Respostas celulares e moleculares
em interacções hospedeiras e não hospedeiras
cafeeiro-ferrugem (Hemileia vastatrix e Uromyces vignae)
Inês Isabel Plácido dos Santos Diniz
Dissertação para a obtenção do Grau de Mestre em
Engenharia Agronómica
Orientador: Doutora Maria do Céu Machado Lavado da Silva
Co-orientador: Doutora Maria Helena Mendes da Costa Ferreira Correia de
Oliveira
Júri:
Presidente: Doutora Cristina Maria Moniz Simões de Oliveira, Professora Associada do Instituto Superior de Agronomia da Universidade Técnica de Lisboa
Vogais:
- Doutora Maria Helena Mendes da Costa Ferreira Correia de Oliveira, Professora Associada do Instituto Superior de Agronomia da Universidade Técnica de Lisboa
- Doutor Arlindo Lima, Professor Auxiliar do Instituto Superior de Agronomia da Universidade Técnica de Lisboa
- Doutora Maria do Céu Machado Lavado da Silva, Investigadora Auxiliar do Centro de Investigação das Ferrugens do Cafeeiro do Instituto de Investigação Científica Tropical
- Doutora Luísa Cristina dos Mártires Ferreira de Carvalho, Investigadora Auxiliar do Instituto Superior de Agronomia da Universidade Técnica de Lisboa
Agradecimentos
Na execução deste trabalho estiveram envolvidadas pessoas e instituições às quais gostaria de agradecer.
À minha orientadora, Doutora Maria do Céu Silva, com o profundo reconhecimento e gratidão, pela forma esclarecedora com que sempre me orientou, pelos valiosos ensinamentos que me transmitiu e pelo apoio e encorajamento sempre prestados.
À Professora Doutora Maria Helena Oliveira, por ter aceite ser minha orientadora durante esta tese perante o Instituto Superior de Agronomia e pela sua pronta disponibilidade e interesse demonstrado.
Ao Doutor Pedro Talhinhas um agradecimento especial pela transmissão de conhecimentos, pelas longas discussões e valiosas sugestões e pela pronta disponibilidade para troca de impressões e colaboração dispensados.
À Doutora Helena Azinheira, um agradecimento especial pela pronta disponibilidade para troca de impressões e valiosas sugestões.
À Doutora Leonor Guerra-Guimarães, Eng.º Vitor Várzea e Doutora Dora Batista pela disponilidade, incentivo e amizade demonstrada.
À Drª Paula Pereira, Doutora Andreia Loureiro e a todos os funcionários do CIFC, em particular à Sandra Emídio, Célia Lopes, Idalina Gomes e Maria João Bettencourt, assim como à Fernanda Rodrigues, pelo incondicional companheirismo, amizade e pela capacidade de rir quando tudo corre mal.
Um agradecimento muito especial à Paula Leandro por todo o apoio técnico dado durante a execução do trabalho experimental e também pela sua amizade, companheirismo, e incansável apoio que sempre me dispensou, quando tudo parecia sem sentido.
A todos os meus amigos em especial à Rita, Tânia, Teresa, Sandro, Ritinha, Isabel e Mafalda por estarem por perto quando eu mais precisei.
Ao João, por partilhar comigo momentos de tristeza e de alegria, pela paciência e apoio demonstrados e por acreditar em mim e me incentivar a continuar.
Aos meus meus pais e irmã pelo apoio incondicional, pelo carinho e compreensão, pela paciência e amizadade durante todo este tempo.
À minha família!
Resumo
Caracterizou-se a nível celular e molecular a resistência dos cafeeiros HDT 832/2 e Sarchimor a Hemileia vastatrix, usando o modelo comparativo de resistência não hospedeira, cafeeiro-Uromyces vignae. Em ambos os cafeeiros, H. vastatrix cessou o crescimento com maior frequência na fase de hifa de penetração formando haustórios numa percentagem reduzida de zonas de infecção, induzindo reacção de hipersensibilidade (RH), acumulação de fenóis e encapsulamento dos haustórios com calose. U. vignae apresentou menor crescimento em HDT 832/2 não formando haustórios em ambas as plantas, induzindo RH e acumulação de fenóis.
Estudou-se por PCR em Tempo Real, a expressão dos genes rlk, wrky1, pal, chs, 13-lox, gt, pr1b e pr10 envolvidos em mecanismos de reconhecimento, sinalização e de defesa. Nas duas interacções, o perfil dos genes rlk e wrky1 sugere um reconhecimento do fungo e sinalização durante a diferenciação dos apressórios e das primeiras fases da infecção; no gene 13-lox o primeiro pico de activação coincidiu com o início da RH. O gene pr1b apresentou elevados níveis de expressão na interacção hospedeira sendo pouco activado na interacção não hospedeira. A activação dos genes 13-lox e pr1b sugere que as vias do SA e JA coexistem, embora a via do SA pareça prevalecer na interacção hospedeira.
Palavras-chave: cafeeiro, ferrugem alaranjada, resistência hospedeira, resistência não-hospedeira,
Cellular and molecular responses in host and non-host coffee-rust
interactions (Hemileia vastatrix and Uromyces vignae)
Abstract
Cellular and molecular characterization of resistance to H. vastatrix in coffee lines HDT 832/2 and Sarchimor, was conducted using the non-host resistance model coffee-Uromyces vignae (cow pea rust) for comparison. In both lines, H. vastatrix ceased growth more frequently at the penetration hypha stage, forming few haustoria, inducing hypersensitive reaction (HR), phenol accumulation and haustorium encasement with calose. U. vignae growth was more restricted in HDT 832/2, but failed to form haustoria in both lines, while inducing HR and phenol accumulation.
The expression of genes involved in mechanisms of recognition, signaling and defence (rlk, wrky1, pal, chs, 13-lox, gt, pr1b and pr10) was monitored by real time PCR. Both in the host and non-host interactions the expression profile of rlk and wrky1 suggests that the recognition of the fungus and signaling occur during the formation of appressoria over stomata and the differentiation of the first post-penetration infection stages. The first activation peak of 13-lox coincided with the onset of cell death detection. pr1b exhibited high expression levels in the host interaction, being poorly activated in the non-host interaction. These results suggest that both JA and SA pathways may coexist, although SA pathway seams to prevail in the host interaction.
Key words: coffee, coffee leaf rust, host resistance, non-host resistance, hypersensitive reaction,
Cellular and molecular responses in host and non-host coffee-rust
interactions (Hemileia vastatrix and Uromyces vignae)
Extended abstract
Leaf rust, caused by the fungus Hemileia vastatrix, is considered the main disease of Arabica coffee around the world. Breeding resistant coffee varieties (such as Sarchimor) has been the most efficacious strategy against this disease, namely using descendents of the Híbrido de Timor (HDT) as a source of resistance against all known races of H. vastatrix.
The objective of this work was the characterization, at cellular and molecular levels, of the resistance to H. vastatrix in coffee lines HDT 832/2 and Sarchimor, using the non-host resistance model coffee-Uromyces vignae (cow pea rust) for comparison. This study fits into a broader objective aiming at inferring the durability of coffee resistance to rust.
Bright field and fluorescence microscopy techniques were used for the analysis of fungal pre- and post-penetration and of responses induced in plant cells. H. vastatrix (isolate 1065, race II) and U. vignae (isolate CPR-1, race I) pre-penetration stages (germinated uredospores and appressoria differentiated over stomata) were similar in HDT 832/2 and in Sarchimor. In both coffee lines, H. vastatrix cessed growth more frequently at the penetration hypha stage, reaching haustoria formation only in a small percentage of infection zones (bellow 10%). Similar responses were induced by the fungus in the host plants, namely a rapid death of the cells in the stomatic zone (hypersensitive reaction - HR), an accumulation of phenols and an encasement of haustoria with calose. On the other hand, U. vignae cessed growth in more advanced infection stages in Sarchimor as compared to HDT 832/2, but failed to form haustoria in both coffee lines. This pre-haustorial resistance was associated to the rapid death of stomatic cells (HR) and the accumulation of phenols. Although these studies showed analogies on resistance to H. vastatrix between the two coffee lines, U. vignae presented a more restrict growth in HDT 832/2, which was therefore chosen for molecular studies.
The expression of eight genes involved in mechanisms of recognition, signaling and defence (rlk, wrky1, pal, chs, 13-lox, gt, pr1b and pr10) was monitored by real time PCR. Constitutive genes gapdh and ubiquitin were used as normative genes after an evaluation of the individual and conjugated gene expression stability. Most genes were activated in the host interaction (HDT 832/2–H. vastatrix). Genes rlk, wrky1, pal, 13-lox, pr1b, pr10 and gt presented two activation peaks, the first between 6 and 12 hours after inoculation (h.a.i.) (a period when appressoria differentiation and penetration hypha formation occur, and the plant responses at cellular level begin) and the second between 21-24 h.a.i. (when anchors and haustoria mother cells differentiate and plant responses are recorded in ca. 50% of infection zones).
