UNIVERSIDADE FEDERAL DO CEARÁ CENTRO DE CIÊNCIAS AGRÁRIAS DEPARTAMENTO DE FITOTECNIA
CURSO DE AGRONOMIA
ELANE BEZERRA DA SILVA
DIVERGÊNCIA GENÉTICA E CARACTERIZAÇÃO DE ACESSOS DOS GÊNEROS
Habranthus E Hippeastrum (Amaryllidaceae)
ELANE BEZERRA DA SILVA
DIVERGÊNCIA GENÉTICA E CARACTERIZAÇÃO DE ACESSOS DOS GÊNEROS
Habranthus E Hippeastrum (Amaryllidaceae)
Monografia apresentada ao Curso de Agronomia do Centro de Ciências Agrárias da Universidade Federal do Ceará, como requisito parcial à obtenção do Título de Bacharel em Agronomia.
Orientadora: Profª. D.Sc. Cândida Hermínia Campos de Magalhães Bertini.
Coorientadora: D.Sc Ana Cecilia Ribeiro Castro
S579d Silva, Elane Bezerra da.
Divergência genética e caracterização de acessos dos gêneros Habranthus e Hippeastrum (Amaryllidaceae) / Elane Bezerra da Silva. – 2018.
59 f. : il. color.
Trabalho de Conclusão de Curso (graduação) – Universidade Federal do Ceará, Centro de Ciências Agrárias, Curso de Agronomia, Fortaleza, 2018.
Orientação: Profa. Dra. Cândida Hermínia Campos de Magalhães Bertini. Coorientação: Profa. Dra. Ana Cecilia Ribeiro Castro.
ELANE BEZERRA DA SILVA
DIVERGÊNCIA GENÉTICA E CARACTERIZAÇÃO DE ACESSOS DOS GÊNEROS
Habranthus E Hippeastrum (Amaryllidaceae)
Monografia apresentada ao Curso de Agronomia do Centro de Ciências Agrárias da Universidade Federal do Ceará, como requisito parcial à obtenção do Título de Bacharel em Agronomia.
Aprovada em: 15 / 06 / 2018.
BANCA EXAMINADORA
__________________________________________
Profa. D.Sc Cândida Hermínia Campos de Magalhães Bertini (Orientadora) Universidade Federal do Ceará (UFC)
__________________________________________ Pesq. D.Sc Ana Cecília Ribeiro de Castro (Coorientadora)
Empresa Agroindústria Tropical
__________________________________________ Pesq. D.Sc Ana Cristina Portugal Pinto de Carvalho
Empresa Agroindústria Tropical
__________________________________________ Pesq. D.Sc Rita de Cássia Alves Pereira
AGRADECIMENTOS
Primeiramente a Deus e ao universo por me concederem oportunidades de aprendizado, por ter conhecido pessoas incríveis e por ter vivenciado experiências únicas durante essa jornada.
À Universidade Federal do Ceará, pela oportunidade de realização do curso de graduação.
À Embrapa Agroindústria Tropical, pelo espaço físico e material concedido durante a realização do trabalho.
A Dra. Ana Cecília Ribeiro Castro, pela a orientação, conselhos e ensinamentos. A Professora Cândida Bertini pela orientação, revisões e comentários, tudo foi de enorme ajuda.
Aos demais membros da banca examinadora, Doutora Ana Cristina Portugal Pinto de Carvalho e Doutora Rita de Cássia Alves Pereira pelas sugestões e contribuições para este trabalho.
Ao Doutor Fernando Aragão pelo auxílio nas análises e discussão dos resultados. A todos os professores do curso de Agronomia da Universidade Federal do Ceará. Aos colegas de Embrapa pela ajuda nos momentos em que não conseguia dar conta das coisas sozinha.
A Suziane pelas ideias, conversas e direções na construção desse trabalho, obrigada também pela ajuda em algumas disciplinas.
Ao Gérffeson por toda paciência, apoio, carinho e conhecimento compartilhado. A todo pessoal do Ministério da Agricultura, Pecuária e Abastecimento, Leandro, Ewérton, Ana Kátia, Pedro Igor e Beatriz.
Aos queridos amigos que fiz durante o curso, Monique, Lero, Jean, Fabiano, Nathaniel, Suane, Isabelle, Celly e Thais, o curso não teria sido o mesmo sem vocês.
“De tudo, ficaram três coisas: a certeza de que
ele estava sempre começando, a certeza de que era preciso continuar e a certeza de que seria interrompido antes de terminar. Fazer da interrupção um caminho novo. Fazer da queda um passo de dança, do medo uma escada, do
sono uma ponte, da procura um encontro.”
RESUMO
As bulbosas ornamentais de maior importância ornamental são as espécies da família Amaryllidaceae, elas possuem grande apelo comercial, sendo a conservação de sua diversidade natural, uma prioridade. A caracterização e o estudo da diversidade genética são importantes na gestão e no uso do germoplasma conservado, gerando dados que podem ser utilizados em trabalhos de melhoramento genético. A partir disto, os objetivos deste estudo foram: caracterizar e estimar a divergência genética entre acessos dos gêneros Habranthus e
Hippeastrum da coleção de germoplasma da Embrapa Agroindústria Tropical e indicar os
indivíduos com maior potencial para uso ornamental como planta de vaso em condições de Fortaleza. Foram avaliados 29 acessos de Habranthus e 23 de Hippeastrum selecionados da
coleção de germoplasma da Embrapa Agroindústria Tropical. As avaliações foram realizadas com base nos descritores: comprimento de folhas (CFO), largura de folhas (LFO) e por atribuição de notas foram avaliados a taxa de recobrimento do solo (TRS), ocorrência de plantas daninhas (OPD), ataque de insetos (AIN) e índice de folhas secas (FSE). Os dados foram analisados utilizando o método de agrupamento de UPGMA para a avaliação da similaridade genética. As análises de componentes principais foram utilizadas para a seleção dos descritores mais importantes. Foram formados seis grupos no dendrograma de
Habranthus. Através do gráfico de componentes principais foi possível observar que os
descritores que mais contribuíram para caracterização foram taxa de recobrimento do solo, comprimento de folhas, índice de folhas secas e largura de folhas são os que melhor caracterizam este gênero. No dendrograma de Hippeastrum foram formados sete grupos.
