UNIVERSIDADE FEDERAL DO CEARÁ CENTRO DE CIÊNCIAS AGRÁRIAS DEPARTAMENTO DE FITOTECNIA
CURSO DE AGRONOMIA
ANA KARINY DE FREITAS FONTELES
TROCAS GASOSAS E PRODUTIVIDADE EM ACESSOS DE FEIJÃO-CAUPI DO BANCO ATIVO DE GERMOPLASMA DA UNIVERSIDADE FEDERAL DO CEARÁ
ANA KARINY DE FREITAS FONTELES
TROCAS GASOSAS E PRODUTIVIDADE EM ACESSOS DE FEIJÃO-CAUPI DO BANCO ATIVO DE GERMOPLASMA DA UNIVERSIDADE FEDERAL DO CEARÁ
Monografia apresentada ao curso de Agronomia do departamento de Fitotecnia da Universidade Federal do Ceará, como requisito parcial para obtenção do título de Engenheiro Agrônomo.
Orientadora: Profª. Dra. Rosilene Oliveira Mesquita
Coorientadora: Dra. Ana Kelly Firmino
Dados Internacionais de Catalogação na Publicação Universidade Federal do Ceará
Biblioteca Universitária
Gerada automaticamente pelo módulo Catalog, mediante os dados fornecidos pelo(a) autor(a)
F762t Fonteles, Ana Kariny de Freitas.
Trocas gasosas e produtividade em acessos de feijão-caupi do Banco Ativo de Germoplasma da Universidade Federal do Ceará / Ana Kariny de Freitas Fonteles. – 2018.
44 f. : il. color.
Trabalho de Conclusão de Curso (graduação) – Universidade Federal do Ceará, Centro de Ciências Agrárias, Curso de Agronomia, Fortaleza, 2018.
Orientação: Profa. Dra. Rosilene Oliveira Mesquita. Coorientação: Profa. Dra. Ana Kelly Firmino.
1. Vigna unguiculata. 2. estresse hídrico. 3. fotossíntese. I. Título.
ANA KARINY DE FREITAS FONTELES
TROCAS GASOSAS E PRODUTIVIDADE EM ACESSOS DE FEIJÃO-CAUPI DO BANCO ATIVO DE GERMOPLASMA DA UNIVERSIDADE FEDERAL DO CEARÁ
Monografia apresentada ao curso de Agronomia do departamento de Fitotecnia da Universidade Federal do Ceará, como requisito parcial para obtenção do título de Engenheiro Agrônomo.
Orientadora: Profª. Dra. Rosilene Oliveira Mesquita
Coorientadora: Dra. Ana Kelly Firmino
Aprovado em: 20/06/2018.
BANCA EXAMINADORA
____________________________________________________ Profa. Dra. Rosilene Oliveira Mesquita
Universidade Federal do Ceará (Orientadora pedagógica)
____________________________________________________ Dra. Eng. Agrônoma Ana Kelly Firmino da Silva
Universidade Federal do Ceará (Coorientadora)
____________________________________________________ Eng. Agrônomo Johny de Souza Silva
Mestrando da Universidade Federal do Ceará
__________________________________________________ Eng. Agrônomo Rafael Santiago da Costa
A Deus.
A minha família, José Valmir, Ana Maria, Valmir Filho e João Victor.
AGRADECIMENTOS
Primeiramente à DEUS por estar sempre ao meu lado me dando força e sabedoria pra eu superar todos os obstáculos da minha vida.
Aos meus pais, José Valmir e Ana Maria por todo o amor, paciência, apoio e ensinamentos que a mim foi dado por toda a minha vida. Sem vocês eu não seria nada e não estaria aonde cheguei. Amo vocês.
Aos meus irmão Valmir Filho e João Victor por todo apoio e incentivo.
À professora Rosilene Mesquita pela orientação, disponibilidade e confiança.
À Ana Kelly minha coorientadora, por ter me ajudado e compartilhado os seus conhecimentos.
Ao Laboratório de Sementes (LAS) por ter concedido o espaço e a todos os seus integrantes para a realização do experimento e o meu estágio.
Aos meus amigos de trabalho Nájla Falcão e Michell Rocha pela parceria no desenvolvimento do trabalho.
Aos mestrandos Rafael Santiago e Johny de Souza, por toda a ajuda nas avaliações, nas orientações, paciência e principalmente no desenvolvimento escrito do presente trabalho.
A todos os meu amigo, em especial minhas amigas Nájla Falcão, Clara Cordeiro, Wendy Honório e Cícera Mesquita, que ao longo de todo o curso foram sempre parceiras, me ajudaram e me deram forças nos piores momentos. Vocês foram essenciais.
“Lute com determinação, abrace a vida com paixão, perca com classe e vença com ousadia,
porque o mundo pertence a quem se atreve e a
vida é muito para ser insignificante.’
RESUMO
O feijão-caupi é uma leguminosa de origem africana de grande importância para alimentação e geração de emprego e renda para as regiões tropicais e subtropicais do mundo, principalmente nas Regiões Norte e Nordeste onde se concentra o seu cultivo. O estresse hídrico é um dos fatores mais impactantes que afetam o crescimento e as respostas fisiológicas da planta, devido à queda da taxa de fotossíntese, que está relacionada com a limitação das trocas gasosas, por consequência do fechamento dos estômatos. Com isso objetivou-se o presente trabalho a caracterização e avaliação dos parâmetros fisiológicos e a produtividade a partir das avaliações das trocas gasosas de 59 genótipos de feijão-caupi sob regime de sequeiro, identificando os genótipos que são mais tolerantes à seca. O experimento foi conduzido na área de multiplicação do feijão-caupi da Universidade Federal do Ceará. Foram realizadas duas avaliações das trocas gasosas, sendo uma na fase vegetativa e outra na fase reprodutiva, tendo como variáveis a taxa fotossintética (A), condutância estomática (gs), taxa de transpiração (E), eficiência do uso da
água EUA e eficiência instantânea de carboxilização EiC. O delineamento experimental utilizado foi o inteiramente casualizados (DIC) em esquema fatorial 59 x 2, com 59 genótipos e 2 estádios fenológicos, com 3 repetições. Os resultados experimentais foram submetidos à análise de variância e as médias comparadas pelo teste de Scott Knot a 5% de significância. Os resultados demonstraram que em relação a variável fotossíntese (A), condutância estomática (gs) e produtividade, os acessos CE693, CE701 e CE880 obtiveram melhores resultados. Na
variável transpiração (E) os acessos CE693, CE880 e CE570 obtiveram melhores resultados na fase vegetativa e os acessos CE735, CE880 e CE659 na fase reprodutiva. Em relação a variável EUA, os acessos CE109, CE659 e o CE735 obtiveram melhores resultados na fase vegetativa, e os acessos CE701, CE693 e CE109 melhores resultados na fase reprodutiva. Na variável eficiência instantânea de carboxilação (EiC), não houve significância entre as fases. Assim, foi possível concluir, que no geral, o CE880, obteve melhores resultados, sendo o mais indicado para a região semiárida, em seguida os acessos CE701, CE693, CE735.
ABSTRACT
Cowpea is a legume of African origin of great importance for food and employment generation and income for the tropical and subtropical regions of the world, mainly in the North and Northeast regions where its cultivation is concentrated. Water stress is one of the most impacting factors affecting plant growth and physiological responses, due to the decrease in the rate of photosynthesis, which is related to the limitation of gas exchange, due to the closure of the stomata. The objective of this study was to characterize and evaluate the physiological parameters and productivity from the gas exchange evaluations of 59 genotypes of cowpea under rainfed conditions, identifying the genotypes that are more tolerant to drought. The experiment was conducted in the area of cowpea multiplication of the Federal University of Ceará. Two gas exchange evaluations were carried out, one in the vegetative phase and the other in the reproductive phase. The variables were the photosynthetic rate (A), stomatal conductance (gs), transpiration rate (E), US water efficiency and efficiency EiC carboxylation
flash. The experimental design was the completely randomized (DIC) in a factorial scheme 59 x 2, with 59 genotypes and 2 phenological stages, with 3 replicates. The experimental results were submitted to analysis of variance and the means compared by the Scott Knot test at 5% significance. The results showed that, in relation to the variable photosynthesis (A), stomatal conductance (gs) and productivity, accesses CE693, CE701 and CE880 obtained better results.
In the transpiration variable (E) the accessions CE693, CE880 and CE570 obtained better results in the vegetative phase and accesses CE735, CE880 and CE659 in the reproductive phase. In relation to US, accessions CE109, CE659 and CE735 obtained better results in the vegetative phase, and accessions CE701, CE693 and CE109 had better results in the reproductive phase. In the variable instantaneous carboxylation efficiency (EiC), there was no significance between the phases. Thus, it was possible to conclude that, in general, CE880, obtained better results, being the most suitable for the semiarid region, then accesses CE701, CE693, CE735.