In the non-host interaction (HDT 832/2–U. vignae) most genes are not regulated or poorly activated in the first hours after inoculation (3-6 h.a.i.), being activated at 9-12 h.a.i. (a period from which no new fungal infection structures are differentiated, and plant responses are detected in over 50% of infection zones).
and the differentiation of the first post-penetration infection stages. Although the accumulation of phenols was observed in the cells of the infection zone (from 12 h.a.i. in the host interaction and from 6 h.a.i. in the non-host interaction), the pal gene was only moderately activated in the host interaction (with inconclusive results in the non-host interaction), while the chs gene was non-regulated. However, this accumulation of phenols may arise from post-translational modifications in pal or from the expression of other genes. The 13-lox gene was moderately activated in both interactions, with the first activation peak coinciding in both cases with the onset of the detection of cell death (HR). 13-lox and pr1b genes are used as markers respectively of the jasmonate and the salicylic acid pathways. pr1b exhibited high expression levels in the host interaction, being poorly activated in the non-host interaction. These results suggest that both pathways may coexist, although the salicylic acid pathway seams to prevail in the host interaction.
Índice Geral
AGRADECIMENTOS ... I RESUMO ...II ABSTRACT ...III EXTENDED ABSTRACT... IV ABREVIATURAS ... XII I – INTRODUÇÃO GERAL ...1 1. O CAFEEIRO ...1 1.1. Taxonomia ...1 1.2. Características gerais ...21.3. Espécies de importância económica e sua distribuição geográfica ...3
1.3.1. Coffea arabica L. ...3
1.3.2. Coffea canephora Pierre ex Froehner ...3
1.4. Importância económica da produção do café ...4
1.5. Produção e consumo mundial de café ...5
2. A FERRUGEM ALARANJADA - HEMILEIA VASTATRIX ...7
2.1. Importância económica ...7
2.2. Origem e distribuição geográfica ...7
2.3. Taxonomia ...7
2.4. Sintomatologia ...8
2.5. Ciclo de vida ...8
2.6. Processo de infecção ...9
2.8. Meios de luta...9
3. A INTERACÇÃO CAFEEIRO-HEMILEIA VASTATRIX ... 11
3.1. Raças fisiológicas de Hemileia vastatrix ... 11
3.2. Grupos fisiológicos de Coffea spp. ... 12
3.3. Hereditariedade da resistência a Hemileia vastatrix em Coffea spp. ... 13
4. CONSIDERAÇÕES GERAIS SOBRE A RESISTÊNCIA DAS PLANTAS AOS AGENTES PATOGÉNICOS ... 14
4.1 Reconhecimento do agente patogénico, transmissão de sinal e respostas de defesa ... 15
4.2. Respostas de defesa induzidas ... 16
4.3. Mecanismos celulares, bioquímicos e moleculares de resistência do cafeeiro a Hemileia vastatrix ... 17
5. PCR EM TEMPO REAL NO ESTUDO DA EXPRESSÃO GÉNICA ... 21
6. OBJECTIVOS ... 23
II - MATERIAL E MÉTODOS ... 24
1. PLANTAS E FUNGOS ... 24
2. MANUTENÇÃO DAS PLANTAS... 24
3. PRODUÇÃO E MANUTENÇÃO DE INÓCULO... 25
4. INOCULAÇÃO ... 25
5. OBSERVAÇÕES AO MICROSCÓPIO ÓPTICO ... 26
5.1. Germinação de uredósporos in vivo e formação de apressórios ... 27
5.2. Processo de colonização do fungo e respostas celulares das plantas ... 27
6. TÉCNICAS MOLECULARES ... 29
6.2 Síntese de cDNA... 31
6.3. PCR em tempo real ... 31
7. GENES EM ESTUDO ... 32
8. ANÁLISE DOS RESULTADOS ... 32
III - RESULTADOS ... 34
1. ESTUDOS DE MICROSCOPIA ... 34
1.1. Análise macro e microscópica das interacções do cafeeiro com Hemileia vastatrix e com Uromyces vignae ... 34
1.1.1. Tipos de reacção ... 34
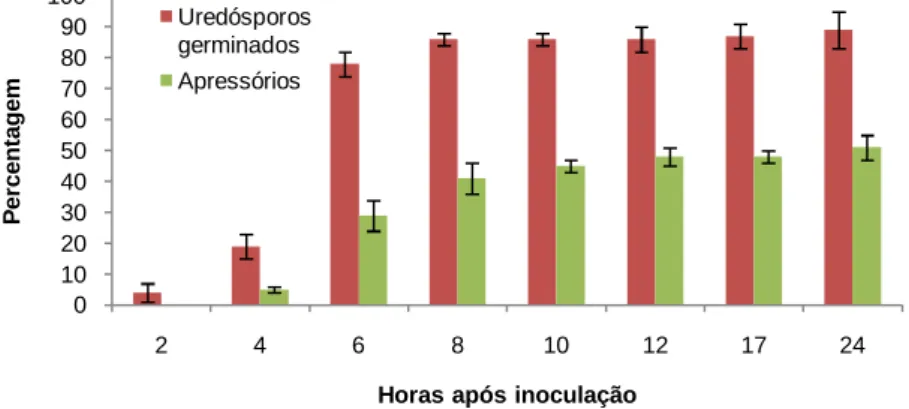
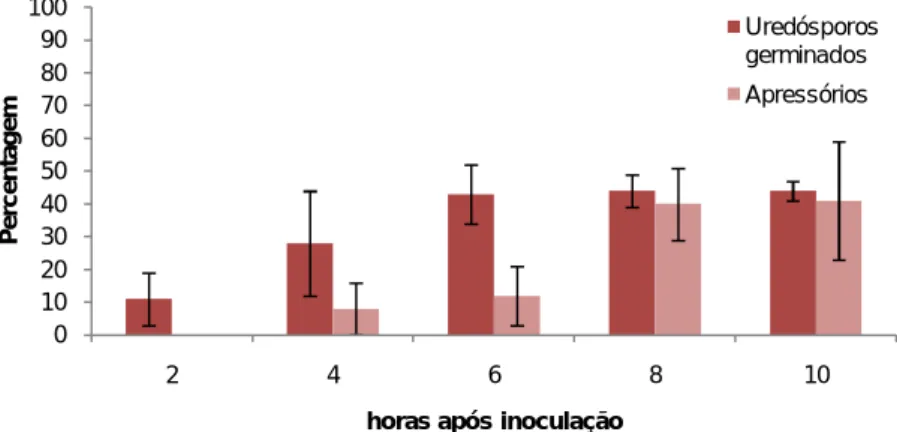
1.1.2. Germinação in vivo e formação de apressórios de Hemileia vastatrix e de Uromyces vignae ... 34
1.1.3. Processo de colonização de Hemileia vastatrix ... 36
1.1.4. Processo de colonização de Uromyces vignae ... 38
1.1.5. Respostas histológicas de HDT e Sarchimor à infecção por Hemileia vastatrix ... 42
1.1.6. Respostas histológicas de HDT e Sarchimor à infecção por Uromyces vignae ... 43
2. DETERMINAÇÃO DOS TEMPOS PARA ESTUDO DA EXPRESSÃO GÉNICA RELATIVA ... 46
2.1. Germinação in vivo de Hemileia vastatrix e de Uromyces vignae ... 46
2.2. Processo de colonização de Hemileia vastatrix e de Uromyces vignae ... 47
2.3. Respostas histológicas induzidas por Hemileia vastatrix e Uromyces vignae ... 48
3. DETERMINAÇÃO DOS GENES CONSTITUTIVOS ... 51
4. EXPRESSÃO GÉNICA RELATIVA ... 54
4.1. Expressão relativa do gene rlk... 54
4.2. Expressão relativa do gene wrky1 ... 54
4.3. Expressão relativa do gene pal... 55
4.6. Expressão relativa do gene gt ... 56
4.7. Expressão relativa do gene pr1b ... 57
4.8. Expressão relativa do gene pr10 ... 57
4.9. Comparação da expressão relativa dos genes selecionados ... 58
IV – DISCUSSÃO ... 59
V- CONCLUSÕES E PERSPECTIVAS ... 64
REFERÊNCIAS BIBLIOGRÁFICAS ... 65
Índice de quadros
Quadro 1. 1 - Produção de café a nível mundial e nos principais países produtores/continente, para a
colheita de 2008/2009 . ...5
Quadro 1. 2 - Consumo mundial de café nos países produtores e importadores, para o ano-safra de
2008/2009 ...6
Quadro 1. 3 - Escala dos tipos de reacção dos cafeeiros a Hemileia vastatrix elaborada por D'Oliveira
(1954-1957). ... 11
Quadro 2. 1 - Cafeeiros, nº de identificação no CIFC, origem, grupo fisiológico e genes de resistência
das plantas de cafeeiro usadas. ... 24
Quadro 2. 2 - Iniciadores utilizados para amplificação por qPCR dos genes em estudo. ... 32 Quadro 3. 1 - Percentagem de uredósporos germinados in vivo e de apressórios formados por
Uromyces vignae, na planta hospedeira (Vigna unguiculata) e no cafeeiro (HDT 832/2), 24 h.a.i... 35
Quadro 3. 2 - Percentagem de uredósporos germinados in vivo e de apressórios formados por
Hemileia vastatrix em HDT 832/2 e Sarchimor, 24 horas após a inoculação. ... 35
Quadro 3. 3 - Percentagem de uredósporos germinados in vivo e de apressórios formados por
Uromyces vignae em HDT 832/2 e Sarchimor, 24 horas após a inoculação... 35
Quadro 3. 4 - Diferenciação das primeiras estruturas de infecção de Hemileia vastatrix, em folhas de
HDT 832/2, em diferentes tempos após inoculação. ... 36
Quadro 3. 5 - Formação de haustórios e comprimento médio das hifas intercelulares de Hemileia
vastatrix em folhas de HDT 832/2, em diferentes tempos após a inoculação. ... 37
Quadro 3. 6 - Diferenciação das primeiras estruturas de infecção de Hemileia vastatrix em folhas de
Sarchimor, em diferentes tempos após inoculação. ... 37
Quadro 3. 7 - Formação de haustórios e comprimento das hifas intercelulares de Hemileia vastatrix
em folhas de Sarchimor, em diferentes tempos após inoculação. ... 38