Através do gráfico de componentes principais foi possível observar que os descritores que mais contribuíram para caracterização foram taxa de recobrimento do solo, comprimento de folhas, índice de folhas secas e largura de folhas são os que melhor caracterizam este gênero. Conclui-se que os descritores utilizados contribuíram para a caracterização dos acessos de
Habranthus e Hippeastrum, determinando também a divergência genética entre os mesmos.
ABSTRACT
The ornamental bulbous of greater ornamental importance are the species of the family Amaryllidaceae, they have great commercial appeal, being the conservation of its natural diversity, a priority. The characterization and the study of genetic diversity are important in the management and use of conserved germplasm, generating data that can be used in breeding work. From this, the objectives of this study were: to characterize and estimate the genetic divergence between accesses of the genus Habranthus and Hippeastrum from the
Embrapa Agroindústria Tropical germplasm collection and to indicate the individuals with the greatest potential for ornamental use as a pot plant under Fortaleza conditions. Were evaluated 29 accessions of Habranthus and 23 of Hippeastrum selected from the germplasm collection
of Embrapa Tropical Agroindustry. The evaluations were carried out based on the descriptors: leaf length (CFO), leaf width (LFO) and by assignment of grades were evaluated the soil cover rate (TRS), weed occurrence (OPD), insect attack (AIN) and dry leaves index (FSE). The data were analyzed using the UPGMA grouping method for the evaluation of genetic similarity. Principal component analyzes were used to select the most important descriptors. Six groups were formed in the Habranthus dendrogram. Through the component graph, it was
possible to observe the descriptors that contributed the most to the categorization of soil cover rates, leaf length, dry leaf index and leaf width, which was a better feature of this genus. In
Hippeastrum dendrogram were formed seven groups. Through the component graph, it was
possible to observe the descriptors that contributed the most to the categorization of soil cover rates, leaf length, dry leaf index and leaf width, which was a better feature of this genus. It is concluded that the descriptors used contributed to the characterization of the accessions of
Habranthus and Hippeastrum, also determining the genetic divergence between them.
SUMÁRIO
1 INTRODUÇÃO... 11
2 OBJETIVOS... 14
3 REVISÃO BIBLIOGRÁFICA... 15
3.1 A família Amaryllidaceae... 15
3.1.1 Uso ornamental... 15
3.1.2 Uso medicinal... 16
3.2 O gênero Habranthus Herb. ... 17
3.3 O gênero Hippeastrum Herb... 18
3.4 Diversidade genética e caracterização de germoplasma... 18
3.5 Recursos genéticos de espécies de Amaryllidaceae... 20
4 MATERIAL E MÉTODOS... 21
5 RESULTADOS E DISCUSSÃO ... 24
5.1 Caracterização do Gênero Habranthus... 24
5.2 Caracterização do Gênero Hippeastrum... 31
6 CONCLUSÕES ... 37
REFERÊNCIAS ... 38
APÊNDICE – Acessos da família Amaryllidaceae usados para caracterização morfoagronômica realizada neste estudo... 44 ANEXO – Procedência dos acessos dos gêneros Habranthus e
1 INTRODUÇÃO
A floricultura brasileira vem adquirindo nos últimos anos um bom desenvolvimento e índices de crescimento relevantes, representados por uma cadeia produtiva crescente em função de um mercado promissor tanto para produtores, quanto para atacadistas, varejistas e outros profissionais relacionados (JUNQUEIRA; PEETZ, 2017).
Somente em 2017, o setor obteve faturamento de R$ 7,2 bilhões, representando um crescimento de 9% em relação ao ano anterior. Responsável pela geração de quase 200 mil empregos diretos, o mercado de flores e plantas ornamentais é uma das apostas dos empreendedores brasileiros (IBRAFLOR, 2017).
Entre plantas vivas e produtos da floricultura, os itens de maior significância comercial são flores e folhagens de corte, plantas de vaso, plantas perenes e bulbos. O nicho dos bulbos, por exemplo, move na Europa algo em torno de 450 milhões de euros, o segundo nicho com maior movimento (EUROPEAN COMMISSION, 2017) e no Brasil este é o segmento onde está concentrado o maior movimento nas exportações (NEVES; PINTO, 2015). Espécies do grupo como alstroeméria (Alstroemeria sp.), amarílis (Hyppeastrum sp.),
copo-de-leite (Zantedeschia sp.), gladíolo (Gladiolus X grandiflorus), hemerocale
(Hemerocallis sp.) e lírio (Lilium sp.) estão entre as principais bulbosas cultivadas e
comercializadas (TOMBOLATO, 2010).
A cadeia de ornamentais é receptiva a inovações de produtos para atender à demanda do mercado. O interesse por novos produtos permite que um grande número de espécies nativas com reconhecido potencial ornamental, possam ser exploradas comercialmente. Atualmente a utilização dessas espécies é frequentemente restrita regionalmente, além de serem extraídas de natureza de forma indiscriminada (MORAIS et al.,
2017).
As bulbosas ornamentais de maior importância ornamental são as espécies da família Amaryllidaceae, cultivadas e comercializadas com diversas finalidades: flor de corte, planta para vasos e paisagismo. Por possuir elevado valor ornamental e grande apelo comercial, a conservação de sua diversidade natural é uma prioridade (TOMBOLATO et al.,
2010).
O extrativismo mineral, destruição do habitat natural, plantio extensivo e outras atividades antropicas têm eliminado diversas populações de espécies brasileiras de Amaryllidaceae (DUTILH et al., 2013), sendo muitos deles de reconhecido valor ornamental.
O gênero Habranthus Herb. apresenta cerca de 40 espécies distribuídas desde o
sudeste dos Estados Unidos, México até a América do Sul, as plantas deste gênero são reconhecidas por apresentarem relevante riqueza florística com alto apelo ornamental (AMARAL-LOPES; CAVALCANTI, 2015). O gênero Hippeastrum Herb. também é bastante
apreciado pelo mercado de ornamentais, possui aproximadamente 75 espécies, sendo muitas endêmicas do Brasil (DUTILH, 1996). A diversidade de espécies que compõem este gênero e sua variação morfológica tornou possível a criação de centenas de variedades nos últimos dois séculos (TOMBOLATO et al., 2010).