LISTA DE ILUSTRAÇÕES
Figura 1- - Imagem de satélite da área de multiplicação de feijão-caupi do Banco Ativo de Germoplasma da UFC onde foi conduzido o experimento ... 27
Figura 2 - Área de multiplicação de feijão-caup onde foi conduzido o experimento, distribuída em fileiras após a semeadura ... 28
Figura 3 - - Avalição das trocas gasosas em genótipos de feijão-caupi do Banco Ativo de Germoplasma da UFC sob regime sequeiro ... 29
Gráfico 1 - Taxa fotossintética na fase vegetativa (FV), fase reprodutiva (FR) e produção por planta (g/planta) em genótipos de feijão-caupi cultivados em regime de sequeiro ... 31
Gráfico 2 - Condutância estomática na fase vegetativa (FV) e fase reprodutiva (FR) em genótipos de feijão-caupi cultivados em regime de sequeiro... 33
Gráfico 3 - Transpiração na fase vegetativa (FV) e fase reprodutiva (FR) em genótipos de feijão-caupi cultivados em regime de sequeiro... 34
Gráfico 4 - Eficiência do uso da água (EUA) na fase vegetativa (FV) e fase reprodutiva (FR) em genótipos de feijão-caupi cultivados em regime de sequeiro ... 35
LISTA DE TABELAS
LISTA DE ABREVIATURAS E SIGLAS
A Taxa fotossintética
BAG Banco Ativo de Germoplasma
CE Ceará
Ci/Ca Razão entre concentração interna e externa de CO2
CONAB Campanha Nacional de Abastecimento
E Taxa de transpiração
EiC Eficiência instantânea de carboxilização. EMBRAPA Empresa Brasileira de Pesquisa Agropecuária EUA Eficiência do uso da água
gs Condutância estomática IRGA Infra-red Gas Analyzer
SUMÁRIO
1 INTRODUÇÃO ... 14
2 REVISÃO DE LITERATURA 2.1 Origem e Importância socioeconômica do Feijão-caupi (Vigna unguiculata (L.) Walp.) ... 16
2.2 Aspectos Taxonômicos, Morfológicos e Fenológicos ... 16
2.3 Exigências edafoclimática e ecofisiológica ... 18
2.4 Estresse hídrico ... 19
2.5 Estudo das trocas gasosas ... 19
2.6 Solos e plantio ... 21
2.7 Manejo e principais doenças e pragas do Feijão-caupi ... 21
2.8 Manejo de plantas daninhas ... 23
2.9 Manejo de irrigação ... 23
2.10 Colheita ... 24
2.11 BAG- Banco Ativo de Germoplasma ... 24
3 MATERIAL E MÉTODO ... 26
4 RESULTADOS E DISCUSSÃO ... 30
5 CONCLUSÕES ... 38
REFERÊNCIAS ... 39
ANEXO A – Quadro de nomes dos genótipos estudados no experimento de acordo com o registro do BAG da Universidade Federal do Ceará ... 43
14
1 INTRODUÇÃO
O Feijão-caupi (Vigna unguiculata (L.) Walp.) é de origem africana, foi introduzido no Brasil pelos colonizadores portugueses no século XVI. A entrada se deu pelo Estado da Bahia, onde se expandiu para todo o País (FREIRE FILHO, 1988). O Brasil é o quarto produtor mundial e sua importância não restringe só no Brasil, mas também inúmeros outros países, principalmente no continente africano (DO VALE et al., 2017). O Feijão-caupi possui uma grande variabilidade genética (FREIRE FILHO et al., 2005). É uma das principais fontes alimentares para as regiões tropicais e subtropicais do mundo, principalmente nas Regiões Norte e Nordeste.
O feijão-caupi tem vários nomes vulgares, como feijão-de-macáçar ou de macaça, feijão-de-corda na Região Nordeste; feijão-da-colônia, feijão-de-praia e feijão-de-estrada na região Norte; e feijão miúdo na Região Sul (FREIRE FILHO, 1988).
Considerando valor do potencial genético do caupi e a necessidade de reconhecer novos genótipos para a sua exploração no pré-melhoramento, faz-se necessária a quantificação da variabilidade genética e, utilização em programas de melhoramento. Para isto, é imprescindível a caracterização e identificação dos genótipos, onde essas atividades são realizadas pela maioria dos pesquisadores que trabalham com feijão (comum e caupi), o que implica tradicionalmente no uso de descritores botânicos, morfológicos e agronômicos (GONÇALVES et al., 2015).
A cultura do feijão-caupi adapta-se bem a diversas condições edafoclimáticas, onde tem predominância em regiões tropicais (TEIXEIRA et al., 2010). O feijão-caupi é uma planta de ciclo curto, baixa exigência hídrica e de fertilidade do solo (DO VALE et al., 2017). Dentre os fatores que afetam o crescimento, comportamento fisiológico e a produtividade dos vegetais, pode-se destacar o estresse hídrico como um dos mais impactante, que é bastante comum na região Nordeste, onde as precipitações se concentram na quadra chuvosa, e apresenta uma alta taxa evaporativa. Dessa forma, o rendimento da cultura é prejudicado, impedindo-o, assim, de alcançar grandes produtividades quando submetidas ao cultivo de sequeiro (GOMES et al., 2000).
Quanto às relações hídricas e trocas gasosas, sabe-se que o influxo de CO2 ocorre
15
disponibilidade hídrica no solo pode causar fechamento estomático limitando o fluxo de CO2 e
a transpiração, o que reduz, consequentemente, a taxa de fotossíntese (SILVA et al., 2010). Boyer (1996) afirma que alguns mecanismos como o controle estomático eficiente pode ser uma forma de redução da perda de água através da transpiração.
As plantas quando submetidas ao estresse hídrico apresentam uma série de mudanças como um menor acúmulo de biomassa, uma menor expansão foliar e consequentemente uma menor área fotossintetizante, acarretando em uma menor produtividade (FARIAS et al., 2011). Vale ressaltar que as elevadas temperaturas do semiárido, quando aliadas ao elevado índice de radiação solar e na concentração de CO2 incidente sobre as plantas, podem influenciar na
formação dos vegetais, repercutindo no seu crescimento e produção (BELTRÃO, OLIVEIRA 2008).
A avaliação das trocas gasosas é uma importante ferramenta na determinação de adaptação e estabilidade de plantas em determinados ecossistemas, isto porque a redução no crescimento e consequentemente da produtividade das plantas pode estar relacionada à redução da taxa fotossintética, limitada por fatores abióticos no local de cultivo (PAIVA et al., 2005).
A partir disso, fica em evidencia a necessidade da realização de trabalhos de melhoramento, visando, sobretudo, à identificação de genótipos adaptados a determinadas condições edafoclimáticas, para que se obtenha um bom resultado no cultivo, aumentando a produção e obtendo maior lucratividade aos produtores.
16
2 REVISÃO DE LITERATURA
2.1 Origem e Importância socioeconômica do Feijão-caupi (Vigna unguiculata (L.) Walp.)
O feijão-caupi (Vigna unguiculata (L.) Walp.) é de origem africana, foi introduzido no Brasil pelos colonizadores portugueses no século XVI. A entrada se deu pelo Estado da Bahia, onde se expandiu para todo o País (FREIRE FILHO, 1988). O Brasil é o quarto produtor mundial e sua importância não restringe só no Brasil, mas também inúmeros outros países, principalmente no continente africano (DO VALE et al., 2017). O Feijão-caupi possui uma grande variabilidade genética (FREIRE FILHO et al., 2005). É uma das principais fontes alimentares para as regiões tropicais e subtropicais do mundo, principalmente nas Regiões Norte e Nordeste.
O caupi é importante para alimentação e fornecimento de emprego e renda. É rico em proteína, minerais e fibras e constitui um componente alimentar básico das populações rurais e urbanas das regiões Norte e Nordeste (FROTA et al., 2008; SINGH, 2007 apud FREIRE FILHO, 2011). De acordo com Dutra (2007), o cultivo do feijão-caupi é de grande importância para o desenvolvimento agrícola da região Nordeste, tanto econômico como no nutricional, devido ser um alimento básico na alimentação da populações mais pobres.
O caupi tem vários nomes vulgares, como macassar ou de macássar, feijão-de-corda na Região Nordeste; feijão-da-colônia, feijão-de-praia e feijão-de-estrada na região Norte; e feijão miúdo na Região Sul (FREIRE FILHO, 1988).
Segundo a Campanha Nacional de Abastecimento (CONAB, 2018), em 2017/18, a área de cultivo com feijão-caupi em sequeiro está em torno de 611,4 mil ha, concentrada na Região Norte e Nordeste. No mês maio de 2018, a produção nacional chegou a 1.533,1 mil toneladas.
2.2 Aspectos Taxonômicos, Morfológicos e Fenológicos
17
O feijão-caupi possui germinação epígea. A emergência das plântulas ocorre entre dois e três dias após a semeadura, caso as sementes esteja na profundidade (2 a 3 cm) e temperatura ambiente (por volta de 28°C) (WIEN et al., 1984 apud FREIRE FILHO et al., 2005).
O sistema radicular é composto por: raiz principal e a pivotante com ramificações nas laterais, que podem crescer até mais de dois metros. A raiz principal e as secundárias é comum encontrar nódulos que auxiliam na absorção de nitrogênio através da fixação biológica de nitrogênio, realizada por bactérias nitrificadoras do gênero Rhizobium (CHAGAS JUNIOR
et al., 2009).
O caule é uma haste herbácea, ramificada e de cor verde. A arquitetura da planta em feijão-caupi é bastante variável entre e dentro dos cultivares (ARAÚJO, 1979). Segundo Freire Filho et al. (2005), os vários tipos de porte apresentados foram classificados em ereto, semi-ereto, semiprostrado e prostrado, variando quanto ao número de nós e de ramificações.
