X ou por ressonancia
magnetica nuclear.
Cristais na presen98 e ausencia de calcio nas condi~oes de cristalizayao foram
obtidos. No sentido de se examinar possivel diferen98s entre os dois conjuntos de dados,
estes foram promediados (Tabela 11-3).A estatlstica resultante mostra diferen98s somente
a
baixa resoluyao (abaixo de
8A),que pode ser explicada devido ao espalhamento do
solvente originario de dois diferentes meios de cristalizayao. Este resultado sugere que os
dois cristais podem ser identicos. Considerando a alta afinidade da proteina em se ligar
aos ions Ca
2+
(Dell'Angelica et ai,
1994),e razoavel supor que os procedimentos de
purificayao nao removeram os ions de calcio Iigado
a
proteina. Alternativamente, a ligayao
ao calcio pode nao ser suficiente para produzir varia~oes conformacionais grandes
0suficiente para resultar em significantes diferen98s na intensidade entre os dois conjuntos
de dados.
Experimentos de purificayao ja foram iniciados em nosso proprio laboratorio. Desta
forma a grande quantidade de material exigida pode agora ser obtida com mais facilidade,
possibilitando experimentos como a obten~ao de derivados isomorficos, quando novas
1 1 . 7 .R E F E R E N C I A S
Boyhan, A, Casimir, C.M., French, J.K. Teahan, C.G., and Sega, A W.(1992)
J.
B io I. C h e r n .267,2928-2933
Cook, W.J., Ealick, S. E., Babu, Y.S., Cox, J.A & Kumar S. V., (1991), J. B io I. C h e r n ., vol.
266, pp. 652-656.
Dell'Angelica, E.C., Schleicher, C.H. and Santome J.A (1994)
J.
B io I. C h e r n ., 2 6 9 ,28929-28936.
Fohr, U.G., Heizmann, C.W., Engelkamp, D., Schafer, B. W. and Cox, J. A (1995)
J.
B io I.C h e r n . vol. 270, 36, 21056-21061.
Jones, H.P., Ghai, G., Petrone, W.F., and McCord, J. (1982) B io c h im . B io p h y s . A c ta 714,
C A P IT u L o II- C A L 6R A N U U N A C D E 6R A N U L O C rrO S P O R C IN O S
Kligman D. and Hilt D. (1988). T r e n d s B io c h e m . S c i., 13,437-443.
Nonato, M.C., Schleicher, C.H., Garrat, R.C., Santome, and Oliva G. Crystallization and
Preliminary Crystallographic Studies of Calgranulin C, a S100 like Calcium-binding Protein
from pig Granulocytes. Acta Cryst. D. (a c c e p te d )
Potts, B.C.M, Smith, J., Akke, Macke, T.J., Okazaki, K., Hiroyoshi, H., Case, D.A. and
Chazin, W. J. (1995). N a t.
Struc.
B io i. Vol 2, 9, 790-796.Schleicher, C.H., DellAngelica, E. C., and Santome, J.A. (1993) In t. J . B io c h e m . 25,
1251-1256
Skelton, N. J., Kordel, J., Akke, M., Forsen, S. and Chazin, W.J. (1994) N a tu r e S tr u c t. B io i. 1 ,
~
C A P i T u L O :
III
TRIPANOTIONA
REDUT
ASE
Df:
TRYPANOSOMA CRUZ!
o
parasita Trypanosomacruzi
(T.CruZI)
e um protozoario flagelado da Ordem doskinetoplastidas, Subordem tripanossomatida, agente causador da doen~ de Chagas, doen~
endemica na America do Sui e America Central. De acordo com as estatisticas (Moncayo,
1993; W orld Health Organization, 1991), 18 milhoes de pessoas estao infectadas com
T. cruz;
e aproxidamente 45 mil mortes ocorrem anualmente.