De acordo com Cavalcante et al. (2017), existe uma tendência atual de redução ou
substituição de plantas ornamentais exóticas por espécies nativas. Considerado um mercado competitivo e sempre atento as novidades, o setor do agronegócio ornamental está tendendo a adotar produtos de impacto ambiental reduzido e que promovam conservação de espécies vegetais.
A conservação de recursos genéticos tem a finalidade de conservar a diversidade e a variabilidade das informações genéticas contida nos genomas dos indivíduos representativos das espécies (COSTA; SPEHAR, 2012). Os Bancos Ativos de Germoplasma (BAGs) são uma opção para a conservação de recursos genéticos vegetais (COSTA et al.,
2011). A caracterização de bancos e coleções de germoplasma constitui uma ferramenta importante para a conservação e o melhoramento da espécie alvo e para uso eficiente do germoplasma (FIGAS et al., 2018).
A caracterização consiste na obtenção planejada e sistemática de dados baseada na avaliação de características capazes de descrever e diferenciar os acessos existentes no banco de germoplasma (MONTEIRO, 2015). A caracterização é baseada em observações de variáveis qualitativas e em medidas de variáveis quantitativas de diversos caracteres morfológicos facilmente distinguíveis (BURLE; OLIVEIRA, 2010).
A caracterização de germoplasma é o importante elo entre a conservação e utilização dos recursos genéticos vegetais de forma eficiente, para que variabilidade genética existente na região seja adequadamente estudada, preservada e utilizada pela sociedade (RODRIGUEZ et al. 2005).
2 OBJETIVOS
2.1 Geral
Caracterizar acessos de dois gêneros da coleção de germoplasma da Embrapa Agroindústria Tropical.
2.2 Específicos
Caracterizar morfologicamente e estimar a diversidade genética entre acessos de
Habranthus Herb.
Caracterizar morfologicamente e estimar a diversidade genética entre acessos de
3 REVISÃO BIBLIOGRÁFICA
3.1 A família Amaryllidaceae
Representadas como ervas bulbosas e menos frequentemente como ervas rizomatozas, as espécies da família Amaryllidaceae podem ser encontradas em todo mundo, sendo as regiões tropicais e subtropicais os locais de maior incidência (AMARAL, 2012). Diversos gêneros são importantes como alimentos, como Allium, que abrange centenas de
espécies oriundas da Europa, América do Norte e África. Vegetais de difícil classificação taxonômica, inclui plantas populares como alho, cebolinha e alho-poró (DUTILH, 2005).
Abrangendo as subfamílias Amaryllidoideae, Allioideae e Agapanthoideae, cerca de 73 gêneros e 1605 espécies estão incluídos nesta família (APG III, 2009). Quatro tribos, 12 gêneros e cerca de 100 espécies, representam a família no Brasil, incluindo táxons exclusivamente brasileiros, como Cearanthes Ravenna, Eithea Ravenna, Griffinia Ker Gawl,
Tocantinia Ravenna e Worsleya (W. Watson ex Traub) (AMARAL, 2012).
As Amaryllidaceae são plantas bulbosas e perenes de hábito herbáceo, com folhas com filotaxia dística ou em espiral, inflorescência escaposa, umbeliforme ou uniflora, envolta por duas ou três brácteas maiores e geralmente com mais brácteas internas pequenas, pedicelos não articulados, estilete longo e estigma seco ou úmido, todas as raízes são adventícias e tem sua origem, e modo geral, na placa basal do bulbo (MEEROW; SNIJMAN, 1998). Os bulbos destas plantas possuem grande capacidade de armazenamento de nutrientes e água que são utilizados em épocas secas, momento em que a planta fica sem a parte aérea (ALVES-ARAÚJO; PESSOA; ALVES, 2012).
Possuindo poucas e/ou pequenas populações, determinadas espécies tem distribuição geográfica limitada. Podem ser encontradas em diversos ambientes como matas, campos, montanhas, restingas brejos e sobre pedras geralmente associadas à vegetação herbácea, em matas densas existem a incidência de plantas epífitas. Inúmeras espécies são exploradas de modo predatório, por possuírem atrativo ornamental (DUTILH, J.H.A. et al.,
2013).
3.1.1 Uso ornamental
floricultura mundial, explorados principalmente como flor de corte ou vaso, seguida pelos gêneros Habranthus e Narcissus, ambos com reconhecido valor ornamental(FERNÁNDEZ et
al., 2013)
No Brasil, a participação das Amaryllidaceae no comércio exterior, no período de janeiro a agosto de 2014 foi de 61,7%. São Paulo e Ceará são os maiores produtores, sendo que a produção é quase totalmente voltada para exportação (JUNQUEIRA; PEETZ, 2015).
O cultivo das Amaryllidaceae, com destaque para os gêneros Habranthus e
Hippeastrum, assim como outras ornamentais nativas, pode ser aprimorado através do
desenvolvimento de cultivares nacionais, ações de pré-melhoramento e melhoramento, focados principalmente em qualidades ornamentais e resistência contra pragas e doenças. Seleção de genótipos superiores e indicação de cultivares podem contribuir para aumentar a competitividade entre produtores, gerando divisas, independência tecnológica e preservação da flora nacional (BOTELHO; RODRIGUES; BRUZI, 2015).
As Amaryllidaceae também são muito bem adaptadas ao cultivo no Brasil em suas diferentes regiões, além disso, é grande número de espécies nativas e não exploradas. A maioria das espécies de Hippeastrum é endêmica da grande Bacia Amazônica, considerada o
centro de dispersão deste gênero (MATEUS et al., 2010; NASCIMENTO et al., 2013 ). “Hippeastrum x hybridum” é a cultivar mais comercializada no Brasil, sendo que seu cultivo é
direcionado para a produção de bulbos, visando à exportação para países como a Holanda, México, Chile e Argentina (AMARAL, 2012).