Quadro 3. 8 – Diferenciação das primeiras estruturas de infecção Uromyces vignae, em diferentes
tempos após inoculação em folhas de HDT 832/2. ... 39
Quadro 3. 9 - Comprimento das hifas intercelulares de Uromyces vignae em folhas de HDT 832/2. . 39 Quadro 3. 10 - Diferenciação das primeiras estruturas de infecção de Uromyces vignae, em
diferentes tempos após inoculação em folhas de Sarchimor. ... 40
Quadro 3. 11 - Comprimento das hifas intercelulares de Uromyces vignae em folhas de Sarchimor. 40 Quadro 3. 12 - Respostas histológicas em tecidos foliares de HDT 832/2 inoculados com Hemileia
vastatrix. ... 42
Quadro 3. 13 - Respostas histológicas em tecidos foliares de Sarchimor inoculados com Hemileia
vastatrix. ... 43
Quadro 3. 14 - Respostas histológicas em tecidos foliares de HDT 832/2 inoculados com Uromyces
vignae. ... 44
Quadro 3. 15 - Respostas histológicas em tecidos foliares de Sarchimor inoculados com Uromyces
Quadro 3. 17 - Respostas histológicas à infecção por Uromyces vignae em folhas de HDT 832/2. ... 49 Quadro 3. 18 – Estabilidade dos genes constitutivos gapdh e ubiquitina e da sua conjugação,
Índice de figuras
Fig. 1. 1 - Uredósporo de Hemielia vastatrix com zona equinulada e outra lisa ...7 Fig. 1. 2 - Página inferior da folha do cafeeiro com pústulas de soros uredospóricos de cor alaranjada.
...8
Fig. 2. 1 - Esquema das primeiras fases do processo de infecção de Hemileia vastatrix... 26 Fig. 2. 2 - Esquema das primeiras fases do processo de infecção de Uromyces vignae. ... 27
Fig. 3. 1 – Tipos de reacção observados. (A) Reacção flt (B) Reacção i 34
Fig. 3. 2 – Diferenciação de estruturas de infecção de Hemileia vastatrix em tecidos foliares de HDT
832/2 ... 36
Fig. 3. 3 – Diferenciação de estruturas de infecção de Uromyces vignae em tecidos foliares de HDT
832/2.. ... 39
Fig. 3. 4 – Percentagem de zonas de infecção com diferenciação das primeiras estruturas de
infecção de Uromyces vignae, em tecidos foliares de HDT 832/2 e de Sarchimor, em diferentes tempos após a inoculação.. ... 41
Fig. 3. 5 – Curvas de crescimento de Uromyces vignae nos tecidos foliares de HDT e Sarchimor, em
diferentes tempos após a inoculação. ... 41
Fig. 3. 6 – Respostas induzidas por Hemileia vastatrix em tecidos foliares de HDT 832/2... 42 Fig. 3. 7 – Respostas induzidas por Uromyces vignae em tecidos foliares de HDT 832/2 ... 44 Fig. 3. 8 – Percentagem de uredósporos germinados e de apressórios diferenciados por Hemileia
vastatrix em folhas de HDT 832/2, em diferentes tempos após inoculação. ... 46
Fig. 3. 9 – Percentagem de uredósporos germinados e de apressórios diferenciados por Uromyces
vignae em folhas de HDT832/2, em diferentes tempos após inoculação. ... 47
Fig. 3. 10 – Percentagem de zonas de infecção com diferenciação das primeiras fases do processo
de infecção de Hemileia vastatrix em folhas de HDT 832/2, em diferentes tempos após inoculação. . 47
Fig. 3. 11- Percentagem de zonas de infecção com diferenciação das primeiras fases do processo de
infecção de Uromyces vignae em folhas de HDT 832/2, em diferentes tempos após inoculação... 48
Fig. 3. 12 – Variabilidade de expressão do gene constitutivo gapdh, em tecidos foliares de HDT 832/2
quando inoculado com Hemileia vastatrix (A) e com Uromyces vignae (B), ensaio 1. ... 51
Fig. 3. 13 - Variabilidade de expressão do gene constitutivo gapdh, em tecidos foliares de HDT 832/2
quando inoculado com Hemileia vastatrix (A) e com Uromyces vignae (B), ensaio 2. ... 51
Fig. 3. 14 - Variabilidade de expressão do gene constitutivo ubiquitina, em tecidos foliares de HDT
832/2 quando inoculado com Hemileia vastatrix (A) e com Uromyces vignae (B), ensaio 1... 52
Fig. 3. 15 - Variabilidade de expressão do gene constitutivo ubiquitina, em tecidos foliares de HDT
832/2 quando inoculado com Hemileia vastatrix (A) e com Uromyces vignae (B), ensaio 2... 52
Fig. 3. 16 - Variabilidade da expressão média dos genes constitutivos gapdh/ubiquitina, em tecidos
foliares de HDT 832/2 quando inoculado com Hemileia vastatrix (A) e com Uromyces vignae (B), no ensaio 1. ... 52
Fig. 3. 17 - Variabilidade de expressão média dos genes constitutivos gapdh/ubiquitina, em tecidos
foliares de HDT 832/2 quando inoculado com Hemileia vastatrix (A) e com Uromyces vignae (B), no ensaio 2. ... 53
Fig. 3. 18 - Expressão relativa do gene rlk, em dois ensaios, ao longo de vários tempos após
inoculação numa interacção hospedeira (A) e numa interacção não hospedeira (B). ... 54
Fig. 3. 19 - Expressão relativa do gene wrky1, em dois ensaios, ao longo de vários tempos após
inoculação numa interacção hospedeira (A) e numa interacção não hospedeira (B). ... 54
Fig. 3. 20 - Expressão relativa do gene pal , em dois ensaios, ao longo de vários tempos após
inoculação numa interacção hospedeira (A) e numa interacção não hospedeira (B). ... 55
Fig. 3. 21 - Expressão relativa do gene chs, em dois ensaios, ao longo de vários tempos após
inoculação numa interacção hospedeira (A) e numa interacção não hospedeira (B). ... 55
Fig. 3. 22 - Expressão relativa do gene 13-lox, em dois ensaios, ao longo de vários tempos após
inoculação numa interacção hospedeira (A) e numa interacção não hospedeira (B). ... 56
Fig. 3. 23 - Expressão relativa do gene gt, em dois ensaios, ao longo de vários tempos após
inoculação numa interacção hospedeira (A) e numa interacção não hospedeira (B). ... 56
Fig. 3. 24 - Expressão relativa do gene pr1b, em dois ensaios, ao longo de vários tempos após
inoculação numa interacção hospedeira (A) e numa interacção não hospedeira (B). ... 57
Fig. 3. 25 - Expressão relativa do gene pr10, em dois ensaios, ao longo de vários tempos após
Abreviaturas
ABA – Ácido abcísico
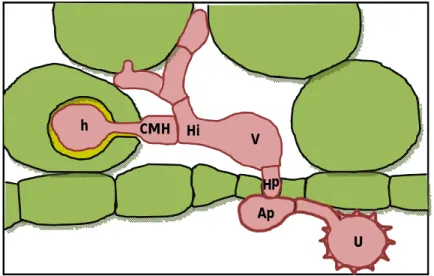
Ap – Apressório
cDNA – DNA complementar
CHS – Chalcona sintase
CIFC – Centro de Investigação das Ferrugens do Cafeeiro
CMH – Célula mãe do haustório
Ct – Threshold cicle
d.a.i. – Dias após inoculação
DNA – Ácido desoxiribonucleico
dNTP – Mistura de desoxiribonucleóticos fostafatos dATP, dCTP, dGTP, dTTP
EDTA – Ácido etilenodiamina tetra-acético
ET – Etileno
GAPDH – Gliceraldeído-3-fosfato desidrogenase
GT – Glicosiltransferase
h – Haustório
h.a.i. – Horas após inoculação
HDT – Híbrido de Timor
Hi – Hifa de infecção
Hp – Hifa de penetração
HR – Reacção de hipersensibilidade
ICO – International Coffee Organization (Organização Internacional do Café)
JA – Jasmonato
LOX – Lipoxigenase
mRNA – RNA mensageiro
NADPH – Fosfato de dinucleótido de nicotinamida e adenina
NO – Óxido nítrico
pb – Pares de bases
PCR – reacção em cadeia da polimerase