A folha do feijão-caupi é composta e contém três foliolos presos a uma haste denominada de pecíolo. O primeiro par de folhas acima dos cotilédones, folhas primárias, são sésseis, simples e opostas, exibindo considerável variação de forma e tamanho. O folíolo terminal é utilizado usualmente para classificar a forma da folha, que apresenta uma grande importância para a classificação taxonômica e identificar os diferentes genótipos (DO VALE
et al., 2017).
O feijãp caupi é classificado quanto a forma da folha em quatro tipos: subglobosa, sublanciolada, globosa, e lanciolada. O folíolo terminal é frequentemente comprido e de maior área que os folíolos assimétricos laterais, as formas variam de linear lanceoladas a ovaladas (POTTORFF et al., 2012 apud DO VALE et al., 2017).
A multiplicação do feijão-caupi ocorre, predominantemente, por autofecundação devido estrutura da flor. Os órgãos reprodutivos são protegidos pelas pétalas e ocorre protoginia (maturação do gineceu antes do androceu) e cleistogamia (polinização antes da abertura da flor), favorecendo a autogamia. A planta de feijão-caupi apresenta uma pequena taxa de cruzamento natural que varia com o ambiente e com o genótipo (DO VALE et al., 2017).
18
O cálice é pentâmero e o gamossépalo varia a coloração de verde a roxa. A corola é pentâmera ou dialipétala. A pétala maior é o estandarte e as duas laterais são as asas, onde ambas podem apresentar a coloração branca, violeta ou outros diferente tons de violeta. As asas cobrem a quilha (duas pétalas inferiores unidas e recurvadas) de coloração branca (DO VALE
et al., 2017). No interior da quilha encontram-se os órgãos femininos e masculinos. (ROCHA et al., 2007 apud DO VALE et al., 2017).
O fruto é do tipo vagem de tamanho, forma, cor e número de sementes variável, as sementes são disposta em fileiras ao longo da vagem, podendo variar a menos de 10 e mais que 30 cm de comprimento. A forma da vagem no ponto de colheita pode ser classificada em: reta cilíndrica, reta achatada, curva cilíndrica e curva achatada e a coloração pode ser amarela, rosada, roxa e rajada. O número de semente por vagem variam de menos de 10 e mais de 20 (DO VALE et al., 2017).
A semente do feijão-caupi é composto pelo embrião e tegumento e, apresentam uma grande diversidade de cores, de formas, onde são classificadas em: reniforme, globosa, ovoide e romboide. O seu tamanho pode variar de 10 a 30 g por 100 sementes. O seu tegumento pode ser rugoso ou liso (DO VALE et al., 2017).
As condições climáticas em determinados locais, podem influenciar nas variações dos estádios fisiológicos do feijão-caupi (DO VALE et al., 2017). Segundo Do Vale et al (2017), existem duas fases diferentes: vegetativa e reprodutiva. A fase vegetativa é compota por 10 estádios e a reprodutiva por 5 (CAMPOS et al., 2000).
Os ciclos do feijão-caupi pode ser classificados em: extraprecoce (< 30 dias), precoce (de 31 a 40 dias), médio (de 41 a 50 dias), tardio (de 51 a 60 dias e extratardio (> 60 dias) (PAIVA et al., 2014).
2.3 Exigências edafoclimática e ecofisiológica
19
A temperatura influência no desenvolvimento vegetativa, temperaturas inferiores a 18ºC desestimulam o início da floração, aumentando assim o ciclo da cultura (RIOS et al., 1984). Do Vale et al. (2017) destaca, que alguns fatores climáticos (abióticos) podem prejudicar o crescimento e desenvolvimento da planta, tais como: precipitação, fotoperíodo, vento, radiação solar, salinidade, concentração de nutrientes minerais e principalmente a temperatura.
2.4 Estresse hídrico
O estresse hídrico, pode ser definido como um fator extrínseco, que exerce uma influência negativa sobre a planta. O conceito de estresse hídrico está relacionado ao de tolerância ao estresse de um ambiente desfavorável. Um ambiente estressante para uma planta pode não ser para outra, ou seja, a tolerância à seca varia de espécie para espécie e mesmo dentro de uma espécie, entre variedades (LUDLOW, 1976 apud SANTOS,2016). Com o passar dos anos e da evolução, as plantas passaram por várias mudanças na sua estrutura e no processo fisiológico, na qual as tornaram adaptáveis a sobreviver em ambientes secos.
Um dos primeiros efeitos do déficit hídrico manifesta-se sobre os estômatos. Com a progressão do dessecamento, ocorre a desidratação do protoplasma e a redução da capacidade fotossintética. A deficiência hídrica e a queda da taxa de fotossíntese pode ser relacionada com a limitação das trocas gasosas, por consequência do fechamento dos estômatos devido a diminuição no potencial hídrico da folha. (SOUZA et al., 2001).
Também em decorrência ao déficit hídrico, Sousa et al (2009) verificaram que o efeito do estresse hídrico em feijoeiro causa reduções nos componentes de produção e é mais severo quando ocorre nas fases de floração e frutificação.
2.5 Estudo das trocas gasosas
As trocas gasosas são processos pelos quais as plantas interagem com a atmosfera liberando ou captando gases. Deste modo, durante a fotossíntese, a planta fixa CO2 e libera O2,
o inverso ocorre na respiração, ou seja, a planta libera CO2 e utiliza O2, revertendo assim as
trocas desses gases. Esse balanço de entrada e saída de CO2 é possível de ser detectado
medindo-se as trocas gasosas, sendo a fotossíntese líquida o processo em que mais CO2 é
20
As trocas gasosas entre as células e a atmosfera ocorrem pelo processo de difusão, que está diretamente ligado ao gradiente de concentração estabelecido em um determinado momento. O trabalho de assimilação fotossintética é melhorado com o aumento da concentração interna do CO2. No entanto, em condições naturais a concentração de CO2 entre
o meio externo e o interior das plantas é muito reduzido, sendo assim, a baixa concentração de CO2 é o fator mais comum para a limitação das taxas fotossintética (SILVA, 2015).
O mecanismo de abertura e fechamento estomático exerce papel essencial para a entrada de gases, que influencia o processo de ganho fotossintético, uma vez que é pela variação na abertura estomática que a planta controla o influxo de CO2 na planta. Diferentes fatores
ambientais influenciam a abertura e fechamento dos estômatos e, consequentemente, a condutância estomática (LARCHER, 2006). Estresses causados pelo ambiente podem provocar o fechamento estomático, queda da condutância estomática e consequente redução da concentração interna de CO2 e da taxa fotossintética (ALVES et al., 2011).
A transpiração também é afetado pelo déficit hídrico o que interfere na produtividade das plantas, pois esta auxilia na translocação de compostos dentro da célula, sendo um importante fator no resfriamento dos tecidos. O resultado do estresse hídrico sobre a transpiração produz significativas diferenças entre plantas sob deficiência hídrica e irrigadas (ALVES et al., 2011).
Com a queda da disponibilidade de água no solo a transpiração diminui, devido à perda de água enquanto os estômatos estão abertos. A vegetação e o estádio de crescimento também são fatores que modificam a taxa de transpiração. Quando o solo está sob condição adequadas de umidade, a transpiração se matem numa taxa potencial, determinada pelas condições meteorológicas. Outros fatores tais como temperatura, umidade relativa, correntes de vento também podem influenciar as taxas de transpiração (KLAR, 1988).
21
irrigação e de seus efeitos sobre o rendimento das culturas (ALI et al., 2007; KARAM et al., 2007).
A eficiência instantânea de carboxilação (EiC) é adquirida a partir da relação entre a concentração intracelular de CO2 (Ci) e a fotossíntese líquida (A), assim possibilitando a
identificação de fatores que interferem na fotossíntese (MACHADO et al., 2005).
Portanto, a análise das características relacionadas às trocas gasosas é uma ferramenta valiosa para correlacionar a influência da restrição hídrica na assimilação de CO2
pelas plantas.
2.6 Solos e plantio
O feijão-caupi pode ser cultivado em diversos tipos de solo, principalmente os latossolos amarelos, latossolos vermelho-amarelo, neossolos flúvicos. Desenvolvem-se em solos com regular teor de matéria orgânica, soltos, leves e profundos, arejados e dotados de média a alta fertilidade. Outros solos com baixa fertilidade também podem ser utilizados, como Latossolos e Neossolos Quartzarenicos, mediante de aplicações de fertilizantes químicos e/ou orgânicos (EMBRAPA, 2002).
“O feijão-caupi é cultivado em todo o território brasileiro principalmente no Nordeste e Norte, onde se encontra os mais variados métodos de plantio, desde o mais rudimentar até a motomecanização com plantadeiras adubadeiras.” (EMBRAPA, 2002).
A escolha adequada do cultivar é um processo de muita importância para o produtor, pois uma cultivar adaptada a região do cultivo é fundamental para obter uma produção de qualidade. Na escolha da cultivar devem-se considerar a região do cultivo, adaptabilidade, o sistema de plantio, manejo, porte, qualidade da semente, a resistência a fatores bióticos e abióticos e valor nutricional, de modo que atenda as exigências do mercado (DO VALE et al., 2017).
2.7 Manejo e principais doenças e pragas do Feijão-caupi
Existem vários fatores que limitam a qualidade da produção, uma delas são as doenças, que provocam grandes perdas e danos na produção do feijão-caupi (RIBEIRO, 2002). O feijão-caupi pode ser atacados por uma diversidade de agentes patogênicos, causando várias doenças, como: Fungos (Antracnose, Carvão-da-folha, Cercosporiose, Ferrugem, Murcha de
22
Bactérias (Crestamento-bacteriano), Vírus (Mosaico-dourado, Mosaico-leve, Mosaico de
Potyvirus eMosaico-severo) e Nematoides (Meloidoginose) (DO VALE et al., 2017).