Segundo Amaral-Lopes & Cavalcanti (2012), estudar e conhecer as espécies pertencentes a esta família é importante não somente pelo conhecimento da diversidade, mas também pelo potencial ornamental na flora tropical que seus membros como espécie têm, dado ao fato de que o setor produtor de flores e plantas ornamentais no Brasil é quase exclusivamente composto por espécies de plantas introduzidas.
3.1.2 Uso medicinal
A família Amaryllidaceae é conhecida pela sua utilidade ornamental e medicinal. Os atributos medicinais geram medicamentos importantes, como o indicado para Alzheimer, galantamina, um inibidor potente e seletivo da enzima acetilcolinesterase, de significância na progressão da neurodegeneração associada a doenças neuromotoras (NAIR; STADEN, 2013).
potencial de interesse farmacológico (KILGORE; KUTCHAN, 2016).
Os alcaloides extraídos dessas plantas têm sido intensamente estudados desde 1877, quando a licorina foi isolada. Essa substância foi estudada pelas suas propriedades anticancerígenas (ANDRADE et al., 2003). Outros grupos de alcaloides encontrados em
Amaryllidaceae como lincanina, crinina, tazetina e galanthamina apresentam propriedades
antimaláricas (ŞENER; ORHAN; SATAYAVIVAD, 2003).
3.2 O Gênero Habranthus Herb.
O gênero Habranthus foi identificado em 1824 pelo estudioso de plantas
bulbosas, William Herbert. Suas espécies são nativas das Américas (GOVAERTS, 2015), tendo proximadamente 40 espécies identificadas na região que compreende desde o sudoeste dos Estados Unidos até o sul da América do Sul (OLIVEIRA; ANTOINETTE; SANO, 2010).
Alguns exemplares do gênero também podem ser encontrados em outras regiões como Índia e África do Sul (GOVAERTS, 2015).
O Brasil possui cerca de 23 espécies nativas (DUTILH; OLIVEIRA, 2015). A maior parte está distribuída nos dominios fitogeógraficos da Caatinga, Cerrado, Mata Atlântica, Pantanal e Pampas, sendo incidentes em vegetação do tipo caatinga arbustivo-arbórea, campo rupestre, carrasco, mata de galeria, campo limpo, cerrado sensu stricto,
cerrado de altitude e também em locais onde ocorrem alagamentos. Duas explicações são utilizadas para justificar a falta de registro de coleta na região amazônica, uma é a sazonalidade pouco evidente, característca ambiental importante para as plantas do gênero, ou pouco empenho na coleta (AMARAL, 2012).
As plantas do gênero possuem folhas anuais, lineares ou filiformes. Escapo oco, cilíndrico; inflorescência reduzida a uma única flor; brácteas fundidas na metade inferior formando um tubo. Flores declinadas, infundibuliformes ou crateriformes, zigomorfas. Corona de escamas pequenas ou de fímbrias às vezes presente; tépalas elípticas ou ovais. Estames declinado-ascendentes, filetes filiformes, desiguais. Anteras oblongas. Estigma trífido ou trilobado, estilete filiforme. Sementes achatadas (OLIVEIRA; ANTOINETTE; SANO, 2010).
Fazer a classificação das espécies de Habranthus nativas do Brasil geralmente é
difícil. Existe apenas uma obra literária que aborda as espécies de forma geral, o trabalho
considerada inconclusiva pois não existem chaves de identificação para as espécies nem pranchas explicativas, a exemplo do que ocorre para outras famílias (AMARAL, 2012).
A separação morfológica entre Habranthus e Zephyranthes não é bem definida.
Ela se baseia na simetria floral, sendo o gênero Zephyrantes caracterizados por flores mais
actinomorfas, eretas, com estames eretos de mesmo comprimento. Enquanto Habranthus tem
flores mais zigomorfas declinadas, com estames declinados e de comprimentos diferentes (DUTILH, 2005).
3.3 O Gênero Hippeastrum Herb.
O gênero tem cerca de 54 espécies nativas da América do Sul. No Brasil existem aproximadamente 30 espécies distribuídas nos domínios fitogeográficos da Amazônia, Caatinga, Cerrado, Mata Atlântica, Pampa e Pantanal. São plantas que ocorrem em vegetações de todos os tipos, como caatinga, campinarana, campos, carrasco, cerrado, florestas, restinga, savana amazônica, vegetação aquática e vegetação sobre afloramentos rochosos. Podem ter hábito terrestre, saxícola, epifítico ou aquático (DUTILH; OLIVEIRA, 2015).
As plantas do gênero Hippeastrum Herb. são caracterizadas como ervas com
folhas anuais, raramente persistentes, sésseis, raramente sub-pecioladas, geralmente com mais de 2 cm largura. Inflorescência com escapo oco, cilíndrico a levemente comprimido; brácteas espatáceas, livres. Flores declinadas, zigomorfas; corona de margem calosa ou fimbriada na fauce; estames declinado-ascendentes, de comprimentos diferentes; estigma trífido ou trilobado. Sementes achatadas enegrecidas. O número cromossômico básico para o gênero é x=11 (FLORA DO BRASIL, 2018).
São popularmente conhecidas com “Amarílis” (embora o gênero Amaryllis seja
exclusivamente africano), os híbridos e variedades comerciais deste gênero são de grande importância econômica no mercado mundial de plantas ornamentais. A maioria das espécies apresenta reprodução vegetativa pela formação de bulbilhos laterais (DUTILH, 2005).
3.4 Diversidade genética e caracterização de germoplasma
espécies e os ecossistemas (MAGNUSSON et al., 2016)
Todo tipo de material de origem biológica (espécies de plantas, animais e microrganismos integrantes da biodiversidade) que tem valor socioeconômico atual e potencial para utilização em programas de melhoramento genético, biotecnologia e outras ciências semelhantes é considerado recurso genético (QUEIRÓZ, 1999).
Os recursos genéticos desempenham um papel fundamental no melhoramento de plantas porque disponibilizam a variabilidade genética para produzir novas combinações gênicas que através da seleção permitem desenvolver novas cultivares de interesse (KARASAWA, 2016).