Os principais orgaos atacados pela doen~ de Chagas sac: 0 musculo cardiaco, 0
musculo Iiso do sistema digestivo e os glanglios, levando a uma hipertrofia do musculo
cardiaco ou alargamento do es6fago ou c6lon (mega sintomas). A quimioterapia disponivel
C A P IT uL o 111- T R IP A N O T IO N A R £D U T A S E D E ToCRUZl 28
efeitos colaterais, e e somente efetivo nos estagios precoces da doen98. Embora milhares de
drogas tenham side testadas contra este parasita, nenhuma ainda provou ser suficientemente
apta para uso gera!.
Recentes pesquisas sobre processos metab61icos basicos do parasita (Fairlamb, 1989)
e sua comparayao com a bioqufmica do hospedeiro tem apontado algumas importantes
diferen98s que podem ser utilizadas na busca de drogas antitripanocidais mais eficazes.
Um exemplo e 0 estudo do cicio metab6lico de tiol e poliaminas. A maioria, se neo
todas as celulas, contem altas concentray5es (milimolares) de ao menos um tiol de baixo peso
molecular. Na maioria dos organismos eucariotos a maior componente e a glutationa (GSH,
L-y-glutamil-L-cisteinil-glicina). GSH possui importantes papeis para 0 metabolismo celular,
incluindo funy5es regulat6rias, de proteyao e como cofator. Com 0 auxflio da enzima
glutationa redutase (GR) , GSH mantem 0 correto balan~ tiol-redox intracelular e tem sido
associado ao controle da sfntese de DNA e regulayao da atividade enzimatica. GSH tambem
serve para proteger a celula contra danos oxidativos atraves da procura de radicais livres e
pela remoyao enzimatica de per6xidos pelas peroxidases dependentes de glutationa e
glutationa-S-transferases. A ultima classe de enzimas tambem funciona como desintoxicante
de certos xenobi6ticos.
Embora tripanossomatfdeos e leishmanias contenham GSH, estes neo contem
glutationa redutase, que em outras celulas e a enzima responsavel em manter a glutationa
dissulfeto (GSSG) na sua forma reduzida (GSH, Figura 1II-1A; Ghisla & Massey, 1989). Ao
contrario, estes organismos reduzem GSSG e outros dissulfetos por intermeclio de uma troca
nao enzimatica tiol-dissulfeto com tripanotiona, um ditiol de baixo peso molecular unicamente
encontrado nestes organismos (Fairlamb, 1985). Tripanotiona (N1,N8_
bis[glutationil]esperimidina) e mantida dentro da celula como 0 ditiol (T[SHh), devido a a~o
A bioslntese da tripanotiona (Figura 111-2)vem da glutationa (tiol) e espermidina
(poliamina) via 0 intermediario N1-glutationil-espermidina (GSH-SPD). Estudos em Crithidia
'H •••~~H 0
o H
N H SH ~HJ,
,
HH'
I t
S H ( ~ H a ) 4
~ NH
• H
H ~ 0
H
coo' 0
~
-H 0
+H,N N-...~ ..•o
o ) H ~
S ( C H a ) ,
\ I
\
NH+
I 1
S ( C H a ) .
~ ~H + H H 0 H COO· 0
Figura 111·1.Formula estrutural dos substratos glutationa (A) e tripanotiona (B) J antes e depois da
r
T[S],T io l H :20 2 R a d ic a is
d is s u lfe to p e r 6 x id o s Iiv r e s
I
.