A ação desse organismos podem levar à morte dos tecidos, órgãos e da planta inteira, causando grandes perdas econômicas (DO VALE et al., 2017). É importante ressaltar, que a cultura do feijão-caupi praticamente não dispõe de agroquímicos registrados para uso corrente no sistema de manejo de doenças, restando apenas como recurso àqueles relacionados com as medidas de exclusão, erradicação não química, evasão e controle genético (EMBRAPA, 2016).
Além das doenças, existem outros fatores que podem causar danos na cultura do caupi, que são as pragas. “Os insetos causadores de danos na cultura do feijão-caupi aparecem de acordo com o período de desenvolvimento da planta.” (NEVES et al., 2011). O ataque desses insetos ocorrem geralmente quando a planta está no estadio fenológico produzindo o alimento ideal para eles (FREIRE FILHO et al., 2005).
As pragas do feijão-caupi são agrupadas de acordo com o local do ataque na planta, na seguinte forma: pragas subterrâneas, pragas da parte aérea (folhas e órgãos reprodutivos) e pragas de armazenamento (FREIRE FILHO et al., 2005).
Em cada ciclo fenológico há ocorrência de pragas, como: Germinação (Paquinha); Fase vegetativa (Paquinha (Neocurtilla hexadactyla), Lagarta elasmo (Elasmoplapus
lignosellus), Lagarta-rosca (Agrotis ípsilon), Larvas de vaquinha (Diabrotica speciosa), Vaquinhas (Diabrotica speciosa), Lagartas desfolhadoras (Spodoptera cosmioides), Cigarrinhas (Enpoasca kraemeri), Pulgão (Toxoptera citricida), Mosca-branca (Bemisia
tabaci) e Minador-das-folhas) e Fase reprodutiva (Vaquinhas (Diabrotica speciosa), Lagartas desfolhadoras (Spodoptera cosmioides), Lagarta das vagens (Etiella zinknella), Cigarrinhas (Enpoasca kraemeri), Pulgão (Toxoptera citricida), Mosca-branca (Bemisia tabaci), Mosca
minadora (Liriomyza huidobrensis), Percevejos (vermelho- Crinocerus sanctus, pequeno- Piezodorus guildini, verde- Nezara viridula), Manhoso (Chaldocermus bimaculatus), Carunchos (Callosobruchus maculatus) e Helicoverpa armigera (EMBRAPA, 2016).
23
e Abastecimento – MAPA, para aplicação nessa cultura. Para a agricultura familiar, o controle mais adequado é o manejo cultural (EMBRAPA, 2016).
2.8 Manejo de plantas daninhas
“Existem várias definições para planta daninhas, porém uma das mais frequentes são aquelas que ocorrem em locais não desejados pelo homem.” (DO VALE et al., 2017). Algumas ações podem ser mais indicadas para o controle de plantas daninhas no cultivo do feijão-caupi, que são elas: a preventiva, cultural e mecânica. O objetivo da ação preventiva é impedir a entrada e o estabelecimento de plantas daninhas nas lavouras por meio de limpeza de máquinas, implementos e ferramentas agrícolas, obtenção de sementes certificadas ou fiscalizadas.
A ação cultural ocorre a partir do manejo adequado do solo (sistema plantio direto ou preparo convencional), adubações de semeadura e em cobertura (quando for o caso), populações de plantas indicadas para cada cultivar, controle de pragas e manejo de irrigação. O controle mecânico é realizado por meio manual (capinas) ou tratorizado (cultivadores) (FONTES, 2016). O controle químico não pode ser indicado para o feijão-caupi, pois não existem produtos comerciais registrados para controle daninhas (EMBRAPA, 2002).
2.9 Manejo de irrigação
O feijão-caupi tem um adaptabilidade moderada às condições contrarias de solo, clima e sistema de cultivo, no entanto o rendimento é baixo quando comparado com outras plantas. Porém, quando é submetido ao uso da irrigação, a alta produtividade de grãos pode ser obtida (CARDOSO et al.,1996 apud FREIRE FILHO, 2005).
O feijão caupi tem potencial genético para alcançar produtividades elevadas quando em cultivo irrigado, porém a deficiência de água é um dos fatores mais limitantes da produção de grãos, por quanto o estresse hídrico prejudica diversas processos fisiológicos da planta (MENDES et al., 2007). Considerando que diferentes genótipos de feijão caupi respondem de formas distintas à aplicação de água e às condições climáticas nas quais estão submetidas (RAMOS et al., 2014).
24
extensão da área da cultura, a disponibilidade de água, a topografia e assistência técnica (FREIRE FILHO et al., 2005).
De acordo com Freire Filho (2005), a irrigação é um elemento essencial para um bom crescimento e desenvolvimento do caupi. No entanto, é necessário um manejo racional de irrigação, pois o excesso e a escassez podem afetar o desenvolvimento da planta.
2.10 Colheita
A etapa da colheita é de fundamental importância para a produção de sementes, e deve ser feita de forma eficiente para evitar perdas. A colheita pode ser manual (mais utilizada), semimecanizada ou mecânica (DO VALE et al., 2017). “A colheita deve ser realizada logo que a lavoura atinja o ponto de maturidade adequado, estágio R5.” (CAMPOS et al., 2000). Após a colheita, as vagens devem ser colocadas para secar, pois apresentam um teor de água quando recém-colhidas, que podem causar a perda do vigor e poder germinativo no período de armazenamento (FREIRE FILHO et al., 2005). É importante que seja feito o expurgo do material no armazém para controlar as pragas associadas às sementes, principalmente o caruncho (Callosobruchus maculatus Febr.), que causa grandes perdas físicas e de qualidade em feijão-caupi. (EMBRAPA, 2002).
As sementes que vão ser comercializadas devem estar bem limpas e padronizadas, quanto ao tamanho. É importante que o beneficiamento seja feito em máquina apropriada e bem regulada para evitar danos às sementes. As sementes podem ser armazenadas de diferentes formas, de acordo com os produtores. Em pequenas propriedades o feijão-caupi é armazenado em garrafas de vidro, em recipientes com camadas de areia fina, latas de flandres e tambores de zinco; nas médias e grandes propriedades, as sementes são armazenadas em tambores de zinco e silos metálicos (EMBRAPA, 2002).
25
2.11 Banco Ativo de Germoplasma (BAG)
Considerando a importância do feijão-caupi na alimentação tradicional da população, um grupo de especialistas de Vigna discutiram a importância da conservação destes recursos (FREIRE, 1999). Segundo Araújo (1988) os programas de melhoramento do caupi, no Brasil, começaram em 1963, por diversas instituições de pesquisa das regiões Norte e Nordeste.
A maior parte dos recursos genéticos de feijão-caupi está disponível em bancos de germoplasma e coleções de trabalho. Os trabalhos de coleta, introdução, caracterização, avaliação, regeneração e intercâmbio do germoplasma são feitos nos bancos de germoplasma. Por isso, os acessos necessitam ser descritos adequadamente (SOBRAL, 2009).
A coleção ativa faz a conservação a curto e médio prazo, para gestão e distribuição (FREIRE FILHO et al., 2009). Freire Filho et al. (2009) mencionam, que os recursos genéticos necessários para o melhoramento genético de novas linhagens e cultivares, encontram-se disponíveis em quatro tipos de coleções de germoplasma: a coleção de base internacional, que está localizado na Nigéria, no Instituto Internacional de Agricultura Tropical (IITA); coleção de base nacional, localizada no Centro Nacional de Recursos Genéticos e Biotecnologia – CENARGEN, em Brasília no Distrito Federal; a coleção ativa, que no caso do feijão-caupi, há duas coleções ativas, onde uma está localizada na Embrapa Meio-Norte ou Centro de Pesquisa Agropecuária do Meio-Norte (CPAMN, em Teresina-PI) e a outra está no Centro de Ciências Agrárias da Universidade Federal do Ceará, Fortaleza, Ceará; e a coleção de trabalho, onde existem três localizadas, uma na Embrapa Meio-Norte, no Instituto Agronômico de Pernambuco – IPA e na Embrapa Semiárido.
O banco ativo da Universidade Federal do Ceará foi iniciado em 1963 pelo professor José Braga Paiva no departamento de Fitotecnia, que hoje pertence ao Centro de Ciências Agrarias da UFC. Atualmente o banco tem mais de 900 acessos registrados (PAIVA
26
3 MATERIAL E MÉTODOS 3.1 Localização do experimento
O experimento foi conduzido na área de multiplicação de feijão-caupi pertencente ao Departamento de Fitotecnia do Centro de Ciências Agrárias da Universidade Federal do Ceará (UFC), no período de Fevereiro a Junho de 2018, com altitude de 47 m, latitude Sul 3º 44’ 27,1’’ e longitude Oeste 38º 34’ 39’’. (Figura 1). A temperatura média na área nesse intervalo de tempo foi de 26°C, de acordo com o CLIMATE-DATA.ORG.
Figura 1- Imagem de satélite da área de multiplicação de feijão-caupi do Banco Ativo de Germoplasma da UFC onde foi conduzido o experimento.
Fonte: Elaborado pela autora utilizando o Google Maps.