A conservação dessa variabilidade das informações genéticas contida nos indivíduos representativos das espécies é muito importante e as principais formas de
conservação de recursos genéticos são “in situ”- quando a preservação das espécies é dentro do ambiente natural de ocorrência a que estão adaptadas –e “ex situ” – quando a conservação das espécies é fora do seu local de ocorrência (COSTA; SPEHAR, 2012).
A conservação ex situ tem enorme relevância não apenas como base para o melhoramento, que busca alto desempenho e maior produtividade é também uma garantia da
contínua utilização das espécies vegetais. O modelo da conservação “ex situ”, sob a forma de
bancos de germoplasma, historicamente foi a forma prioritária de combate ao processo de erosão genética e constitui-se um poderoso aliado para sobrevivência desses (SANTONIERI; BUSTAMANTE, 2016) .
Os Bancos Ativos de Germoplasma (BAGs) quando bem formados, se tornam ferramentas essenciais para a conservação do máximo possível da variabilidade genética existente na natureza. Os acessos conservados devem ser regenerados, caracterizados, documentados e disponibilizados para o uso e intercâmbio (VEIGA; BARBOSA, 2012).
A caracterização é indispensável para a manutenção de coleções de germoplasma, que consiste em descrever, identificar e diferenciar acessos de uma coleção. A primeira caracterização realizada no germoplasma, depois que ele é adicionado ao banco é a caracterização morfológica, ela é baseada em observações de variáveis qualitativas e em mensurações de variáveis quantitativos de diversos caracteres morfológicos facilmente distinguíveis a olho nu (BURLE; OLIVEIRA, 2010).
3.5 Recursos genéticos de espécies de Amaryllidaceae
Apesar das Amaryllidaceae serem nativas das Américas do Sul e Central e haverem cerca de 40 espécies nativas de Amarílis no Brasil, há carência de acessos destas plantas em bancos de germoplasma no país (AMARAL et al., 2007).
As coleções de germoplasma de espécies de Amaryllidaceae com maior destaque são a coleção do Jardim Botânico do Instituto Agronômico – JBIAC – é provavelmente uma das mais completas do país e ali há também preocupação com a perpetuação das espécies com potencial ornamental, como é o caso das Amaryllidaceae (TOMBOLATO et al., 2009) e a
coleção da Embrapa Recursos Genéticos e Biotecnologia, que conserva mais de 100 acessos, coletados em várias regiões do Brasil. O enriquecimento por meio de coletas e a documentação desta coleção são realizados periodicamente, mas seu acervo não vem sendo caracterizado.
Existem relatos de algum germoplasma desta família conservado em jardins botânicos, a exemplo do Jardim Botânico da Fundação Zoobotânica de Belo Horizonte e o Instituto de Pesquisas Jardim Botânico – RJ (VEIGA et al., 2009).
4 MATERIAL E MÉTODOS
Foram utilizados neste estudo 29 acessos do gênero Habranthus e 23 acessos do
gênero Hippeastrum pertencentes ao Banco Ativo da Embrapa Agroindústria Tropical,
localizado em Fortaleza – CE sob as coordenadas geográficas 3°45'05"S e 38°34'36"W (Tabela 1). Esses acessos são provenientes de coletas realizadas em diversas localidades do Brasil.
As avaliações foram realizadas no período de janeiro a abril de 2018. As plantas tinham 24 meses e se encontravam no estádio vegetativo do desenvolvimento. Foram cultivadas em vasos de 5 litros com substrato comercial e matéria orgânica (3:1), irrigadas por aspersão e mantidas em telado com sombreamento 50%. Foram avaliados 5 vasos por acesso e o delineamento utilizado foi o inteiramente casualizado.
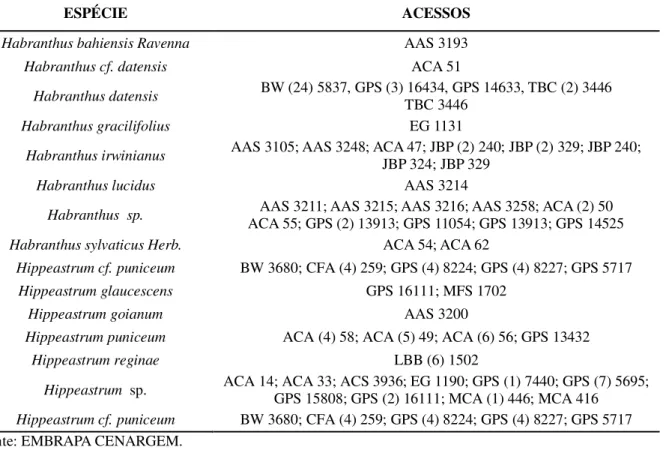
Tabela 1 – Dados referentes aos acessos de Habranthus e Hippeastrum pertencentes à coleção da Embrapa
Agroindústria Tropical Fortaleza, 2018.
ESPÉCIE ACESSOS
Habranthus bahiensis Ravenna AAS 3193
Habranthus cf. datensis ACA 51
Habranthus datensis BW (24) 5837, GPS (3) 16434, GPS 14633, TBC (2) 3446 TBC 3446
Habranthus gracilifolius EG 1131
Habranthus irwinianus AAS 3105; AAS 3248; ACA 47; JBP (2) 240; JBP (2) 329; JBP 240;
JBP 324; JBP 329
Habranthus lucidus AAS 3214
Habranthus sp. ACA 55; GPS (2) 13913; GPS 11054; GPS 13913; GPS 14525 AAS 3211; AAS 3215; AAS 3216; AAS 3258; ACA (2) 50
Habranthus sylvaticus Herb. ACA 54; ACA 62
Hippeastrum cf. puniceum BW 3680; CFA (4) 259; GPS (4) 8224; GPS (4) 8227; GPS 5717
Hippeastrum glaucescens GPS 16111; MFS 1702
Hippeastrum goianum AAS 3200
Hippeastrum puniceum ACA (4) 58; ACA (5) 49; ACA (6) 56; GPS 13432
Hippeastrum reginae LBB (6) 1502
Hippeastrum sp. ACA 14; ACA 33; ACS 3936; EG 1190; GPS (1) 7440; GPS (7) 5695; GPS 15808; GPS (2) 16111; MCA (1) 446; MCA 416 Hippeastrum cf. puniceum BW 3680; CFA (4) 259; GPS (4) 8224; GPS (4) 8227; GPS 5717
Fonte: EMBRAPA CENARGEM.
estádio de desenvolvimento em que as plantas se encontravam (vegetativo) e nos descritores adaptados de Amarílis (Hippeastrum Herb.) da UPOV – União Internacional para Proteção de
Novas Variedades de Plantas na língua Portuguesa para uso no SNPC – Sistema Nacional para Proteção de Cultivares, do Ministério da Agricultura, Pecuária e Abastecimento do Brasil (TOMBOLATO et al., 2007).
