Caracterização molecular de isolados de nematóides entomopatogênicos, Heterorhabditis...
Texto
Imagem
Documentos relacionados
1 Doutora em Ciências da Comunicação; professora da Universidade Federal do Paraná nos cursos de graduação em Comunicação Social (Publicidade e Propaganda e Relações Públicas)
The aim of this study was to evaluate antioxidant capacity of the methanolic extracts from four cultivated strains and four Internal Transcribed Spacer (ITS)-identified, Iranian
Based on its 16S rRNA gene sequence analysis and its morphological, physiological, and biochemical charac- teristics, strain BJ71 was identified as Cupriavidus campinensis..
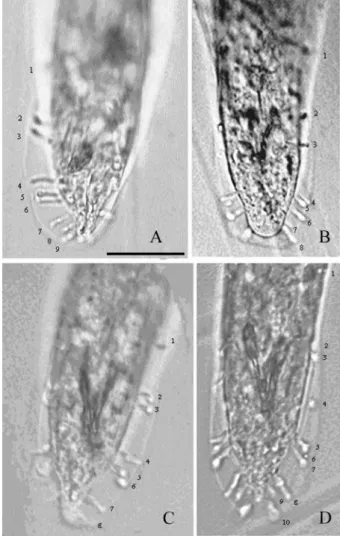
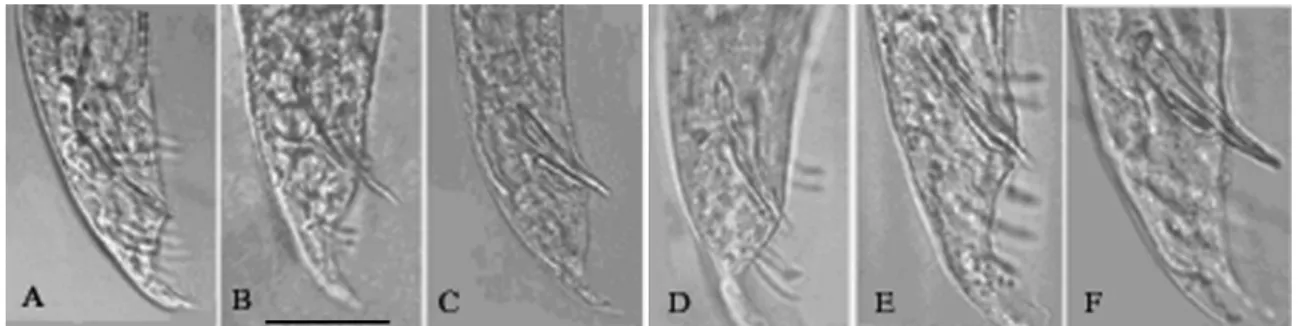
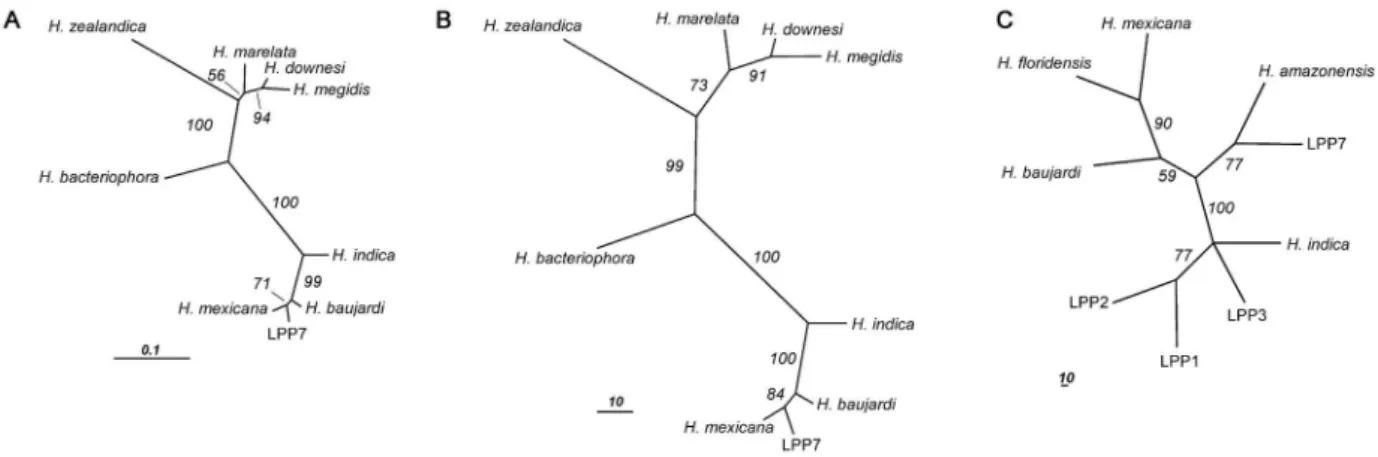
Two species of heterorhabditid nematodes were cited for Argentina: Heterorhabditis argentinensis Stock, 1993 in Santa Fe and La Pampa provinces, and Heterorhabditis bacteriophora
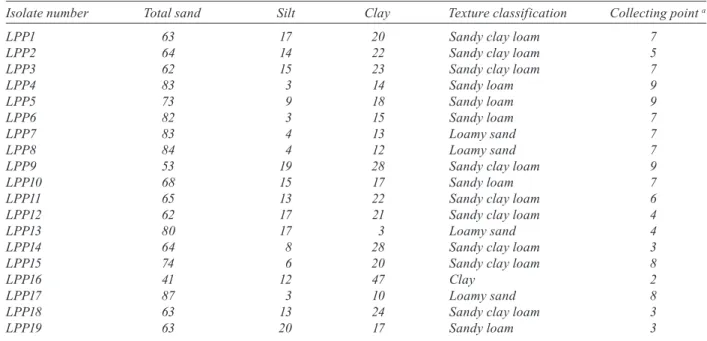
Thus, a molecular characterization of the isolates was performed based on a combined analysis of the following sequences: internal transcribed spacer (ITS) regions 1 and
Este estudo analisou o direcionamento das atividades de lazer desenvolvidas nos Centros de Convivência de Idosos (CCI) nos Municípios da Região Norte do Paraná e a importância
Além disso, a sociedade moderna é dependente em muito dos heterociclos, pois estes compõem produtos sintéticos como fármacos,1 defensivos agrícolas,2 corantes3 e plásticos,4 bem
Para uma melhor compreenção das etapas, estes valores serão analisados em grupos, de acordo com suas características construtivas: grupo comparativo de