o r g lin ic o s1--
L
T [S H hI
10; m e ta is p e s a d o s
F ig u r a 1 1 1 - 3D ia g r a m a e s q u e m a tic o p a r a in d ic a r a r e g r a d e p r o te ~ a o b io l6 g ic a n o m e ta b o lis m o d o s
53 58
1 MMSKI PDLVVIGAGSGGLEAAWNAATL YKKRVAVIDVQMVHGPPPPSALGG
1
_ _
.l·PKKLMVTGAQYMEHLRESAGPGWEFD lACRQEPQPQGPPPAAGAVASYDYLVIGGGSGGLASARRAAEL GARAAVVESHK LGG~PKKVMWNTAVHSEPMHDHADYG FPI
IOIRTTLRAEWKNLIAVKDEAVLNINKSYDEMPRDTEGLEPPLGWGSLESKNVVNVRESADPASAVKERLETEHILLASGSWPHMPNIPGIEHCISSNEAFYL101SCEGKFNWRVlKE~YVSRLNAIYQNNLTKSH IEIIRGHAAFTSDPKPTIEVSGKKYTAPHILIATGG MPST PHESQIPGASLGITSDGFFNL1201PEPPRRVLTVGGGFISVEFAGIFNAYKPKDGQVTLCYRGEMILRGPDHTLREELTKQLTANGIQIL201EELPGRSVIVGAGY IAVEMAGILSALGSKTSLKI RHDKVLRSPDSMI STNCTEELENAGVEVLKPSQVKEVKKTLSGLEVSMVTAVPGRLPVMTMIPTKENPAKVELNADGSKSVTFESGKKMDFDLV
1301301DVDCLLWAIGRVPNTKDLSLNKLGIQTDDKGHIIVDEFQNTNVKGIYAVGDVCGKALLTPVAlAAGRKLAHRLPEYKEDSKLDYNNIPTVVPSHPPIGTVMM AIGRSPRTKDLQLQNAGVMIKNGG VQVDEYSRTNVSNIYAIGDVTNRVMLTPVAINEAAALVDTVFG TTPRKTDHTRVASAVPSIPPIGTC
1401GLIEEVASKRYEV401GLTEDEAIHKYGIENVKTYSTSFTPMYHAVTKRKTKCVAVYLSSFTPLMHKVSGSKYKTPVAKIITNHSDGTVLGVHLLGDNAPEIIQGIGICLKLNAKISDFYNTIG\fVMLMVCANKEEKVVGIHMQGLGCDEMLQGFAVA VKMGAYKADFDNTVAl{il>!fYSSEELVTLR'rSAEELCSMRT
I
501PSYYYVKGEKMEKP501F ig u r a 1 1 1-A lin h a m e n to4 s e q u e n c ia I e n tr e T R d e Trypanosoma cruz; e G R h u m a n a . O s r e s id u o s m a is im p o r ta n te s p a r a a c a ta lis e e s U io s a lie n ta d o s e m v e r m e lh o .
, .C
,r-.
i ~ ~i)
C A P iT U L o ill-T R IP IlN O T IO N A R E D U T A S E D E T o C R lJ Z J 32
Segundo, os compostos podem ser enzimaticamente reduzidos e enta~ reoxidados pelo
oxigenio molecular, sabre 0 cicio redox. Muitos destes compostos tem atividade tripanocidal
contra T . c r u z i in v itr o comprovando sua capacidade de atuar como um substrato do cicio
redox.
Do ponto de vista de desenho de drogas, tripanotiona a um excelente alvo para a
modelagem de inibidores especificos. TR e seu substrato tripanotiona sao essenciais para a
sabrevivencia da celula do protozoario, unicamente encontrada nestes parasitas e a
especifica com relac;ao ao seu substrato quando com parada com a enzima analoga humana,
a glutationa redutase. Desta forma, a razoavel buscar um composto quimioterapeutico que
possa ser reconhecido pela TR do parasita, mas nao pelo hospedeiro.