O CFO foi medido da extremidade da folha até o início do pedicelo, as medições foram feitas com régua e os valores obtidos em centímetros (cm). A LFO foi medida na parte mais larga do limbo das folhas, as medições foram feitas com paquímetro manual obtendo valores em centímetros (adaptada de Alves-Araújo; Alves, 2007). Todos os descritores relacionados às folhas foram avaliados em todas as folhas totalmente expandidas em cada repetição e foram obtidas médias dos valores de cada descritor para caracterização dos acessos.
A TRS (adaptada de Castro et al., 2015), a AIN (adaptada Eumura-Lima et al., 2017),
a OPD (adaptada de Castro et al., 2011) e a FSE (adaptada de Castro et al., 2011) foram
avaliadas por meio de escala arbitrária de notas (Tabela 2). A taxa de recobrimento do solo foi obtida estimando-se a porcentagem de área de substrato coberto, sob o ponto de vista do observador a aproximadamente 1 metro de distância do vaso.
O índice de ataques de insetos foi obtido pela contagem do número de folhas com ataques evidentes de insetos (lagartas, gafanhotos, etc.), foram consideradas injúrias, recortes, raspagens e folhas totalmente consumidas. A ocorrência de plantas daninhas foi avaliada através da quantificação de plantas daninhas presentes em cada vaso. O índice de folhas secas foi obtido pela porcentagem de folhas secas e/ ou amareladas em relação ao total de folhas em cada vaso.
Inicialmente, os dados de CFO e LFO foram submetidos à análise de variância para averiguação de variabilidade genética entre os acessos e, em seguida, os valores médios foram ordenados segundo o teste de agrupamento Scott-Knott (SCOTT; KNOTT, 1974), a 5% de probabilidade. Para análise da variabilidade e da estimativa da divergência genética entre os acessos, foram obtidas as distâncias euclidianas. Com as estimativas de similaridade genética, foi construído o dendrograma, pelo método UPGMA.
Tabela 2 – Sistema de notas utilizado para avaliar acessos de Amaryllidaceae pertencentes à coleção da Embrapa Agroindústria Tropical. Fortaleza, 2018.
Fonte: adaptada de Castro et al. (2015), Eumura-Lima et al. (2017), Castro et al. (2011).
Nota Taxa de recobrimento do solo (TRS) Ocorrência de plantas daninhas (OPD) Ataque de insetos (AIN) Índice de folhas secas (FSE)
5 Substrato totalmente coberto ou com poucas
falhas, cobertura de mais de 80%;
Sem ocorrência; Nenhuma folha com ataques aparentes;
Nenhuma ou poucas folhas secas, até 15%
por vaso;
4 Poucas falhas, cobertura de 61% até 80% do
substrato;
Baixa ocorrência, até
5 unidades por vaso; ataques, até 5 folhas Nível baixo de por vaso;
Baixa porcentagem de folhas secas, de 16% a
30% por vaso;
3 Moderado número de falhas, cobertura de
41% até 60% do substrato.
Ocorrência moderada, de 6 a 10 unidades por
vaso;
Nível moderado de ataques, de 6 a 10
folhas por vaso;
Moderada porcentagem de folhas
secas, de 31% a 45%;
2 Muitas falhas, cobertura de 21% a 40% do
substrato;
Alta ocorrência, de 11 a 15 unidades por
vaso;
Nível alto de ataques, de 11 a 15 folhas por
vaso;
Alta porcentagem de folhas secas, de 46% a
60% por vaso;
1 Alto número de falhas ou substrato totalmente
a mostra, cobertura de até 20%.
Altíssima ocorrência, mais de 15 unidades
por vaso.
Nível altíssimo de ataques, mais 15
folhas atacadas.
Mais de 60% de folhas secas e/ou amareladas
5 RESULTADOS E DISCUSSÃO
5.1 Caracterização e divergência genética de Habranthus
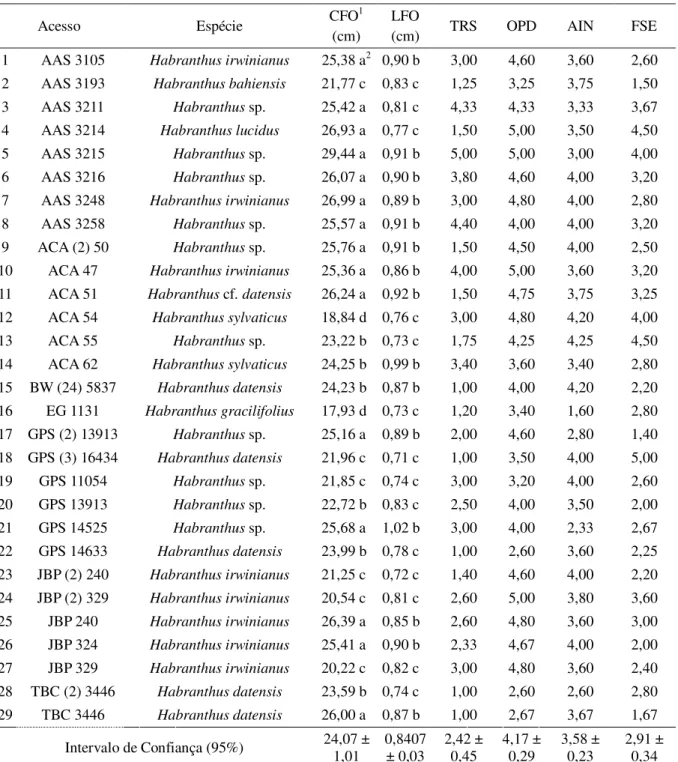
Houve variação para todos os caracteres avaliados dentro dos acessos do gênero
Habranthus (Tabela 3). O comprimento das folhas (CFO) variou de 17,93 a 29,44 cm. Os
menores comprimentos de folhas foram dos acessos EG 1131 (17,93 cm), ACA 54 (18,84 cm) e JBP (2) 329 (20,22 cm). Já os que apresentaram os maiores comprimentos foram os acessos AAS 3214 (26,93 cm), AAS 3215 (29,44 cm), AAS 3248 (26,99 cm) e JBP 240 (26,39 cm). A largura de folhas (LFO) variou de 0,71 cm a 1,02 cm. As menores larguras de folhas foram dos acessos ACA 55 (0,73 cm), EG 1131 (0,73 cm), GPS (3) 16434 (0,71 cm) e JBP (2) 240 (0,72 cm). Os que apresentaram maior largura foram ACA 51 (0,92 cm), ACA 62 (0,99 cm) e GPS 14525 (1,02 cm).