3.2 Instalação e Condução do experimento
O experimento foi implantado em de Fevereiro 2018, onde foram utilizadas 59 variedades integrante do Banco Ativo de Germoplasma de Feijão-Caupi da Universidade Federal do Ceará.
27
regularmente com o auxílio de um facão, essa pratica é o método mais comum para o controle de plantas daninhas.
Durante o ciclo da cultura, foi realizado o monitoramento da cultura, no qual foi percebido o surgimento de doenças como virose e carvão, e de pragas como Percevejo vermelho (Crinocerus sanctus), Pulgão (Toxoptera citricida), Vaquinha (Diabrotica speciosa) e Manhoso (Chaldocermus bimaculatus), onde foi realizado o controle manual e controle alternativo, quando necessário, com agroquímicos seguindo as recomendações para aplicação dos mesmos.
Figura 2 - Área de multiplicação de feijão-caupi onde foi conduzido o experimento distribuída em fileiras após a semeadura.
Fonte: Elaborado pela própria autora.
3.3 Caracterização morfoagronômica
A caracterização morfoagronômica é uma das principais procedimentos realizados no Banco Ativo de Germoplasma. Devido os genótipos utilizados no presente trabalho, serem acessos de renovação, foi extrema importância fazer essa caracterização. Para a realização desse procedimento, foi necessário a utilização da metodologia descrita por Paiva et al., 2014.
28
A segunda etapa foi realizada em laboratório, onde foi feita a caracterização das vagens quanto a sua cor, forma, comprimento e número de sementes por vagem; e a caracterização da semente quanto a cor, forma, textura e peso de 100 sementes.
3.4 Trocas gasosas
O entendimento das trocas gasosas nas plantas em relação com o estresse hídrico é parte fundamental para a escolha de cultivares tolerantes. Compreender as resposta fisiológica das plantas, diante de fatores ambientais como a escassez de água no solo, temperaturas elevadas, entre outros, deixou de ser visto como uma simples questão fisiológica ou ecológica, mas como uma importante ferramenta para avaliar ou minimizar os problemas sociais e econômicos.
As avaliações das trocas gasosas foram realizadas a partir do terceiro ou quarto trifólio completamente expandido e, as medições foram realizadas entre 08:00 e 12:00 a.m., As variáveis avaliadas foram: taxa de assimilação fotossintética líquida por unidade de área (A), condutância estomática (gs), taxa de transpiração (E), razão entre concentração interna e externa
de CO2 (Ci/Ca), eficiência do uso da água (EUA; (A/E)) e eficiência instantânea de
carboxilização (EiC; (A/Ci)). As variáveis foram determinadas por analisador de gás por infravermelho (IRGA; modelo portátil LI-6400XT, LI-COR Biosciences Inc., Lincon, Nebraska, USA), utilizando radiação fotossinteticamente ativa (PAR) constante (1200 μmol fótons m-2 s-1), concentração constante de CO
2 (400ppm), temperatura e umidade ambiente.
29
Figura 3 - Avalição das trocas gasosas em genótipos de feijão-caupi do Banco Ativo de Germoplasma da UFC sob regime sequeiro.
Fonte: Elaborado pela própria autora. 3.5 Delineamento experimental e análise estatística
O delineamento experimental utilizado foi o inteiramente casualizados (DIC) em esquema fatorial 59 x 2, com 59 genótipos e 2 fases de desenvolvimento (vegetativa e reprodutiva) com 3 repetições, totalizando 354 parcelas experimentais.
30
4 RESULTADOS E DISCUSSÃO
As trocas gasosas são processos pelos quais as plantas interagem com a atmosfera liberando ou captando gases. Deste modo, durante a fotossíntese, a planta fixa CO2 e libera O2,
o inverso ocorre na respiração, ou seja, a planta libera CO2 e utiliza O2, revertendo assim as
trocas desses gases. Esse balanço de entrada e saída de CO2 é possível de ser detectado medindo-se as trocas gasosas, sendo a fotossíntese líquida o processo em que mais CO2 é consumido na fotossíntese do que liberado pela respiração simultaneamente. Essa variável é naturalmente reduzida no escuro ou quando a planta está sob condições desfavoráveis para a assimilação (LARCHER, 2006).
O entendimento das troca gasosas nas plantas em relação com o estresse hídrico é parte fundamental para a escolha de cultivares tolerantes, além de ser uma ferramenta que determina a possível adaptação e estabilidade de plantas a determinados ecossistemas (FERRAZ et al., 2012).
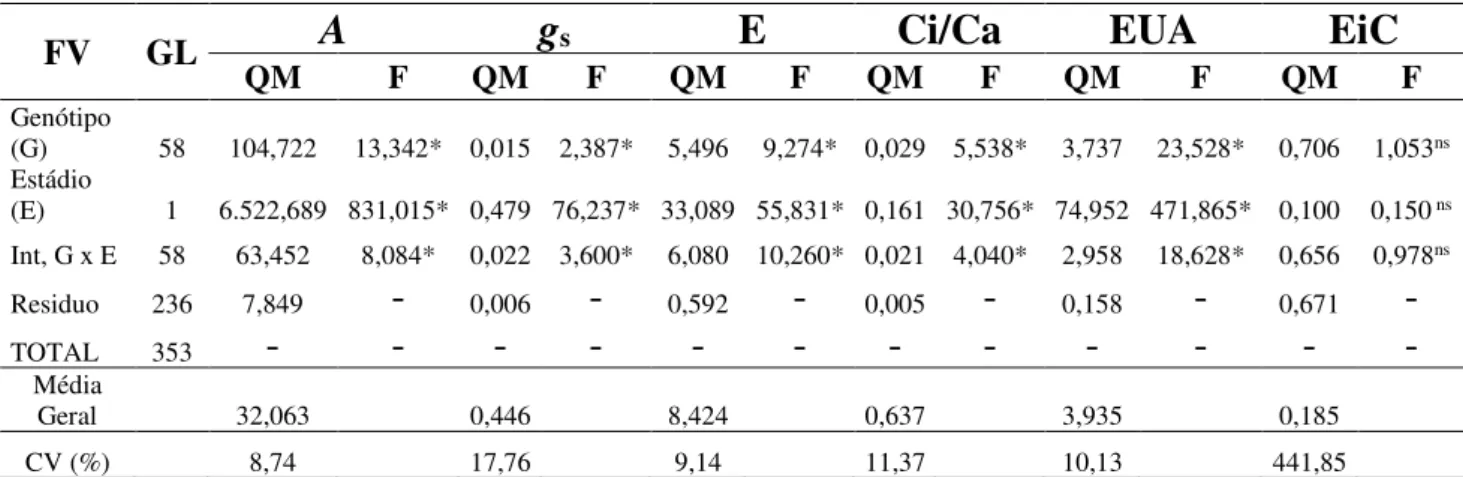
Conforme o resumo da análise de variância para trocas gasosas, apresentados na Tabela 1, podemos observar que houve diferenças significativas dos fatores isolados, genótipos e estádios fenológicos, como também da interação (genótipos x estádios), para todas as variáveis estudadas, exceto a variável EiC.
Tabela 1 - Resumo das análises de variância para as variáveis (A), (gs), (E), (Ci/Ca), (EUA), (EiC), analisadas nas
duas fases fenológicas (vegetativa e reprodutiva) de genótipos de feijão-caupi.
FV GL
A
g
sE
Ci/Ca
EUA
EiC
QM F QM F QM F QM F QM F QM F
Genótipo
(G) 58 104,722 13,342* 0,015 2,387* 5,496 9,274* 0,029 5,538* 3,737 23,528* 0,706 1,053ns Estádio
(E) 1 6.522,689 831,015* 0,479 76,237* 33,089 55,831* 0,161 30,756* 74,952 471,865* 0,100 0,150 ns Int, G x E 58 63,452 8,084* 0,022 3,600* 6,080 10,260* 0,021 4,040* 2,958 18,628* 0,656 0,978ns
Residuo 236 7,849
-
0,006-
0,592-
0,005-
0,158-
0,671-TOTAL 353
-
-
-
-
-
-
-
-
-
-
-
-Média
Geral 32,063 0,446 8,424 0,637 3,935 0,185 CV (%) 8,74 17,76 9,14 11,37 10,13 441,85
Sigla: FV - Fontes de variação; GL - Graus de liberdade; CV - Coeficientes de variação; A - Fotossíntese; gs -
Condutância estomática; E– Transpiração; Ci/Ca - Razão entre concentração interna e externa de CO2; EUA –
Eficiência de uso da água; EiC - Eficiência intrínseca de carboxilação; * - Significativo a 5% pelo teste F e ns= não significativo.
31
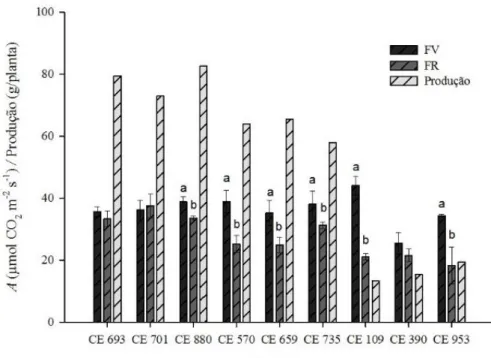
discussão dos resultados, adotou-se como parâmetro de escolha, produtividade e fotossíntese na fase reprodutiva. Os acessos utilizados foram: os melhores (CE693, CE701 e CE880), os medianos (CE570, CE659 e CE735) e os piores (CE109, CE390 e CE953).