POD – Peroxidase
PR – Proteínas relacionadas com a patogenicidade
qPCR – PCR quantitativo
RAPD – Amplificação aleatória de polimorfismo de DNA
RLK – Receptor-like kinase
RNA – Ácido ribonucleico
RSA – Resistência sistémica adquirida
ROS – Espécies reactivas de oxigénio
rpm – Rotações por minuto
RT – Transcriptase reversa
SA – Ácido Salicílico
SDS – Dodecil-sulfato de sódio
SOD – superóxido dismutase
TAE – Tris-acetato EDTA
Tris – Tris(hidroximetil)aminometano
U – Unidade de actividade enzimática
Ur – Uredósporo
UV – Ultravioleta
NOTA PRÉVIA
O trabalho reportado nesta dissertação foi efectuado no Centro de Investigação das Ferrugens do Cafeeiro/Instituto de Investigação Cientifica Tropical, em colaboração com o Departamento de Protecção das Plantas e Fitoecologia do Instituto Superior de Agronomia/UTL e insere-se no âmbito do projecto PTDC/AGR-AAM/71866/2006 - "Abordagem interdisciplinar para a caracterização de potenciais fontes de resistência durável do cafeeiro (Coffea spp.) à ferrugem alaranjada (H. vastatrix)” financiado pela Fundação para a Ciência e a Tecnologia.
I – Introdução Geral
1. O cafeeiro
1.1. Taxonomia
O cafeeiro (Coffea spp.) pertence à família Rubiaceae e à sub-família Ixoroideae (Andreasen & Bermer, 2000; Davis et al., 2006). A primeira descrição botânica do cafeeiro surgiu em 1713 por A. de Jussieu. Usando apenas uma planta originária do Jardim Botânico de Amsterdão, Jussieu classificou o cafeeiro como Jasminum arabicanum (Correia, 1995), homenageando assim a semelhança do aroma das suas flores com o jasmim e o local de origem da planta, a Arábia (Ferrão, 2009). Em 1734, Linnaeus integrou-a num género diferente, Coffea, classificando-a como Coffea arabica, a única espécie de cafeeiro conhecida até então.
Na década de 40 do século XX, Auguste Chevalier elaborou uma classificação baseada, exclusivamente, na distribuição geográfica e nas características morfológicas dos cafeeiros originários das regiões tropicais e subtropicais da África e Ásia conhecidos até então (Correia, 1995). Chevalier dividiu então o género Coffea L. em 4 secções; Paracoffea (13 spp.), Argocoffea (11 spp.), Mascarocoffea (18 spp.) e Eucoffea (24 spp.). Por sua vez, a secção Eucoffea, divisão onde se encontram reunidas todas as espécies de interesse económico, foi dividida em 5 subsecções, de acordo com vários critérios morfológicos e geográficos: Erythrocoffea (4 spp.) e Melanocoffea (3 spp.) (coloração do fruto), Nanocoffea (5 spp.) (altura da planta), Pachycoffea (5 spp.) (consistência da folha), e Mozambicoffea (7 spp.) (distribuição geográfica). As espécies Coffea arabica, C. congensis, C. canephora e C. eugenioides foram reunidas na subsecção Erythrocoffea, enquanto C. liberica foi englobada na subsecção Pachycoffea (Carvalho et al., 1969).
O desenvolvimento de novas técnicas aplicadas ao estudo da sistemática permitiu criticar e sugerir alterações à classificação de Chevalier. Leroy (1980), que propôs uma nova divisão do género Coffea, chega mesmo a afirmar que apenas uma taxonomia integrativa, com base em estudos de fisiologia e biologia, poderá relacionar os diversos caracteres de forma a colocar em evidência os processos de evolução, conduzindo assim à obtenção de resultados mais objectivos e clarificar as relações filogenéticas entre os táxones.
A classificação mais recente considera o género Coffea (103 spp.) dividido em dois subgéneros; Coffea (95 spp.) e Baracoffea (8 spp.) (Orozco-Castillo et al., 1996; Cros et al., 1998). O subgénero Coffea, composto por espécies todas elas originárias de África, Madagáscar e Ilhas das Mascarenhas, engloba todas as espécies de importância económica, enquanto o subgénero Baracoffea engloba apenas oito espécies que se localizam, exclusivamente, nas florestas secas do oeste de Madagáscar (Davis et al., 2006).
A aplicação de técnicas de biologia molecular no estudo filogenético do género Coffea (Anthony et al., 2002; Aga et al., 2003; Moncada & McCouch, 2004; Poncet et al., 2006) tem revelado resultados muito semelhantes aos obtidos até agora pela taxonomia clássica, como seja, a confirmação da
origem africana do género e a sua diferenciação em inúmeras espécies (Orozco-Castillo et al., 1996; Cros et al., 1998), ao mesmo tempo que demonstra a enorme variabilidade genética dentro deste. Para além disso, estas técnicas têm sido usadas para conhecer a origem de C. arabica uma vez que alguns estudos apontam para que C. eugenioides e C. congensis sejam os progenitores diplóides do cafeeiro Arábica (Raina et al., 1998), enquanto outros apontam para que sejam C. eugenioides e C. canephora (Orozco-Castillo et al., 1996; Cros et al., 1998; Lashermes et al., 1999; Moncada & McCouch, 2004).
A grande proximidade genética entre o género Coffea e o género Psilanthus sugere a necessidade de uma revisão na sistemática do género Coffea de forma a abranger espécies de outros géneros próximos (Lashermers et al. 1997; Cros et al. 1998).
É ainda de referir que recentes prospecções aos locais de origem do género Coffea, como a Tanzânia, resultaram na identificação de mais duas espécies (Davis et al., 2006).
Coffea é um género complexo e de grande diversidade genética, cuja taxonomia está sujeita a frequentes revisões resultantes de inovações introduzidas nos métodos modernos de biologia molecular que, em conjugação com a aplicação dos métodos táxonómicos clássicos, deverão visar um conhecimento mais aprofundado da variabilidade genética de Coffea, de forma a suportar os programas de melhoramento (Ruas et al., 2003).
1.2. Características gerais
O cafeeiro é uma dicotiledónea, perene e de porte arbustivo que no estado selvagem pode atingir entre 8 a 10 m de altura. Possui um caule vertical e lenhoso do qual se desenvolvem pares de ramos laterais, opostos e horizontais (plagiotrópicos), dos quais se produzem, nas axilas das folhas do ano anterior, flores e consequentemente frutos.
As folhas são pecioladas, opostas, aos pares, ovais ou lanceoladas, com um tamanho que pode variar entre 1 a 40 cm, e possuem uma cor que varia entre o amarelo e o verde-escuro.
Flores normalmente brancas, de corola 4-9-lobada, agrupadas em inflorescências com duas a seis flores. O período de floração ocorre depois da época das chuvas e a maturação dos frutos ocorre nove meses depois da floração.
Os frutos são drupas, normalmente carnudas, com duas sementes revestidas por um endocarpo e mesocarpo. Antes de amadurecer, os frutos são de cor verde, e consoante a variedade podem ser amarelos ou vermelhos quando maduros.