Neste sentido, tada e qualquer informac;ao a respeito da enzima pede ser importante
quando da modelagem de inibidores. Estudos bioquimicos e cristalograficos tem side
realizados e detalhes estruturais em nivel atomico da tripanotiona redutase (Zhang, Y. et ai,
1993; Zhang, Y. et ai, 1995) ja se encontram na Iiteratura.
o
objetivo deste projeto a mais um passe na caracterizac;ao da TR. Atravas de estudosde mutagenese sitio dirigido (Borges, et ai, 1995) e da determinac;ao da estrutura
cristalografica da enzima nativa e de diferentes complexos (Hunter, W.N. et ai, 1992; Bailey,S.
et ai, 1993; Bailey,S. et ai, 1994), TR tem side caracterizada, residuos associados
a
suafunc;ao tem side identificados, e a variac;ao conformacional diante da ligac;ao com diferentes
substratos (T[S]2,[GSPd]2) obseNadas, mas nao havia nenhuma evidencia do comportamento
da enzima durante a ligac;ao do substrato, isto a, quando 0 substrato se encontra Iigado a
enzima.
Neste sentido 0 projeto visou 0 estudo cristalografico da mutante C58S TR + GSPd,
onde a enzima Tripanotiona redutase teve sua cisteina catalitica 58 ( destacadas na Figura
-glutationil-espermidina,
0substrato precursor da tripanotiona no cicio metab6lico das poliaminas (Figura
111-3).A estrutura foi determinada utilizando difra9Bo de raios-X em monocristais a
2 .4 Ade
CAPITuLO 111- TRlPANOTlONA R£DUTASE DE To CRUZ! 34 ( F ig u r a 1 1 1 - 5 )c r e s c e r a m a d im e n s c 5 e s d e a p r o x im a d a m e n t e O . 5 x O.5 x 1 . 0 m m e m t o m e d e u m a
s e m a n a .
111.3Coleta e processamento dos dados
C r is t a is d o c o m p le x o m u t a n t e C 5 8 S T r ip a n o t io n a R e d u t a s e + G S P d p e r t e n c e m a o
g r u p o e s p a c ia l P 4 3 , p a r a m e t r o s d e c e la d e a = b = 9 2.7, c = 1 5 6.2 A c o m u m d f m e r o n a u n id a d e
a s s im e t r ic a .
O s c r is t a is d if r a t a m a t e 2 . 3 A d e r e s o lu c ; : a o,e m b o r a c o m a a n a lis e d a s e s t a t f s t ic a s d e
in t e g r a c ; : a o e p r o m e d ia c ; : a o d o s d a d o s, s o m e n t e d a d o s a t e 2.4A t e n h a m s id o u t iliz a d o s n o
C A P I T u L O 1 1 1 - T R I P A N O T I O N A R E D U T A S E D E ToCRUZl 3 5
O s p rim e iro s d a d o s fo ra m c o le ta d o s a 2 .6 A d e re s o lu y a o , u s a n d o ra d ia y a o s in c ro tro n
c o m c o m p rim e n to d e o n d a ( A ) d e 0 .9 2 A . A s 1 2 5 6 1 0 re fle x 6 e s m e d id a s fo ra m p ro m e d ia d a s e m 4 9 1 6 6 re fle x o e s in d e p e n d e n te s p a ra u m R s y md e 0 .0 5 8 (c o n ju n to d e d a d o s I). E s te s
d a d o s fo ra m s u p le m e n ta d o s p o r n o v a s m e d id a s re a liz a d a s a 2 .3 A d e re s o lu y a o u s a n d o u m
c o m p rim e n to d e o n d a d e 1 .1 5 A (c o n ju n to d e d a d o s II) n u m to ta l d e 5 5 4 2 5 re fle x o e s m e d id a s
p ro m e d ia d a s e m 2 9 8 8 3 re fle x O e s in d e p e n d e n te s , R s y md e 0 .0 8 8 .
