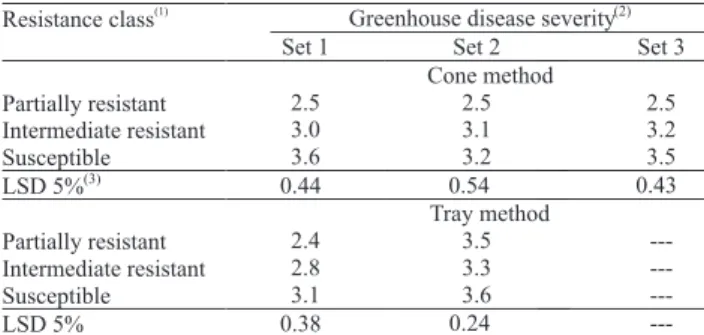
Inoculation methods under greenhouse conditions for evaluating soybean resistance to sudden death syndrome
Texto
Imagem


Documentos relacionados
Para a análise da relação das variáveis independentes com as variáveis dependentes, para além da análise documental, nomeadamente publicações periódicas, artigos de
Progress curves of disease incidence (A) and disease severity (B) of the Asian Soybean Rust on soybean plants inoculated under greenhouse conditions with uredospores of the fungus
5 - Area under disease progress curves measured by the incidence (A) or the severity (B) of soybean (Glycine max) stem canker in different soybean cultivars and plant densities.
Thus, three experiments were conducted: pathogen inoculation of different leaf types ( B, D and F ) of pineapple (1), pathogen inoculation of pineapple cuttings and detached D
Abstract – The objective of this work was to determine soybean resistance inheritance to Heterodera glycines Ichinohe (soybean cyst nematode – SCN) races 3 and 9, as well as
Three additional QTL for resistance to SDS (one in each linkage groups g, n, and c2) were identified by using RIL derived from 'Pyramid' × 'Douglas' (Njiti et al., 2002). These
Table 2 - Analysis of variance of F 2 plants (crosses) and parents, in hills, for the traits number of days to maturity.. (NDM), grain yield (GY), oil content (OC) and oil
includens in soybean genotypes, in no-choice test, in greenhouse and number of trichomes obtained in 16 mm 2 and colorimetric evaluation of leaves of soybean genotypesC. 1