As sementes são ovais ou elípticas e plano-convexas (Purseglove, 1974; Wrigley, 1988; Cardoso, 1994).
Entre as espécies do género Coffea, apenas C. arabica L. é autogâmica e alotetraplóide com 2n = 44 cromossomas (Davis et al., 2006). As restantes espécies, como sejam, C. canephora, C. dewerei, C. congensis, C. eugenioides, C. racemosa e C. stenoplylla entre outras, são alogâmicas e diplóides com 2n = 22 cromossomas (Carvalho et al., 1969; Rodrigues Jr. & Bettencourt, 1975).
1.3. Espécies de importância económica e sua distribuição geográfica
Das inúmeras espécies de cafeeiros existentes, apenas duas têm importância económica. São elas C. arabica L., vulgarmente conhecido como cafeeiro Arábica e C. canephora Pierre ex Froehner, vulgarmente conhecido por cafeeiro Robusta (Carvalho et al., 1969; Orozco-Castillo et al., 1996; Davis et al., 2006; Maurin et al., 2007). C. liberica e C. racemosa contribuem com quantidades ínfimas de café produzidos mundialmente (Osório, 2009).
Das espécies mais importantes, C. arabica L. representa cerca de 70 a 75% da produção mundial de café, enquanto C. canephora representa os restantes 25 a 30% (Osório, 2009).
1.3.1. Coffea arabica L.
O cafeeiro Arábica é originário das regiões montanhosas da Etiópia e do planalto de Boma do Sudão. É uma espécie adaptada a regiões de elevadas altitudes, entre os 1000 a 2000 m, com precipitações bem distribuídas, entre os 1500 a 2000 mm anuais, intercaladas por um período seco bem definido, e com uma temperatura variando entre 15ºC e 24ºC, sendo o ideal de crescimento os 20ºC (www.ico.org).
O seu processo de domesticação iniciou-se na Arábia e aí se manteve exclusivamente cultivada durante cinco séculos. Pela mão dos Holandeses expandiu-se pela Ásia, Ceilão (actual Sri-Lanka) (1690) e Java (1696 a 1699), de onde foi introduzida no continente americano (1718). Os Portugueses levaram a cultura para o Brasil (1727). O maior período de expansão desta espécie foi somente em fins do século XIX e princípios do século XX, onde chegou ao Malawi (1878), ao Zaire (1885), ao Quénia (1896), ao Uganda e à Tanzânia (1910), e aos Camarões (1929) (Cardoso, 1994).
Actualmente, produz-se café Arábica essencialmente na América Latina e em alguns países da África Central e Oriental e Ásia (Rodrigues Jr., 2002). A sua importância económica deve-se ao café de boa qualidade que produz e com baixo teor de cafeína (0,8 a 1,4%) (Rodrigues Jr. & Bettencourt, 1975; Correia, 1995).
Esta espécie, no entanto, é susceptível a uma série de pragas e doenças, sendo importante referir a sua susceptibilidade a Hemileia vastatrix, agente causal da ferrugem alaranjada do cafeeiro (Correia, 1995).
Por ser uma espécie autogâmica, as suas variedades tendem a ser geneticamente estáveis mas as mutações espontâneas com características desejáveis têm sido exploradas para a obtenção de cultivares. Exemplos disso são as variedades Bourbon Vermelho, Bourbon Amarelo, Caturra Vermelho, Caturra Amarelo, Novo Mundo, Catuaí Vermelho, Catuaí Amarelo, Blue Montain, Pacas, San Ramon, Villa Sarchi e Villa Lobos (Carvalho, 1969).
1.3.2. Coffea canephora Pierre ex Froehner
O cafeeiro Robusta é originário das florestas equatoriais africanas. É uma espécie adaptada a baixas altitudes que vão desde o nível do mar até os 1000 m, a elevadas precipitações entre os 2000 a 3000 mm anuais, e a uma temperatura variável entre os 24ºC e os 30ºC, sendo os 27ºC a temperatura ideal de crescimento (www.ico.org).
Só nos finais do século XIX é que se iniciou a sua domesticação na Ásia. A resistência do cafeeiro Robusta à ferrugem alaranjada permitiu repor os cafezais da Índia, Sri-Lanka e Java que tinham sido dizimados pela ferrugem alaranjada (Cardoso, 1994). Tornou-se assim a espécie mais cultivada na Ásia e em África, onde ocupa regiões climaticamente menos favoráveis ao cultivo de Arábica.
Actualmente é cultivado, essencialmente, na África Ocidental e Central, em todo o sudeste da Ásia e no Brasil (Rodrigues Jr., 2002).
C. canephora produz um café de menor qualidade, com teores elevados de cafeína (1,7 a 4,0%) e com um sabor amargo. O Robusta é largamente usado para lotar com o Arábica de forma a melhorar a bebida do café ao conferir-lhe mais corpo (Correia, 1995; Amidou et al., 2005; Ferrão, 2009), para além de ser também usado para a produção de café solúvel (Rodrigues Jr., 2002; Ferrão, 2009).
Esta espécie do género Coffea é uma fonte de resistência a uma série de pragas e doenças, nomeadamente à ferrugem alaranjada, tal como já foi referido.
Por ser uma espécie alogâmica, é muito polimórfica, e por vezes, as suas variedades são difíceis de caracterizar. As principais variedades de C. canephora são as Robusta (Typica, Gosseweilleri, Welwitshsii), Kouillou (Conillon), Guarani, Laurentii, Oka, Uganda, Crassifolia e Bukobensis (Carvalho, 1969).
1.4. Importância económica da produção do café
O café é o segundo produto mais importante no comércio internacional, logo depois do petróleo. Estima-se que toda a actividade envolvida na produção do café comercial (cultivo, processamento e comercialização) representa uma fonte de rendimento para cerca de 125 milhões de pessoas a nível mundial (Rodrigues Jr., 2002; Osório, 2005).
Para muitos países em desenvolvimento, a produção de café é a principal fonte de divisas e uma das principais ocupações de mão-de-obra, chegando a ser a principal fonte de rendimento de países como o Burundi, Uganda, Etiópia e Tanzânia (Rodrigues Jr., 2002).
Para a maioria dos países cafeicultores, a indústria do café representa a chave das suas economias. No entanto, o desequilíbrio entre a oferta e a procura tem provocado grandes oscilações de preços levando a indústria do café a atravessar várias crises. Exemplo disso foi a situação vivida em 2001/2002 quando a produção total de café foi de 113 milhões de sacas (60 Kg cada saca), enquanto o consumo mundial foi de apenas 106 milhões de sacas (www.ico.org). Estas situações de crise resultam na falência de pequenos e médios produtores e, consequentemente, em graves situações sociais como a fome e a emigração.
Com o intuito de controlar o mercado de produção de café, foi criada, em 1963, a International Coffee Organization (ICO) sediada em Londres e da qual fazem parte os principais países produtores e importadores de café. Desde a sua formação, a ICO passou a gerir todos os “Acordos Internacionais do Café” que se vinham a realizar desde 1962, tendo como objectivo evitar as oscilações nos preços do café através do estabelecimento de cotas de exportação (Correia, 1990). Mesmo com uma gestão feita pela ICO, os preços do café continuaram a oscilar muito devido ao conflito de interesses entre os países produtores e importadores (Correia, 1995), mas também devido
aos 27% dos custos totais de produção, e da mão-de-obra em muitos países exportadores (Osório, 2009).
1.5. Produção e consumo mundial de café
Segundo o relatório de Dezembro de 2009 da ICO, o ano-safra de 2008/2009 terminou com uma produção total de café de cerca de 128 milhões de sacas, o que equivale a um aumento de 7,4% em relação à produção total no ano-safra anterior de 2007/2008.
Em 2008/2009 a produção de café Arábica totalizou 61% da produção total e a produção de café Robusta totalizou os restantes 39% da produção total.
A observação do Quadro 1.1 permite verificar que a América do Sul foi o maior produtor (47,1%), seguido da Ásia e Oceânia (27,3%), América do Norte e Central (13,8%), e por fim África (11,9%). Dos países produtores de café, o Brasil foi o principal produtor contribuindo com 35,9% da produção mundial, seguido do Vietname (14,4%), Indonésia (7,3%) e Colômbia (6,8%).
Quadro 1. 1 - Produção de café a nível mundial e nos principais países produtores/continente, para a colheita de 2008/2009 (saca = 60Kg) (Osório, 2009).
Em relação ao consumo mundial, segundo o relatório de Dezembro da ICO (2009), houve um aumento de cerca de 1,5% em 2008 em relação ao ano civil de 2007. A título informativo, no Quadro 1.2 estão representados os valores de consumo mundial de café em 2008.
