Arq. Gastroenterol. vol.47 número1
Texto
Imagem
Documentos relacionados
Os Editores das principais revistas médicas do país, diretores da ABEC (Associação Brasileira de Editores Cientíicos) e os coordenadores das áreas de Medicina II e Medicina III
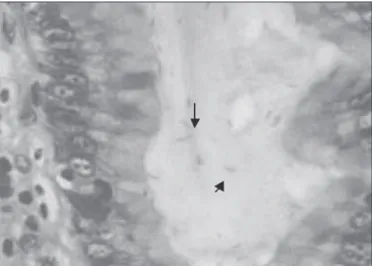
Infection with Helicobacter pylori strains lacking dupA is associated with an increased risk of gastric ulcer and gastric cancer development.. Helicobacter pylori homB , but Not
The purpose of this study was to determine endemic and epidemic infection due to Gram-negative bacilli, risk factors associated with colonization and infection by these organisms
In the present study, smoking was associated with the risk of gastric cancer for both former smokers and current smok- association of smoking and alcohol
Women with human immunodeficiency virus (HIV) infection present a higher risk of infection by the human papil- lomavirus (HPV) and cervical cancer.. To determine HPV genotypes
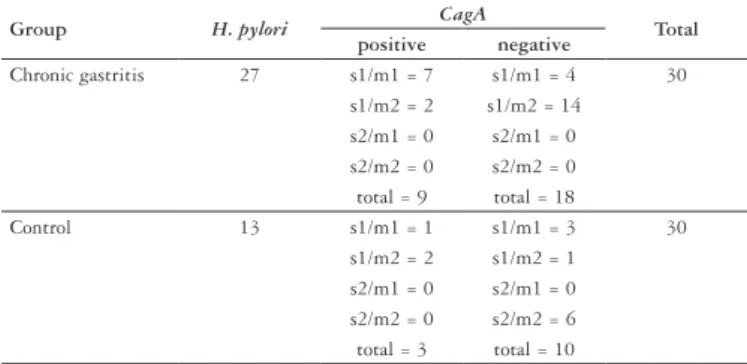
IL-1ra anti-inflammatory cytokine polymorphism is associated with risk of gastric cancer and chronic gastritis in a Brazilian population, but the
The objective of this study was to investigate the prevalence of anti-Neospora caninum antibodies in cattle from milk producing farms of the microregion of Batalha, state of Ala-
To observe effect of E2F decoy DNA on proliferation and apoptosis of androgen-independent prostate cancer cell line PC-3M the binding specificity of the E2F decoy DNA to