A m b o s c o n ju n to s d e d a d o s fo ra m c o le ta d o s n a e s ta y a o P X 9 .5 d a F o n te d e R a d ia y a o
S in c ro tro n , L a b o ra t6 rio d e D a re s b u ry , u s a n d o a p la c a d e im a g e m (3 0 0 m m ), M A R c o m o
d e te to r. Q u a tro c ris ta is fo ra m u tiliz a d o s , e v a ria s tra n s la 9 5 e s a o lo n g o d o s e u e ix o m a io r fo ra m
n e c e s s a ria s d e s d e q u e o s c ris ta is s e m o s tra ra m s e n s iv e is
a
in te n s a ra d ia y a o d o fe ix e .A lg u m a s im a g e n s n a o fo ra m u tiliz a d a s d e v id o a m a q u a lid a d e c a u s a d a p e lo d a n o p a r
ra d ia c a o . O s d a d o s fo ra m p ro c e s s a d o s u s a n d o 0 p ro g ra m a M O S F L M (L e s lie e t a i, 1 9 8 6 ) e 0
p a c o te d e p ro g ra m a s C C P 4 (C C P 4 , 1 9 9 4 ), p a ra c a d a p o s iy a o tra n s la c io n a l d o s c ris ta is
s e p a ra d a m e n te . U m to ta l d e 1 7 0 1 5 5 re fle x O e s fo ra m m e d id a s , d o s q u a is 5 0 4 1 0
in d e p e n d e n te s , 9 8 .1 % d o s d a d o s a 2 .4 A d e re s o lu y a o , R s y m
=
0 .0 8 3 . E s ta tis tic a s re le v a n te sa
c o m p le ta c o le ta d e d a d o s s a o a p re s e n ta d a s n a T a b e la 1 1 1 -1 .
111.4
D e te rm in a ~ io
da estrutura e refinamento
O s c ris ta is d o c o m p le x o m u ta n te C 5 8 S T R + G S P d s e m o s tra ra m is o m 6 rfic o s a o s d a
C A P I T u L O I I I · T R I P A N O T l O N A R E D U T A S E D E ToCRUZI
T a b e la 1 1 1 -1E s ta tis tic a s re le v a n te s a c o le ta d e d a d o s . R s y m
=
LI
I(k )-(I) 1 1 L 1 (k )o n d e I(k ) e (I) re p re s e n ta m a In te n s id a d e d e u m a d a d a re fle d o e a m e d ia d a s in te n s id a d e s re s p e c tiv a m e n te e( Jo d e s v io p a d rA o .
R e s o l u y a o
(A )
R e f l e x o e s I n d e p e n d e n t e s
1 > 3 0 ' ( % )
7.14 1824 0.053 96.4
5.22 3018 0.057 98.2
4.31 3857 0.054 99.1
3.75 4528 0.062 99.4
3.37 5097 0.074 99.1
3.08 5588 0.097 98.7
2.86 6076 0.137 99.0
2.68 6493 0.173 98.7
2.53 6906 0.244 98.7
2.40 7023 0.307 95.1
C A P I T u L O 1 1 1 - T R I P A N O T I O N A R E D U T A S E D E ToCRUZ/ 37
u m p ro to c o lo d e re s fria m e n to le n to n o q u a l a te m p e ra tu ra s im u la d a d o s is te m a fo i re d u z id a d e
2 0 0 0 K p a ra 3 0 0 K e m p a s s o s d e 5 0 K n o te m p o d e 0 .5 fs (R fa c to r=2 2 .6 % ).
F o ra m e n ta o c o n s tru id o s o s p rim e iro s m a p a s d e d e n s id a d e e le tr6 n ic a (2 IFc b s
I -
IFc a lcIe IFo b s l- IF c a lc !)e e x a m in a d o s n a s e s ta c ;O e s g ra fic a s a tra v e s d o p ro g ra m a g ra fic o 0 (J o n e s
et
aI,
1 9 9 1 ), p ro g ra m a e s te u tiliz a d o e m to d a s a s m a n ip u la c ;O e s g ra fic a s d o m o d e lo .O s p rim e iro s m a p a s ja m o s tra v a m c la ra d e n s id a d e p a ra p a rte d o lig a n te (G s p d ) e m
a m b a s s u b u n id a d e s . A m u ta ~ o (C 5 8 S ) e 0 lig a n te (G S P d ) fo ra m in c lu id o s n o re fin a m e n to e
s e in ic io u a m o d e la g e m p a ra a s re g i6 e s o n d e a m a p a d e d e n s id a d e n a o c o in c id ia c o m a
m o d e lo .