No descritor taxa de recobrimento do solo (TRS) houve variação entre os acessos, apresentando valores médios variando de 1,00 a 5,00. Todos os acessos pertencentes à espécie
Habranthus datensis deste grupo apresentaram nota mínima para esse caractere. A ocorrência
de plantas daninhas (OPD) variou entre 2,60 e 5,00, os acessos que apresentaram as menores notas nesse critério (inferior a três) foram novamente os acessos pertencentes à espécie
Habranthus datensis. O ataque de insetos (AIN) apresentou variação de 1,60 a 4,20. O único
acesso da espécie Habranthus gracilifolius apresentou a menor nota neste critério,
demonstrando ser o mais passível de ataques de insetos. O índice de folhas secas (FSE) variou de 1,40 a 5,00.
Utilizando-se do método de agrupamento UPGMA foi possível observar a formação de seis grupos entre os acessos do gênero Habranthus (Figura 1). O grupo 1 reuniu
o maior número de acessos, foram eles: AAS 3105, AAS 3216, AAS 3248, AAS 3258, ACA (2) 50, ACA 47, ACA 51, AAS 3211, AAS 3214, ACA 62, GPS (2) 13913, GPS 14525, JBP 240 e JBP 324. Os acessos deste grupo apresentaram CFO (24,25 a 26, 99 cm) e LFO (0,77 a 1,02 cm) bastante similares, outra característica comum é a baixa ocorrência de plantas daninhas. Se comparadas as médias de AIN dos outros grupos, o grupo 1 apresenta a terceira maior nota, o que indica um número médio de folhas atacadas (Tabela 4).
Quatro dos cinco acessos da espécie Habranthus datensis (BW (24) 5837, GPS
14633, TBC (2) 3446 e TBC 3446 formaram o grupo 2. Além da semelhança fenotípica
demonstraram os valores mais baixos para os caracteres TRS e FSE dentre todos os grupos (Tabela 4).
Tabela 3 – Médias e valores referentes aos descritores morfológicos avaliados em 29 acessos de Habranthus,
Fortaleza-CE, 2018.
Fonte: própria autoria.
1/comprimento de folhas – CFO; largura de folhas – LFO; taxa de recobrimento do solo – TRS; ocorrência de
plantas daninhas – OPD; ataque de insetos – AIN; folhas secas – FSE. 2/ Médias seguidas da mesma letra na
coluna não diferem entre si, pelo teste Scott-Knott, a 5% de significância.
Acesso Espécie CFO
1
(cm)
LFO
(cm) TRS OPD AIN FSE 1 AAS 3105 Habranthus irwinianus 25,38 a2 0,90 b 3,00 4,60 3,60 2,60
2 AAS 3193 Habranthus bahiensis 21,77 c 0,83 c 1,25 3,25 3,75 1,50
3 AAS 3211 Habranthus sp. 25,42 a 0,81 c 4,33 4,33 3,33 3,67
4 AAS 3214 Habranthus lucidus 26,93 a 0,77 c 1,50 5,00 3,50 4,50
5 AAS 3215 Habranthus sp. 29,44 a 0,91 b 5,00 5,00 3,00 4,00
6 AAS 3216 Habranthus sp. 26,07 a 0,90 b 3,80 4,60 4,00 3,20
7 AAS 3248 Habranthus irwinianus 26,99 a 0,89 b 3,00 4,80 4,00 2,80
8 AAS 3258 Habranthus sp. 25,57 a 0,91 b 4,40 4,00 4,00 3,20
9 ACA (2) 50 Habranthus sp. 25,76 a 0,91 b 1,50 4,50 4,00 2,50
10 ACA 47 Habranthus irwinianus 25,36 a 0,86 b 4,00 5,00 3,60 3,20
11 ACA 51 Habranthus cf. datensis 26,24 a 0,92 b 1,50 4,75 3,75 3,25
12 ACA 54 Habranthussylvaticus 18,84 d 0,76 c 3,00 4,80 4,20 4,00
13 ACA 55 Habranthus sp. 23,22 b 0,73 c 1,75 4,25 4,25 4,50
14 ACA 62 Habranthus sylvaticus 24,25 b 0,99 b 3,40 3,60 3,40 2,80
15 BW (24) 5837 Habranthus datensis 24,23 b 0,87 b 1,00 4,00 4,20 2,20
16 EG 1131 Habranthus gracilifolius 17,93 d 0,73 c 1,20 3,40 1,60 2,80
17 GPS (2) 13913 Habranthus sp. 25,16 a 0,89 b 2,00 4,60 2,80 1,40
18 GPS (3) 16434 Habranthus datensis 21,96 c 0,71 c 1,00 3,50 4,00 5,00
19 GPS 11054 Habranthus sp. 21,85 c 0,74 c 3,00 3,20 4,00 2,60
20 GPS 13913 Habranthus sp. 22,72 b 0,83 c 2,50 4,00 3,50 2,00
21 GPS 14525 Habranthus sp. 25,68 a 1,02 b 3,00 4,00 2,33 2,67
22 GPS 14633 Habranthus datensis 23,99 b 0,78 c 1,00 2,60 3,60 2,25
23 JBP (2) 240 Habranthus irwinianus 21,25 c 0,72 c 1,40 4,60 4,00 2,20
24 JBP (2) 329 Habranthus irwinianus 20,54 c 0,81 c 2,60 5,00 3,80 3,60
25 JBP 240 Habranthus irwinianus 26,39 a 0,85 b 2,60 4,80 3,60 3,00
26 JBP 324 Habranthus irwinianus 25,41 a 0,90 b 2,33 4,67 4,00 2,00
27 JBP 329 Habranthus irwinianus 20,22 c 0,82 c 3,00 4,80 3,60 2,40
28 TBC (2) 3446 Habranthus datensis 23,59 b 0,74 c 1,00 2,60 2,60 2,80
29 TBC 3446 Habranthus datensis 26,00 a 0,87 b 1,00 2,67 3,67 1,67
Figura 1 – Dendrograma obtido com base nas medidas de similaridade genética e método de agrupamento UPGMA a partir da distância Euclidiana, considerando os seis descritores morfoagronômicos avaliados nos 29 acessos* de Habranthus. Fortaleza-CE, 2018.