4.1 Fotossíntese (A) e Produtividade
No gráfico 1, podemos observar os acessos CE693, CE701 e CE880, que apresentaram as maiores taxas fotossintéticas durante a fase reprodutiva. Para esses acessos, a fotossíntese líquida oscilou entorno de 40 μmol CO2 m-2 s-1, e consequentemente apresentou
melhor produtividade. Já os acessos CE570, CE659 e CE735, foram selecionados por apresentar valores medianos às taxas fotossintética durante a fase reprodutiva, com assimilação de CO2 em torno de 25 a 35 μmol CO2 m-2 s-1, já a produtividade foi em média 60 g/planta. Os
acessos com baixa taxa fotossintética e produção foram: CE109, CE390 e CE953, apresentaram assimilação de carbono em torno de 20 a 25 μmol CO2 m-2 s-1, resultando em uma menor
produtividade.
Gráfico 1. Taxa fotossintética na fase vegetativa (FV), fase reprodutiva (FR) e produção por planta (g/planta) em genótipos de feijão-caupi cultivados em regime de sequeiro.
Fonte: Elaborada pela autora.
32
A redução da disponibilidade hídrica é um fator comum na região Nordeste, que por apresentar clima semiárido, sofre constante situações de déficit hídrico durante o ciclo das culturas, o que ocasiona redução na produtividade (FERNANDES et al., 2015). Além disso, baixa taxa de precipitação fazem com que as plantas utilizem mecanismos de defesa, entre eles, o fechamento estomático, afim deevitar a perda de água pela transpiração. Como consequência, dessa ação, há um menor influxo de CO2 para o mesófilo foliar, havendo assim uma queda na
atividade fotossintética (BOSCO et al., 2009).
Em condições naturais, a concentração de CO2 entre o meio externo e o interior das
plantas é muito reduzido, sendo assim, a baixa concentração de CO2 é o fator mais comum para
a limitação das taxas fotossintética (SILVA, 2015).
Resultado semelhante foi encontrado por Silva et al. (2010), ao acompanhar o curso diário das trocas gasosas em plantas de feijão (Vigna unguiculata) sob deficiência hídrica, em que evidenciaram efeito significativo das lâminas de irrigação sobre os aspectos fisiológicos, encontrando, reduções nas taxas de fotossíntese, condutância estomática e transpiração. Portanto, a ocorrência de estresses abióticos, mas, principalmente, por falta de água, acarreta séries de alterações fisiológicas desfavoráveis aos vegetais, como observados no presente trabalho.
Alguns estudos tem demostrado efeito significativo do déficit hídrico sobre a fotossíntese, como por exemplo o estudo realizado por Souza et al. (2004) que observaram que após a imposição do estresse hídrico, plantas de feijão caupi tiveram menores rendimentos nas trocas gasosas. Corroborando com os dados encontrados na presente pesquisa, Rivas et al. (2016), também encontram redução da fotossíntese em razão de um estresse por seca.
4.2 Condutância estomática (gs)
33
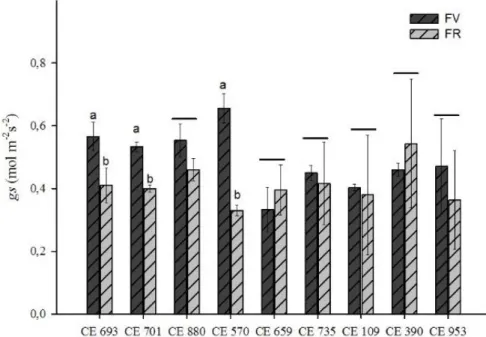
Gráfico 2. Condutância estomática na fase vegetativa (FV) e fase reprodutiva (FR) em genótipos de feijão-caupi cultivados em regime de sequeiro.
Fonte: Elaborada pela autora
Condutância estomática dos acessos na fase vegetativa são indicados pela barra escura, na fase reprodutiva pela barra clara. A médias foram comparadas através do teste Scott Knot a 5% de significância. As médias que apresentarem um traço, não diferem entre fases.
As plantas perdem água para o meio externo ao absorverem CO2, essa perda ocorre
através dos estômatos, que apresentam um controle da abertura e fechamento dos estômatos, que é atribuído a condutância estomática (MC DERMIT, 1990). A baixa disponibilidade de água no solo ocasiona a queda no potencial hídrico nas folhas das plantas, reduzindo a turgescência da planta e consequentemente a condutância estomática (SHALHEVET, 1983).
Nesse estudo, foi evidenciado que a queda na fotossíntese se deve principalmente ao fechamento estomático. Nessa pesquisa os acessos CE693, CE701 e CE880, foram os que apresentaram maiores médias de condutância estomática, como consequência das maiores médias de fotossíntese. Essa série de reações, provavelmente elevou a taxa produtiva.
Em geral, nota-se que na fase vegetativa, ocorreu maior abertura estomática. Nessa fase as plantas requerem energia pra crescer e se desenvolver. A abertura estomática é responsável pelo o influxo de CO2 na planta e, quando controlada, afeta o processo
34
semelhantes nos genótipos BRS Paraguaçu e Pingo-de-ouro-1,2, quando avaliou a tolerância ao déficit hídrico em genótipos de feijão-caupi, onde apresentaram maiores valores na condutância estomática, em condições de deficiência hídrica, que demonstraram eficiência no controle estomático, permitindo uma alta atividade fotossintética. Souza et al. (2011) verificaram que as plantas de feijão-caupi sob estresse salino apresentaram menor taxa de assimilação de carbono e justificaram que isso foi ocasionado pela redução da abertura dos estômatos. Silva et al (2016), também relataram que houve diferença significativa na condutância estomática quando analisado interação do genótipo x técnica, no estudo do crescimento e trocas gasosas de genótipos de feijão-caupi sob estratégias de cultivo.
4.3 Transpiração (E)
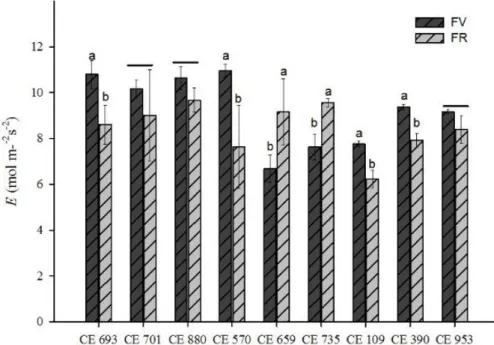
Observou-se no gráfico 3, que os acessos CE693, CE880 e CE570 obtiveram uma maior taxa de transpiração na fase vegetativa e os acessos CE735, CE880 e CE659 tiveram maior destaque na fase reprodutiva. Já o acesso CE880 não apresentou diferença significativa quando analisados a interação entre as duas fases.
Gráfico 3. Transpiração na fase vegetativa (FV) e fase reprodutiva (FR) em genótipos de feijão-caupi cultivados em regime de sequeiro.
Fonte: Elaborada pela autora
35
A transpiração também é afetada pelo déficit hídrico o que interfere na produtividade das plantas,pois esta auxilia na translocação de compostos dentro da célula, sendo um importante fator no resfriamento dos tecidos. Com a queda da disponibilidade de água no solo a transpiração diminui, devido à perda de água, enquanto os estômatos estão abertos.
Outros fatores também podem influenciar as taxas de transpiração, tais como temperatura, umidade relativa e correntes de vento. Quando a perda de água é maior que a absorção, pode ocorrer o retardamento do crescimento das plantas e até mesmo a morte em muitas, por desidratação.
Nesse trabalho, observa-se que em geral na fase vegetativa, houve maior taxa transpiratória. Com a queda da disponibilidade de água no solo a transpiração diminui, devido à perda de água enquanto os estômatos estão abertos.
Machado et al. (2005) enfatizam que havendo menores aberturas estomáticas haverá a diminuição da transpiração, com consequente aumento da temperatura foliar. Silva et al (2013), ao estudar Trocas gasosas em plantas de girassol submetidas à deficiência hídrica em diferentes estádios fenológicos, observaram que as plantas no estádio vegetativo ocorreu uma maior transpiração, como no presente trabalho, indicando que a maioria das plantas neste estádio fenológico a cultura possui maior demanda hídrica.
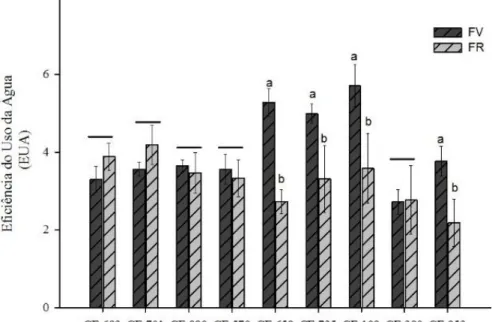
4.4 Eficiência do uso da água (EUA)
36
Gráfico 4. Eficiência do uso da água (EUA) na fase vegetativa (FV) e fase reprodutiva (FR) em genótipos de feijão-caupi cultivados em regime de sequeiro.
Fonte: Elaborada pela autora
Eficiência do uso da água (EUA) dos acessos na fase vegetativa são indicados pela barra escura, na fase reprodutiva pela barra clara. A médias foram comparadas através do teste Scott Knot a 5% de significância. As médias que apresentarem um traço, não diferem entre fases.