N o q u a rto c ic io , a v in c u lo d e s im e tria n a o c ris ta lo g ra fic a (e ix o d e o rd e m 2 ) fo i lib e ra d o
e a s d u a s s u b u n id a d e s re fin a d a s in d e p e n d e n te m e n te . A s a Q u a s fo ra m id e n tific a d a s c o m o
p ic o s d e a ltu ra m a io r q u e 3 c r ( o n d e c r e o d e s v io p a d ra o d o m a p a ) e m m a p a s d ife re n c ;a
(I
F o b s l-lF c a lc !) u tiliz a n d o a p ro g ra m a P E A K M A P (C C P 4 , 1 9 9 4 ).
A p 6 s 4 c ic lo s d e re fin a m e n to p o s ic io n a l e d o s fa to re s d e te m p e ra tu ra (A P E N D IC £ 6 )
c o n ju n to d e d a d o s IIfo i in c lu id o n o re fin a m e n to . U m a u m e n to n o fa to r d e d is c o rd a n c ia (R fa cto r)
c ris ta lo g ra fic o p a ra 2 1 .6 fo i v e rific a d o .
C ic lo s d e re fin a m e n to p o s te rio re s in c o rp o ra n d o v in c u lo s in d iv id u a is , fa to re s d e
te m p e ra tu ra e a d i~ o c o n s e rv a tiv a d e m o le c u la s d e s o lv e n te fo ra m u tiliz a d o s p a ra c o m p le ta r
a re fin a m e n to . 0 m o d e lo fin a l p o s s u i u m fa to r d e d is c o rd a n c ia d e 1 9 .8 % c o m a d e s v io d a
g e o m e tria id e a l d e 0 .0 1
A
p a ra c o m p rim e n to s d e lig a ~ o , 2 .0 0 0 p a ra a n g u lo s d e lig a ~ o e2 3 .40
p a ra a n g u lo s d ie d ric o s . A T a b e la 1 1 1 -2in c lu i a s e s ta tis tic a s d e re fin a m e n to d o c o m p le x o
C ic io R e s o lu ~ o R -fa c to r r.m .s m o le c u la s
A in ic . - fin . c o m p o (A ) a n g . ( 0 ) d e a Q u a
1 2 . 6 2 4 . 1 - 2 1 . 5 0 . 0 0 6 1 . 4 4 5 a 2 - 3 2 . 6 2 5 . 1 - 1 9 . 8 0 . 2 1 1 2 . 8 5 5 9 1 4 - 5 2 . 4 2 0 . 0 - 2 3 . 3 0 . 0 0 9 4 . 4 1 8 1 1 5 6 - 2 6 2 . 4 2 3 . 3 - 1 9 . 8 0 . 0 1 0 2 . 0 0 0 2 6 5
111.5Resultados
cristalograficos
o
m o d e lo fin a l c o m p re e n d e u m d fm e ro , d u a s m o le c u la s d e F A D , d u a s m o le c u la s d eN1-g lu ta tio n il-e s p e rm id in a (G s p d ) e 2 6 5 m o le c u la s d e s o lv e n te tra ta d a s c o m o a to m o s d e
to ta l d e 7 8 7 4 a to m o s n a o h id ro g e n io s c o m u m p a ra m e tro te rm ic o is o tr6 p ic o m e d io , Ba v =
T a b e la 1 1 1-3 .V a lo re s p a rc ia is p a ra o s p a ra m e tre s te rm ic o s is o tr6 p ic o s p a ra C 5 3 5 T R + G s p d . N A e e q u iv a le n te a o n u rn e ro d e a to m o s e Ba v a o fa to r d e te m p e ra tu ra m e d io e m (A2). M C e q u iv a le a c a d e ia pli':l~ ipC lI ..~.S .~.a.C :C lciE Jia.IC ltE JrC lI •.Y C lI~ ~ s..c:a.Ic:~ IC lci()S.H~ ~ ':lci~. E 5 " " , t ; ~ 9 J : ,.~ ~ r : » ~.( ~ ~ r : » ~ , 1 9.~ ) .