Continente País Produção (milhões de sacas) Continente País Produção (milhões de sacas) Mundo 128,181
África 15,259 América do Norte e Central 17,689
Etiópia 4,35 México 4,651
Uganda 3,2 Honduras 3,450
Costa do Marfim 2,353 Guatemal a 3,785
Tanzânia 1,186 Costa Rica 1,318
Quénia 0,572 El Salvador 1,547
Camarões 0,750 Ásia e Oceânia 34,926
América do Sul 60,308 Vietname 18,5
Brasil 45,992 Indonésia 9,35
Colômbia 8,664 Índia 4,372
Equador 0,691 Papua Nova Guiné 1,028
Quadro 1. 2- Consumo mundial de café nos países produtores e importadores, para o ano-safra de 2008 (Osório, 2009).
Verifica-se assim que os Estados Unidos da América são o principal país consumidor (16,7%), seguido do Brasil (13,4%) e Alemanha (7,3%).
País Consumo
(milhões de sacas) País
Consumo (milhões de sacas)
Mundo 130,004
Países produtores 36,703 Países importadores 93,302
Brasil 17,526 Países da U. Europeia 39,850
Indonésia 3,333 Alemanha 9,535
México 2,2 Itália 5,937
Etiópia 1,833 França 5,156
Índia 1,518 Espanha 3,485
Filipinas 1,07 Reino Unido 3,067
Colômbia 1,4 Holanda 1,324 Vietname 1,021 Suécia 1,272 Venezuela 1,599 Polónia 1,19 Finlândia 1,115 Grécia 0,978 EUA 21,652 Japão 7,065 Rússia 3,716 Canadá 3,214 Argélia 2,118 Ucrânia 1,733 Rep. Da Correia 1,665 Austrália 1,145
2. A ferrugem alaranjada - Hemileia vastatrix
2.1. Importância económica
A ferrugem alaranjada é considerada a principal doença do cafeeiro Arábica a nível mundial e estima-se que seja responsável por quebras de 15 a 20% da produção mundial de café, se não forem tomadas quaisquer medidas de controlo (van der Vossen, 2001).
2.2. Origem e distribuição geográfica
A ferrugem alaranjada foi registada pela primeira vez em 1861, perto do Lago Vitória, Quénia, na África Oriental. No entanto, foi no Sri-Lanka em 1868, que causou maior impacto ao dizimar em menos de 10 anos a cultura do café nesse país, levando à falência de empresas cafeicultoras, ao abandono dos campos cultivados ou à sua substituição pela cultura do chá. Rapidamente a doença se propagou para outras regiões como a Índia, Java e Filipinas, causando sempre enormes prejuízos.
Em 1878 a doença atingiu o continente Africano proliferando, ainda que lentamente, da província do Natal, África do Sul (1878) até à África Oriental. Angola, na África Ocidental, registou os primeiros ataques da ferrugem alaranjada em 1966. Pouco mais tarde, a doença atingiu o Brasil (1970), a Colômbia (1983), a Papua Nova Guiné e a Jamaica (1986). Actualmente, a ferrugem alaranjada está presente em todos os países cafeicultores, excepto no Hawai e Austrália (Bettencourt & Rodrigues Jr., 1988; van der Vossen, 2001; Rodrigues Jr., 2002; Várzea et al., 2004).
2.3. Taxonomia
O género Hemileia (hemi, meio e leia, lisa) engloba cerca de 40 espécies, provenientes de zonas tropicais, que infectam monocotiledóneas e dicotiledóneas mas predominantemente Rubiáceas (Rodrigues Jr., 1990). São apenas duas as espécies com importância económica, Hemileia coffeicola e H. vastatrix, ambas infectando plantas do género Coffea de elevado valor económico (Rodrigues Jr. & Bettencourt, 1975; McGrain & Hennen, 1984). Das duas espécies referidas, a mais importante é H. vastatrix (Fig. 1.1), uma vez que se encontra disseminada por todos os países produtores de café, para além de causar maiores prejuízos.
H. vastatrix foi pela primeira vez descrita por Berkeley e Broome, a partir de amostras de folhas de C. arabica L. colhidas no Sri-Lanka.
Fig. 1. 1 -Uredósporo de Hemielia vastatrix
com zona equinulada e outra lisa
Actualmente, o fungo H. vastatrix está classificado da seguinte forma (Kirk et al., 2008):
REINO - Fungi
DIVISÃO - Basidiomycota CLASSE - Pucciniomycetes ORDEM - Pucciniales FAMÍLIA - Incertae sedis GÉNERO - Hemileia
ESPÉCIE – Hemileia vastatrix Berk & Br.
2.4. Sintomatologia
H. vastatrix forma, na página inferior das folhas de cafeeiro, lesões cloróticas que, posteriormente, se cobrem de uredósporos formando pústulas de soros uredospóricos de cor alaranjada e de dimensão variável (Fig. 1.2
).
Mais tarde geralmente ocorre a necrose e a senescência prematura da folha. Nos casos de ataques mais intensos, pode ocorrer desfoliação severa ou completa e eventualmente a morte da planta (Wellman, 1952; D’Oliveira 1954-57).Fig. 1. 2 – Página inferior da folha do
cafeeiro com pústulas de soros
uredospóricos de cor alaranjada.
2.5. Ciclo de vida
H. vastatrix é um fungo biotrófico, com um ciclo de vida onde se conhecem três tipos de esporos: uredósporos, teleutósporos e basidiósporos. Os uredósporos e os teleutósporos são produzidos no mesmo tipo de soros, mas em condições diferentes. Os uredósporos são dicarióticos e correspondem à fase assexuada do ciclo, re-infectando sucessivamente as folhas de cafeeiros, sempre que as condições lhe sejam favoráveis, iniciando assim a fase de doença (Rodrigues Jr., 1980).
Os teleutósporos são considerados esporos de resistência e hibernação que se desenvolvem consoante a época do ano. Germinando in situ, sem necessidade de período de dormência e após meiose, os teleutósporos originam os basidiósporos (monocarióticos), esporos característicos da fase sexuada que, apesar de também apresentarem germinação in situ, não infectam as folhas do cafeeiro. Por essa razão, pensa-se que H. vastatrix seja uma ferrugem heteróica. No entanto, uma vez que H. vastatrix não está relacionada com qualquer grupo de ferrugens heteróicas e as tentativas de se encontrar o hospedeiro alternante têm falhado, será também de considerar que esta ferrugem poderá ser do tipo autóica (Rodrigues Jr., 1980; Rodrigues Jr. et al., 2000).
Em condições de estufa, é ainda possível observar teleutósporos uredinóides, que são morfologicamente idênticos aos uredósporos mas que germinam como teleutósporos. A sua função, no entanto, não é conhecida (Rodrigues Jr., 1980).
2.6. Processo de infecção
O processo de infecção das folhas de cafeeiro por H. vastatrix inicia-se com a germinação dos uredósporos na página inferior das folhas do cafeeiro, onde se localizam os estomas, sendo obrigatória a presença de água líquida e temperaturas entre 15ºC - 30ºC (Rijo & Rodrigues Jr., 1977). Depois de penetrar pelo estoma, o fungo coloniza os tecidos foliares com formação de hifas intercelulares e de haustórios. Cerca de três a quatro semanas após a infecção, junto à área de penetração, forma-se uma grande densidade micelial de onde surge para o exterior, novamente através do estoma, um soro uredospórico em forma de bouquet. Destes soros são libertados uredósporos que, em condições favoráveis, germinam dando origem a um novo processo de infecção (Rijo & Rodrigues Jr., 1977).
2.7. Modos de disseminação
Os uredósporos são transportados facilmente e em grandes quantidades pelo vento e pela chuva, assim como pelo Homem e pelos animais. Para além disso, o transporte de material contaminado entre países, do mesmo continente ou de continentes diferentes, também contribuiu para a rápida disseminação de H. vastatrix. Os uredósporos resistem a temperaturas muito baixas mas são particularmente sensíveis à secura (Agrios, 2005).
2.8. Meios de luta
A luta química tem sido o meio mais utilizado para combater a ferrugem alaranjada. De entre os diversos fungicidas os cúpricos, como o oxicloreto de cobre, são os mais eficazes como meio preventivo (Rodrigues Jr. & Bettencourt, 1975; Hillocks et al., 1999; Rodrigues Jr., 2002; Várzea et al., 2002). Estes fungicidas possuem um efeito tónico nas plantas que se traduz na cor verde-escura das folhas e no retardamento da desfolha (Cunha et al., 2004).