A eficiência do uso da água (EUA) é estipulada pela relação entre fotossíntese (A) e transpiração (E). Alta taxa fotossintética com baixa concentração de carbono pode induzir o fechamento estomático, reduzindo o processo de transpiração e obtendo maior eficiência no uso da água.
As plantas quando submetidas ao estresse prolongado, tendem a economizar água de todas as formas possíveis como estratégia de sobrevivência. Uma das estratégia envolve o fechamento estomático que está entre as primeiras respostas à seca, protegendo as plantas contra uma perda de água excessiva.
37
de indicadores como o EUA tem se tornado uma das formas mais práticas de se analisar a resposta dos cultivos às diferentes condições de disponibilidade hídrica (SOUZA et al., 2011).
Oliveira (2016), ao estudar a caracterização fisiológica e bioquímica de variedades locais de feijão-caupi para tolerância a seca no estado do Ceará, obteve os seguintes resultados, com a imposição de déficit hídrico do genótipo Pingo de Ouro 1,2 apresentou uma maior EUA, no entanto, a cultivar Meia Corda apresentou menores valores de eficiência.
4.5 Eficiência instantânea de carboxilação (EiC)
A eficiência instantânea de carboxilação (EiC) é estipulada a partir da relação entre a concentração intracelular de CO2 (Ci) e a fotossíntese líquida (A), assim possibilitando a
identificação de fatores que interferem na fotossíntese (MACHADO et al., 2005). Na variável EiC no presente trabalho, foi observado que não houve diferença significativa entre as fases fenológica.
Dutra et al (2015), verificaram que houve diferenças significativas para as variáveis: transpiração (E), condutância estomática (gs), fotossíntese (A), eficiência instantânea
38
5 CONCLUSÕES
O acesso CE880 apresentou melhores resultados para as variáveis: A, gs e E, no
entanto não houve diferença significativa da EUA entre as fases de desenvolvimento. Além do CE880, os acessos CE701, CE693 e CE735 respectivamente, mantiveram boas respostas dentro dos parâmetros avaliados para cultivo em sequeiro. Esses acessos produziram bem e podem ser genótipos indicados para regiões com restrição hídrica, como a região semiárida do Nordeste.
No entanto, pode- se destacar, que os acessos CE109, CE390 e CE953, obtiveram resultados não satisfatórios para as variáveis estudadas, sendo assim mais sensíveis a restrição hídrica sendo os menos indicados ao cultivo em sequeiro.
39
REFERÊNCIAS
AMARAL, J. A. T. do; RENA, A. B.; AMARAL, J. F. T. do. Crescimento vegetativo sazonal do cafeeiro e suas relações com fotoperíodo, frutificação, resistência estomática e
fotossíntese. Pesquisa Agropecuária Brasileira. v. 41, n. 3, p. 377-384, 2006. BELTRÃO, N. E. de M.; OLIVEIRA, M. I. P. Oleaginosas e seus óleos: vantagens e desvantagens para produção de biodiesel. Embrapa Algodão. Documentos 201, 2008. BOYER, J. S. Advances in drought tolerance in plants. Advances in Agronomy, v. 56, p. 189-218, 1996
BOYER, J. S. Water deficits and photosynthesis. In: Kozlowski, T.T. (ed.) Water deficits and plant growth. New York: Academic Press, v.4, p.154-191, 1978.
CAMPOS, F. L.; FREIRE FILHO, F. R.; LOPES, A. C. de A.; RIBEIRO, V. Q.; SILVA, R. Q. B.; ROCHA, M. de M. Ciclo fenológico em caupi (Vigna unguiculata (L.)walp): uma proposta de escala de desenvolvimento. Revista Científica Rural. v.5, n.2, p.110-116, 2000. CARDOSO, M. J. (Org.). A cultura do feijão caupi no Meio Norte do Brasil. Embrapa Meio-Norte, Circular Técnica 28, p.264, 2000.
CARDOSO, M. J.; RIBEIRO, V. Q. 2006. Desempenho agronômico do feijão-caupi, cv. Rouxinol, em função de espaçamentos entre linhas e densidades de plantas sob regime de sequeiro. Revista Ciência Agronômica, v. 37, n. 01, p.102-105.
COELHO, D.S.; MARQUES, M. A. D.; BEZERRA, J. A. da S.; GARRIDO, M. da S.; CARVALHO, P. G. S. Respostas fisiológicas em variedades de feijão caupi submetidas a diferentes níveis de sombreamento. Revista Brasileira de Biociências, p. 14-19, 2014. DE ARAÚJO, J. P. P.; RIOS, G. P.; WATT, E. E.; DAS NEVES, B. P.; FAGERIA, N. K.; OLIVEIRA, I. P.; GUIMARÃES, C. M.; FILHO, A. S. Cultura do caupi, Vigna unguiculata (L.) walp; Descrição e recomendações de cultivo. Embrapa Arroz e Feijão. Circular técnico n. 18, p. 44, 1984.
DO VALE, J. C.; BERTINI, C.; BORÉM, A. Feijão-caupi do plantio à colheita. 1 ed. Minas Gerais: Universidade Federal de Viçosa. 2017.
DUTRA, A. S.; TEÓFILO, E. M.; MEDEIROS FILHO, S.; DIAS, F. T. C. Qualidade fisiológica de sementes de feijão caupi em quatro regiões do estado do Ceará. Revista Brasileira de Sementes, vol. 29, nº 2, p.111-116, 2007.
E SILVA, K. J. D.; ROCHA, M. de M.; MENEZES JÚNIOR, J. A. N.; DA SILVA, P. H. S.; SOBRINHO, C. A.; FONTES, J. R. A. A cultura do feijão-caupi no Brasil. Teresina: Embrapa Meio Norte, p. 68, 2016.
EMBRAPA. A cultura do feijão-caupi no Brasil. 1. ed. Teresina: Embrapa Meio Norte, 2016.
EMBRAPA. Sistema de produção 2- Cultura do feijão-caupi (Vigna unguiculata (L.)
walp). 1. ed. Teresina: Embrapa Meio Norte, p. 17-32, 2002.
40
FREIRE FILHO, F. R. Genética no feijão-caupi. In: ARAÚJO, J. P. P.; WATT, E. E., de. O Feijão-caupi no Brasil. Brasília. IITA/EMBRAPA – CNPAF, p.159-229, , 1988.
FREIRE FILHO, F.R.; LIMA, J.A de A.; RIBEIRO, V.Q. Feijão-caupi Avanços tecnológicos. (1 ed.), 2005.
FREIRE FILHO, F.R.; RIBEIRO, V.Q.; BARRETO, P.D.; SANTOS, C.A.F. Melhoramento genético de Caupi (Vigna unguiculata (L.) Walp.) na Região do Nordeste. In: QUEIROZ, M.A.; GOEDERT, C.O.; RAMOS, S.R.R. (Ed.). Recursos genéticos e melhoramento de plantas para o Nordeste brasileiro. Petrolina: EmbrapaCPATSA; Brasília, DF: Embrapa-Cenargen, 1999.
GOMES, A.A.; ARAÚJO, A.P.; ROSSIELLO, R.O.P; PIMENTEL, C. Acumulação de biomassa, características fisiológicas e rendimento de grãos em cultivares de feijoeiro irrigado e sob sequeiro. Pesquisa Agropecuária Brasileira, 35(10), 1927-1937, 2000
KLAR, A.E. Evapotranspiração. In: A água no sistema solo-planta-atmosfera. 2ª ed. São Paulo: Nobel, 1988. Guanagara Koogan, p. 720, 2001.
LARCHER, W. Ecofisiologia vegetal, São Carlos, SP: Rima artes, p. 531, 2006.
LEMOS, L. B. et al. Características agronômicas e tecnológicas de genótipos de feijão do grupo comercial carioca. Pesquisa Agropecuária Brasileira. v. 39, n. 4, p. 319-326, 2004. LONGUI, B. M. Fotossíntese e relações hídricas de espécies de Eucalyptus sp. submetidas a diferentes regimes hídricos. In: Dissertação apresentada à Universidade do Oeste Paulista ao Programa de Pós- Graduação em Produção Vegetal, 2007.
MENDES, R. M. S.; TÁVORA, F. J. A. F.; PINHO, J. L. N.; PITOMBEIRA, J. B. Relações fonte-dreno em feijão-de-corda submetido à deficiência hídrica. Revista Ciência
Agronômica, v. 38, p. 95-103, 2007.
NEVES, A. C.; CÂMARA, J. A. da S.; CARDOSO, M. J.; DA SILVA, P. H. S.; SOBRINHO, C. A. Cultivo do Feijão-caupi em Sistema Agrícola Familiar. Teresina: Embrapa Meio-Norte. Circular técnico, n. 51, p. 7, 2011.
PAIVA, A. S. et al. Condutância estomática em folhas de feijoeiro submetido a diferentes regimes de irrigação. Engenharia Agrícola. v. 25, n. 1, p. 161-169, 2005.
PAIVA, J. B.; FREIRE FILHO, F. R.; TEÓFILO, E. M.; RIBEIRO, V.Q. Feijão-caupi: melhoramento genético no Centro de Ciências Agrárias. Fortaleza: UFC, p. 30, 2014. RAMOS, H. M. M; BASTOS, E. A.; CARDOSO, M. J.; RIBEIRO, V. Q.; NASCIMENTO, F. N. Produtividade de grãos verdes do feijão-caupi sob diferentes regimes hídricos.