S u b u n id a d eA S u b u n id a d e B A g u a s T o ta l
M C S C G s p d F A D M C S C G s p d F A D
N A 1 9 3 1 1 7 8 7 2 9 5 3 1 9 3 5 1 7 9 2 2 9 5 3 2 6 5 7 8 7 4
Ba v
2 6.1 3 1 . 1 5 3 . 7 1 6 . 3 3 3.7 3 8.4 7 2.6 2 5.3 3 8 . 1 3 4.9. .. . . . • • • •••• •• •• •• • • • ••• • • • • • . ?• . • . ••• ... ,. .............,...,..,.•.... , ...,...
~
· u
c
'§.
'"
. 0 ;
- 0
~
[ 3 0 :·lr·ir.i··H····~·J~n··t'.·f"··!'fl·.rft;\·ft-tH ~ti;.HHH,~·¥\l1r1tll.:;;1~i'-,:iIH··Jyl:·,,H .~...H_~.._.HiH H.. 0 '- ~: ~ ".".~ _~ ~ ~~1JJ VI: ;'1 ~i l~.: , " l.:,fl.t ~~A li ~tilJ::,' ":;J"~~~I '-,
20 .,'H ·••··l~ r'··H" ..H _H H !H ~H .~ . ..,~_H _H H HIo..¥H-·.a··.•.~··,··V V .····,.-+ ~·t;H : r,.~v; .
!
10 .:.:.H H····I··H~H H..H + .H__H _.H l.~ :H H..~~ !:'.LHH.H H.I;.._ ~H H H_.l.H..:H H~ .]..YH\ .-
·
f
:
Hr .Figura 111-8Mapa de omissao (Fobs-Fcalc, acalc, mapa calculado excluindo as coordenadas da serina 58) ao nivel de 30".
, . ~ ; :. u 'c. rB l ! ~T ! [C A
IF s rI, i - .D ~L:o '
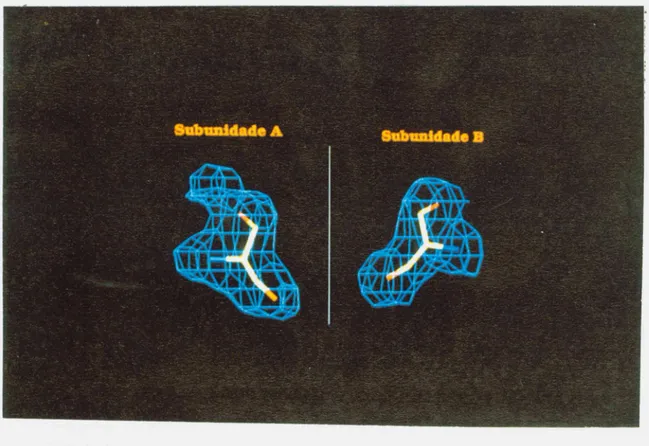
F ig u ra 1 1 1 -9V is ta e s te re o g rM ic a p a ra G S P d e s e u a ju s te c o m 0 m a p a d e o m is s a o F o b s -F c a lc ( c a lc u lo d o
m a p a o m itin d o a s c o o rd e n d a s d o lig a n te n o m o d e le a o n iv e l3 a . a ) s u b u n id a d e A b ) s u b u n id a d e B .
F ig u ra 1 1 1 -1 0S u p e rp o s i~ a o e n tre a s m o le c u la s d e G S P d p a ra a s d u a s s u b u n id a d e s . E m a z u l s u b u n id a d e