No entanto, para além da aplicação dos fungicidas cúpricos ser de elevado custo (podendo chegar aos 10% dos custos de produção) e de estar dependente das condições meteorológicas, a sua aplicação regular tem uma acção poluente e tóxica na camada superficial do solo, para além de causar um desequilíbrio na flora e fauna aí existente e provocar alterações nas propriedades do solo como o aumento do pH e desequilíbrio nutricional (Akinnifesi et al., 2006
)
. Para além disso, quando o cobre não é adsorvido, é lixiviado pela água pluvial e canalizado para os rios. O excesso de cobre também tem efeitos nocivos para as plantas tais como inibição do alongamento das raízes, inibição da fotossíntese, inibição ou mesmo supressão da actividade enzimática envolvida na síntese da clorofila e perturbações no metabolismo do azoto (Rodrigues Jr., 2002).Os fungicidas sistémicos têm sido também bastante utilizados. Quando comparados com os fungicidas cúpricos, os sistémicos são mais eficientes na sua absorção, translocação e modo de acção no controlo da ferrugem, para além de necessitarem de um menor número de aplicações e de estas serem menos dependentes das condições meteorológicas (Cunha et al., 2004).
Os elevados custos dos fungicidas, assim como a sua difícil aplicação devido ao declive de algumas plantações, elevado custo de maquinaria para sua aplicação, necessidade de água para a sua aplicação, falta de formação e falta de informação adequada, como acontece frequentemente nos países africanos (Hillocks et al., 1999), tem direccionado os centros de investigação e experimentação como o Centro de Investigação das Ferrugens do Cafeeiro (CIFC, Portugal), o Central Coffee Research Institute (Índia), vários institutos no Brasil, o Central Research Station (Quénia) entre outros, para a selecção de cafeeiros resistentes à ferrugem alaranjada, uma vez que só assim se obtém um controlo eficiente da doença.
Um marco importante na obtenção de variedades resistentes a H. vastatrix deu-se em finais dos anos 50 quando, em Timor-Leste, foram descobertos alguns genótipos de Híbridos de Timor (HDT) (híbrido natural entre C. arabica e C. canephora) resistente a todas as raças conhecidas de H. vastatrix.
Muitas são as variedades de cafeeiro resistentes à ferrugem actualmente cultivadas em diferentes países cafeicultores provenientes de cafeeiros e populações, nomeadamente derivados de HDT, criadas e estudadas no CIFC. Destacam-se as populações vulgarmente designadas por Catimor (cv Caturra x HDT CIFC 832/1) e Sarchimor (cv Villa Sarchi x HDT CIFC 832/2) que deram origem a várias cultivares. A partir de Catimor foram criadas as seguintes cultivares: Oeiras MG 6851 e Katipó (Brasil), Cauvery (Índia), Costa Rica 95 (Costa Rica), IHCAFÉ 90 e Lempira (Honduras), Oroazteca (México), Catisic (El Salvador) e MIDA 96 (Panamá). Da população Sarchimor originaram-se as cultivares Lapar 59, Obatã, Tupi e IPR 98 (Brasil) e Chandragiri (Índia). Referem-se ainda a variedade Colombia derivada da população Caturra x CIFC HDT 1343, assim como as cultivares brasileiras Araponga MG1, Catiguá MG1, MG2, MG3, Paraíso MG H419-1, Pau Brasil MG1 e Sacramento MG1, descendentes da população (Catuaí x HDT CIFC 2570) e IBC-Palma 1 e IBC Palma 2 da população Catuaí x Catimor (Bettencourt et al., 1998; Silva et al., 2006; Carvalho, 2008; Várzea et al., 2008).
O aparecimento de novas raças fisiológicas de H. vastatrix com maiores espectros de virulência tem, no entanto, levado à quebra de resistência de algumas dessas variedades (Várzea & Marques, 2005).
3. A interacção cafeeiro-Hemileia vastatrix
Os diversos tipos de reacção das interações cafeeiro-ferrugem são determinados usando uma escala de classificação qualitativa elaborada por D’Oliveira (1954-1957), apresentada no Quadro 1.3.
Quadro 1. 3 - Escala dos tipos de reacção dos cafeeiros a Hemileia vastatrix elaborada por D'Oliveira (1954-1957).
A expressão mais comum de resistência é o fleck geralmente associado à tumefacção (flt). As reacções do tipo 1 e 2 são designadas, respectivamente, como moderadamente resistente (MR) e moderadamente susceptível (MS), e as reacções 3 e 4 como susceptível (S).
3.1. Raças fisiológicas de Hemileia vastatrix
De um modo geral, a quase totalidade dos agentes patogénicos, em particular os fungos, apresenta raças fisiológicas ou patótipos, ou seja, variantes com diferentes genes de virulência e consequente capacidade para infectar de forma diferenciada distintos genótipos de uma dada espécie (Rodrigues Jr., 2002).
Tipos de Rea cção
Expressão fenotípica da interacção cafeeiro- ferrugem
i Imune, sem quaisquer sinais macroscópicos que indiquem que se deu a infecção
fI Fleck reacção de hipersensibilidade, às vezes difícil de observar macroscopicamente, mas visível à lupa
; Pontuações necróticas, visíveis macroscopicamente, situadas no ponto de penetração da ferrugem ou dispersas pela área de infecção
T Pequena tumefacção noponto de penetração da ferrugem, bem visível à lupa
0 Clorose, mais ou menos intensa, na área de infecção, às vezes acompanhada de pequenas necroses, mas sem formação de soros uredosporíferos
1 Raros soros uredosporíferos, sempre muito pequenos, por vezes só distinguíveis à lupa, em áreas predominantemente cloróticas, por vezes acompanhadas de necroses
2 Pústulas uredospóricas pequenas ou médias, difusas mas bem visíveis macroscopicamente, em áreas intensamente cloróticas
3 Pústulas uredospóricas média s ou grandes, rodeadas de clorose
4 Grandes pústulas uredospóricas sem verdadeira hipersensibilidade, mas podendo apresentar uma leve clorose na margem das infecções (altamente congenial ou susceptível)
X Reacção heterogénea, pústulas uredospóricas de tamanho muito variável, incluindo na aparência diversos tipos ou graus de infecção com expressões de congenialidade e incongenialidade
As primeiras quatro raças fisiológicas de H. vastatrix foram identificadas na Índia, por Mayne (1932, 1935, 1936, 1942). Este trabalho foi depois continuado no CIFC, e actualmente estão identificadas cerca de 45 raças fisiológicas, aí mantidas como colecção viva (Várzea & Marques, 2005).
Estas raças foram isoladas a partir de amostras de ferrugem de várias regiões cafeicultoras. A caracterização da(s) raça(s) de H. vastatrix é realizada pela observação das reacções fenotípicas por esta(s) induzida(s) em 18 cafeeiros diferenciadores provenientes de diferentes regiões cafeicultoras (Várzea et al., 2002).
Os diferenciadores são um conjunto de plantas que consistem em clones de: cinco plantas da espécie tetraplóide C. arabica; seis híbridos interspecíficos tetraplóides C. arabica x C. canephora e C. arabica x C. liberica; seis plantas das espécies diplóides C. canephora, C. congensis, C. excelsa e C. racemosa e um C. arabica x HDT (Rodrigues Jr. & Bettencourt, 1975; Bettencourt & Rodrigues Jr., 1988; Várzea & Marques, 2005).
A raça II é a predominante estando presente em quase todos as regiões produtoras de café (Rodrigues Jr. & Bettencourt, 1975; Bettencourt & Rodrigues Jr., 1988; Várzea & Marques, 2005).
O recurso a marcadores moleculares, como RAPD, sequenciação de regiões ITS do rDNA e a análise de perfis ISSR, permitiu encontrar níveis de diversidade genética entre isolados de H. vastatrix que, embora baixos, são mais elevados no conjunto de isolados provenientes de África e da Ásia, ou seja, nos locais onde a doença existe há mais tempo (Gouveia et al., 2005; Gonzalez, 2009).
Pensa-se que a principal causa da variabilidade genética de H. vastatrix seja a mutação genética, uma vez que é desconhecida a sua fase sexuada e o seu hospedeiro alternante. Assim sendo, o aparecimento de novas raças fisiológicas de H. vastatrix pode estar relacionado com a pressão exercida pela selecção de genes de resistência do hospedeiro (Rodrigues Jr. et al., 2000; Várzea et al., 2002).
3.2. Grupos fisiológicos de Coffea spp.
Os grupos fisiológicos de cafeeiro distinguem-se entre si pelo espectro de reacção que apresentam em relação às diferentes raças fisiológicas de H. vastatrix. O espectro de reacção pode variar entre a resistência total (grupo A), parcial ou intermédia (grupos D,α, C, γ, J, L, I, G, H, e W) ou a susceptibilidade total a todas as raças fisiológicas de H. vastatrix (grupos E, F, β e N) (Rodrigues Jr., 2002).