Engenharia Agrícola, v. 34, n. 4, p. 683-694, 2014.
RAVEN P.H.; EVERT, R.F.; EICHHORN, S.E. Movimento de água e soluto nas plantas. In: Biologia Vegetal. 6ª ed.
SALVESTRO, A. C.; BRADÃO, D.; DE FREITAS, P. S. L. Métodos de determinação da fotossíntese no cultivo do feijoeiro. In: VII EPCC - Encontro Internacional de Produção Científica, 2011.
41
SILVA, F. L. S. Efeitos de estresses bióticos sobre os parâmetros ecofisiológicos e
componentes de produção de quatro variedades de cana-de-açúcar. Dissertação (Mestrado em Agricultura e Biodiversidade) – Universidade Federal de Sergipe. 2015.
SOBRAL, P. V.C. Caracterização morfoagronômica e divergência genética entre acessos africanos de Feijão-caupi. Dissertação (Mestrado) – Universidade Federal do Piauí, 2009. TEIXEIRA, I. R.; SILVA, G. C.; OLIVEIRA, J. P. R.; SILVA, A. G.; PELÁ, A. Desempenho agronômico e qualidade de sementes de cultivares de feijão-caupi na região do cerrado. Revista Ciência Agronômica, v. 41, n. 2, p. 300-307, 2010.
TEÓFILO, E. M., DUTRA, A.S., PITIMBEIRA, J. B., DIAS, F. T. C. & BARBOSA, F. S. Potencial fisiológicos de sementes de feijão caupi produzidas em duas regiões do estado do Ceará. Revista Ciência Agronômica, 39(3):443-448, 2008.
Mc DERMIT, D.K. Sources of error in the estimation of stomatal conductance and
transpiration from porometer data. HortScience, Alexandria, v.25, n.12, p.1538-48, 1990. RIVAS, R. et al. Drought tolerance in cowpea species is driven by less sensitivity of leaf gas exchange to water deficit and rapid recovery of photosynthesis after rehydration. South African Journal of Botany, v. 103, p. 101–107, 2016.
SOUZA, R. P. et al. Photosynthetic gas exchange , chlorophyll fluorescence and some associated metabolic changes in cowpea ( Vigna unguiculata ) during water stress and recovery. v. 51, p. 45–56, 2004.
SOUZA, B. DE et al. Eficiência do uso da água das culturas do milho e do feijão-caupi sob sistemas de plantio exclusivo e consorciado no semiárido brasileiro. 2011.
NASCIMENTO, S. P., BASTOS, E. A., ARAÚJO, E. C. E., FREIRE FILHO, F.R., SILVA, E.M. Tolerância ao déficit hídrico em genótipos de feijão-caupi. Revista Brasileira de Engenharia Ambiental. V. 15, 2011.
FERNANDES, F, B. P.; LACERDA, C. F.; ANDRADE, E. M., NEVES, A. L. R.; SOUSA, C.H.C. Efeito de manejo do solo no déficit hídrico, trocas gasosas e rendimento do feijão-de-corda no semiárido. Revista Ciência Agronômica. V. 46, n.3, p. 506-515, 2015.
ALVES, A. A.; GUIMARÃES, L. M. da S.; CHAVES, A. R. de M.; DAMATTA, F. M.; ALFENAS, A. C. Leaf gas exchange and chlorophyll a fluorescence of Eucalyptus urophylla in response to Puccinia psidii infection. Acta Physiologiae Plantarum, v. 33, p. 1831-1839, 2011.
KLAR, A.E. Evapotranspiração. In: A água no sistema solo-planta-atmosfera. 2ª ed. São Paulo: Nobel, 1988.
42
GONÇALVES, J. G. R.; CHIORATO, A.F.; DA SILVS, D. A.; ESTEVES, J. A. de F.; BOSETTI, F.; CARBONELL, S. A. M. Análise da capacidade combinatória em feijoeiro comum submetido ao déficit hídrico. Revista de Melhoramento Genético. V. 74. N. 2, p. 149-155, 2015
CHAGAS JUNIOR, A. F.; OLIVEIRA, L. A.; OLIVEIRA, A. N.; WILLERDING, A. L.Efetividade de rizóbios e caracterização fenotípica dos isolados que nodulam feijão-caupi em solos da Amazônia Central. Acta Amazônica vol. 39 p. 489 – 494. 2009.
SOUZA, C. R.; SOARES, A. M.; REGINA, M. de A. Trocas gasosas de mudas de videira, obtidas por dois porta-enxertos, submetidas à deficiência hídrica. Pesquisa agropecuária brasileira. V.36, n. 10, p. 1221-1230, 2001.
ALI, M.H.; HOQUE, M.R.; HASSAN, A.A.; KHAIR, A. Effects of deficit irrigation on yield, water productivity and economic returns of wheat. Agricultural water management, v.92, p.151-161, 2007.
KARAM, F.; LAHOUD, R.; MASAAD, R.; KABALAN, R.; BREIDI, J.; CHALITA, C.; ROUPHAEL, Y. Evapotranspiration, seed yield and water use efficiency of drip irrigated sunflower under full and deficit irrigation conditions. Agricultural water management, v.90, p.213-223, 2007.
FERRAZ, R. L. de S.; MELO, A. S.; SUASSUNA, J. F.; BRITO, M. E. B.; FERNANDES, P. D.; JÚNIOR, E. da S. N. Trocas gasosas e eficiência fotossintética em ecótipos de feijoeiro cultivados no semiárido. Pesquisa Agropecuária Tropical. v. 42, n. 2, p. 181-188, 2012. SOUZA, R. P.; MACHADO, E. C.; SILVEIRA, J. A. G.; RIBEIRO, R. V. Fotossíntese e acúmulo de solutos em feijoeiro caupi submetido à salinidade. Pesquisa Agropecuária Brasileira. v. 46, n. 6, p. 586-592, 2011.
DA SILVA, A.; BRITO, M. E. B.; FRADE, L. J. G.; NOBRE, R. G.; DA COSTA, F. B.; DE MELO, A. S.; SILVA, L. de A. Crescimento e trocas gasosas de genótipos de feijão-caupi sob estratégias de cultivo. Revista Ambiente Água. v.11, n.3, 2016.
MACHADO, E. C.; MEDINA, C. L.; GOMES, M. de M. de A.; HABERMAN, G. Variação sazonal da fotossíntese, condutância estomática e potencial da água na folha de laranjeira ‘Valência’. Revista Scientia Agricola, v. 59, n. 01, p. 53-58, 2002.
SILVA, A. R. A.; BEZERRA, F. M. L.; LACERDA, C. F.; PEREIRA FILHO. V.; FREITAS, C. A. S. Trocas gasosas em plantas de girassol submetidas à deficiência hídrica em diferentes estádios fenológicos. Revista Ciência Agronômica. v. 44, n. 1, p. 86-93, 2013.
DUTRA, A. F.; DE MELO, A. S.; FILGUEIRAS, L. M. B.; DA SILVA, A. R. F.; DE OLIVEIRA, I. M. Parâmetros fisiológicos e componentes de produção de feijão-caupi cultivado sob deficiência hídrica. Revista Brasileira de Ciências Agrárias. v.10, n.2, p.189-197, 2015
OLIVEIRA, J. V. Caracterização fisiológica e bioquímica de variedades locais de feijão caupi para tolerância a seca no estado do Ceará. 2016. 65f. Monografia (Graduação em
43
ANEXOS
Anexo A – Quadro de nomes dos genótipos estudados no experimento de acordo com o registro do BAG da Universidade Federal do Ceará.
REGISTRO CULTIVAR REGISTRO CULTIVAR
CE-003 Vinagre-1 CE-579 CE-315 x CE-1 CE-024 Cowpea-535 CE-581 CE-315 x CE-1
CE-099 Floricrean CE-602 -
CE-109 Lot7909-aik-83 CE-627 CNCx 163-18F
CE-118 Precoce CE-659 CNCx 190-2E/P
CE-121 - CE-688 CNCx 666-29E
CE-125 - CE-693 CNCx 666-41E
CE-129 539 CE-695 CNCx 666-44E
CE-149 1249 CE-701 CNCx 666-51
CE-153 1558 CE-735 CNCx 660-7E
CE-167 2913 CE-753 CNCx 679-50E
CE-186 - CE-767 TVx 5050-07C
CE-197 Coleção Pernambuco-V-3 CE-789 CNCx 251-4E-
CE-202 V-14 CE-821 [016014(03)(01)]
CE-206 V-33 CE-880 CNCx 249-308F-1
CE-223 V-Chiapas 275 CE-881 CNCx 249-313F
CE-245 TVu 57 CE-897 CNCx 333-35E
CE-352 Vita-5 (TVu 4557) CE-902 -
CE-355 Vita-3 (TVu 1190) CE-903 CNCx 333-55E
CE-357 SVS-3 CE-906 CNCx 333-66E
CE-367 TVx 289-4 G CE-953 IT 82D-60
CE-369 TVx 1843-1 C CE-956 UCR 95-701 CE-372 TVx 1836-90 E CE-965 MNC-06-887B-835
CE-390 - CE-966 TVu 3961
CE-457 TVu-2769 CE-968 TE 97-309G-4
CE-470 TVu3509 CE-969 Pampo
CE-545 S-107 CE-987 IT 81D-1064
CE-566 CE-31 x CE-25 CE-990 IT 82E-49 CE-570 CE-315 x CE-31 CE-1007 MNC-01-625D