No grupo A estão incluídas algumas espécies de Coffea, híbridos espontâneos (Híbrido de Timor) e híbridos artificiais (híbridos resultantes do cruzamento de C. arabica e HDT), como o Catimor, Cavimor, Sarchimor entre outros obtidos no CIFC. No grupo E estão incluídos a maioria dos cafeeiros Arábica cultivados, como Typica, Bourbon, Caturra e outros.
3.3. Hereditariedade da resistência a Hemileia vastatrix em Coffea spp.
Os primeiros estudos sobre a genética da interacção cafeeiro-ferrugem foram realizados por Mayne (1936) na Índia, com interacções de C. arabica e de híbridos C. arabica x C. liberica com as quatro raças de H. vastatrix até então diferenciadas.
Noronha-Wagner e Bettencourt (1967) verificaram que a teoria gene-a-gene de Flor (1942, 1971), segundo a qual para cada gene que condiciona a resistência (R) no hospedeiro existe um gene específico que condiciona a avirulência (Avr) no agente patogénico, se aplica à interacção cafeeiro-ferrugem alaranjada.
Estes estudos de hereditariedade foram continuados no CIFC onde foram identificados, até hoje, nove genes dominantes (SH1 a SH9) que, simples ou associados, conferem resistência de Coffea spp.
as raças de H. vastatrix (Rodrigues Jr. & Bettencourt, 1975; Bettencourt & Rodrigues Jr., 1988). Os genes SH1 a SH5 são provenientes de C. arabica, excepto o SH3 que tem origem em C.
liberica. Todos os restantes genes são provenientes do Híbrido de Timor. A existência de outros genes de resistência em derivados de Híbridos de Timor e outros híbridos interespecíficos tem sido confirmada pela quebra de resistência de alguns desses cafeeiros em relação a novas raças de ferrugem (Rodrigues Jr., 2000).
4. Considerações gerais sobre a resistência das plantas aos
agentes patogénicos
Ao longo da sua evolução, as plantas elaboraram estratégias de reconhecimento e defesa que lhes permitem impedir ou cessar eficazmente a invasão de potenciais agentes patogénicos, ao mesmo tempo que estes desenvolveram novas estratégias de invasão. Apesar deste jogo de ping-pong, verifica-se que na natureza as plantas são resistentes à maioria dos agentes patogénicos sendo a situação de susceptibilidade a excepção (Mysore & Ryu, 2004).
Muitas das defesas das plantas que impedem a invasão dos agentes patogénicos são constitutivas ou pré-existentes. Entre estas defesas destacam-se características da superfície da planta (a topografia da folha, a quantidade e qualidade das ceras presentes na cutícula, o tamanho, a localização e a forma dos estomas, a existência de pêlos, etc.) e ainda a presença de compostos antifúngicos e antimicrobianos (como sejam péptidos, proteínas, compostos fenólicos, alcalóides, saponinas) (Rodrigues Jr., 1980; Heath, 1991; Mysore & Ryu, 2004; Agrios, 2005). Outras defesas são induzidas após a infecção, como a produção de formas reactivas de oxigénio (ROS), a reacção de hipersensibilidade (RH), a acumulação de fitalexinas (compostos antimicrobianos de baixo peso molecular), alterações estruturais ao nível da parede celular da planta, o encapsulamento dos haustórios com calose e a síntese de proteínas relacionadas com a patogenicidade (proteínas PR) (Littlefield & Heath, 1979; Dixon & Lamb, 1990; Skipp et al., 1995; Mysore & Ryu, 2004).
A resistência das plantas aos agentes patogénicos pode ser de dois tipos: resistência do não hospedeiro e resistência do hospedeiro. A resistência do não hospedeiro, também designada por resistência basal, é a forma mais comum e durável de resistência das plantas, sendo expressa por todos os genótipos de uma determinada espécie de plantas a todos os genótipos de uma espécie de agentes patogénicos (Heath, 2000a). A resistência do não hospedeiro aos agentes patogénicos é não específica, geneticamente complexa, envolvendo múltiplos mecanismos de defesa o que pode, pelo menos em parte, contribuir para a sua durabilidade (Heath, 2000a; Hammond-Kosack & Jones, 2003). A resistência do não hospedeiro pode ser do tipo I se não houver sintomas visíveis de necrose ou, do tipo II se houver sintomas de necrose (Mysore & Ryu, 2004).
A resistência do hospedeiro pode ser classificada em resistência vertical e horizontal. A resistência vertical é considerada específica à raça do agente patogénico sendo condicionada por um ou poucos genes frequentemente dominantes que conferem resistência completa à planta relativamente a algumas raças do agente patogénico, mas que são muitas vezes “quebrados” (expressão de susceptibilidade) por outras raças (Agrios, 2005). Esta resistência qualitativa está de acordo com a teoria de gene-a-gene de Flor (1942, 1955). A resistência horizontal é considerada não específica à raça do agente patogénico sendo condicionada por vários genes e caracteriza-se por uma resistência moderada a todas as raças do agente patogénico (Agrios, 2005). Os parâmetros considerados para a avaliação da resistência horizontal são quantitativos, podendo considerar-se entre outros, a frequência de infecção, o período de latência e a intensidade da esporulação (Zhang et al., 1994).
Numa interacção planta-agente patogénico, uma das principais preocupações dos melhoradores é a especificidade e a durabilidade da resistência a incorporar nas cultivares (Niks & Rubiales, 2002). A
enquanto a cultivar que a possui é extensivamente cultivada num ambiente favorável à doença. A resistência horizontal (poligénica) é considerada mais durável do que a resistência vertical (oligogénica), pois a probabilidade de vários genes do agente patogénico sofrerem mutações em simultâneo que lhe permitam ultrapassar a resistência conferida pelo sistema poligénico da resistência horizontal é considerada baixa.
4.1 Reconhecimento do agente patogénico, transmissão de sinal e
respostas de defesa
O sucesso da defesa da planta no combate à invasão do agente patogénico depende em muito da rapidez e intensidade de resposta. Quando as respostas de defesa são tardias e/ou pouco intensas, a planta hospedeira apresenta susceptibilidade ao agente patogénico e a interacção denomina-se compatível. Pelo contrário, quando as respostas de defesa são rápidas e/ou intensas permitem bloquear eficazmente o avanço do agente patogénico, a planta apresenta resistência e a interacção denomina-se incompatível (Laugé & Wil, 1998; Duhoux & Nicole, 2004; van Loon et al., 2006; Bari & Jones, 2009).
O reconhecimento do agente patogénico pela planta é uma etapa crucial no processo de defesa da planta (Peck, 2003). A presença do agente invasor é detectada através do reconhecimento das moléculas do agente patogénico (eliciadores) pelas moléculas receptoras (receptores) da célula hospedeira (Peck, 2003; Agrios, 2005). Os eliciadores são moléculas de baixo peso molecular com diversas estruturas químicas (oligossacáridos, péptidos, proteínas, glicoproteínas e lípidos) que normalmente estão presentes em baixas concentrações. Os eliciadores podem ter vários graus de especificidade: (i) eliciadores específicos, que são o produto do gene Avr reconhecido apenas pelo produto do gene R do hospedeiro, seguindo assim a teoria gene-a-gene de Flor (Peck, 2003); (ii) eliciadores não específicos, que são compostos do próprio organismo invasor, como por ex. fragmentos de quitina da parede celular do fungo, reconhecidos por várias espécies de plantas (Peck, 2003).
O reconhecimento dos eliciadores é feito por receptores específicos localizados, quer na membrana plasmática (Pontier et al., 2002), quer no citoplasma (Dangl & Jones, 2001; Agrios, 2005), sendo assim possível o seu reconhecimento quer no exterior da célula quer já no seu interior (Dangl & Jones, 2001).
Após o reconhecimento do agente patogénico pela planta, são rapidamente accionados mecanismos de transmissão de sinal que resultam em alterações celulares, como produção de formas reactivas de oxigénio (ex. peróxido de hidrogénio (H2O2) eo ião superóxido (O
-2), influxo de Ca2+, activação de cinases e alterações no citoesqueleto e nos níveis de transcrição de genes (Dangl & Jones, 2001; Peck, 2003; Duhoux & Nicole, 2004). Diversos estudos têm referido a importância e regulação de outras enzimas, para além das cinases, no processo de sinalização, tais como as NADPH oxidases, peroxidases apoplásticas e fosfoproteínas (Bestwick et al., 1997; Heath, 2000a).