Fonte: própria autoria.
*/Legenda dos acessos consta na Tabela 3.
Tabela 4 - Médias e valores referentes aos descritores morfológicos avaliados em grupos de Habranthus
formados a partir de dendrograma obtido pelo método UPGMA. Fortaleza-CE, 2018.
Grupo CFO(cm) 1 LFO (cm) TRS OPD AIN FSE
1 25,76 0,89 2,88 4,52 3,57 2,91
2 24,45 0,82 1,00 4,00 3,52 2,23
3 21,03 0,79 2,39 4,24 3,84 2,61
4 22,59 0,72 1,38 3,88 4,13 4,75
5 17,93 0,73 1,20 3,40 1,60 2,80
6 29,44 0,91 5,00 5,00 3,00 4,00
Fonte: própria autoria.
1/comprimento de folhas – CFO; largura de folhas – LFO; taxa de recobrimento do solo – TRS; ocorrência de
plantas daninhas – OPD; ataque de insetos – AIN; folhas secas – FSE.
O terceiro grupo foi formado por 7 acessos das espécies Habranthus bahiensis
(AAS 3193), Habranthus sylvaticus (ACA 54), Habranthus sp. (GPS 11054 e GPS 13913) e
Habranthus irwinianus (JBP (2) 240, JBP (2) 329, JBP 329). As plantas deste grupo
apresentaram semelhanças nos caracteres OPD (3,20 a 5,00) e AIN (3,50 a 4,20).
Apenas os acessos ACA 55, GPS (3) 16434 compuseram o grupo 4. As plantas deste grupo demonstraram semelhança nas seguintes características: menor valor médio de largura de folhas, baixa TRS (1,00 e 1,75), baixo número de folhas atacadas por insetos e menor número de folhas secas por vaso (Tabela 4). O grupo 5 foi formado pelo o único acesso pertencente a espécie Habranthus gracilifolius (EG 1131), o qual se diferenciou por
apresentar folhas menos compridas dentre todos os acessos, baixos valores para TRS, AIN e FSE, contudo apresentou valor médio para OPD.
O grupo 6 foi formado apenas pelo acesso AAS 3215, da espécie Habranthus sp..
Fenotipicamente se mostrou diferente dos demais por possuir folhas de maior comprimento e largura. Além disso, exibiu valor máximo para os caracteres TRS e OPD (5,00). Para AIN obteve o valor 3,00 e para FSE valor 4,00.
O método de análise por componentes principais (PCA) proporcionou um estudo multivariado dos dados experimentais obtidos, facilitando a visualização da relação entre os acessos e caracteres (Figura 2). Os pesos nas duas primeiras componentes principais representaram aproximadamente 76,20% das informações contidas nos valores médios dos seis caracteres avaliados. A técnica dos componentes principais possibilita a avaliação da importância de cada caráter estudado sobre a variação total disponível entre os genótipos avaliados e também gera a possibilidade de descarte de caracteres que contribuem pouco para a discrição do acesso avaliado (CRUZ; FERREIRA; PESSONI, 2011).
Um vetor muito próximo de um dos eixos terá maior influência sobre esse eixo que sobre outro mais afastado. Então, diz-se que aquele eixo explica a maior parte da informação contida no vetor (GRANATE et. al., 2001). Pode-se notar que as variáveis que tiveram maior influência no 1º componente (eixo horizontal) foram TRS e CFO e as que tiveram maior influência no 2º componente (eixo vertical) foram FSE e LFO. Esse fato também pode ser ressaltado quando se observa o peso de cada um dos seis caracteres nos componentes principais (Tabela 5).
demonstrando menor variabilidade entre os acessos nessas variáveis.
Figura 2 – Gráfico dos componentes principais englobando os acessos de Habranthus e o biplot das
características avaliadas. Fortaleza-CE, 2018.
Fonte: própria autoria.
Comprimento de folhas – CFO; largura de folhas – LFO; taxa de recobrimento do solo – TRS; ocorrência de plantas daninhas – OPD; ataque de insetos – AIN; folhas secas – FSE.
Tabela 5 - Peso de cada um dos 6 descritores no primeiro e segundo componente principal para Habranthus
Fortaleza-CE, 2018.
Variável Componente Principal
1º 2º 3º 4º 5º 6º
TRS1 0.519 0.114 -0.323 0.320 -0.664 0.265
OPD 0.460 0.355 -0.004 0.434 0.678 0.117
AIN 0.106 0.403 0.860 0.030 -0.283 -0.079
FSE 0.125 0.652 -0.353 -0.505 -0.032 -0.423
CFO 0.499 -0.229 0.155 -0.670 0.133 0.456
LFO 0.494 -0.470 0.091 0.057 0.000 -0.723
Fonte: própria autoria.
1/comprimento de folhas – CFO; largura de folhas – LFO; taxa de recobrimento do solo – TRS; ocorrência de
plantas daninhas – OPD; ataque de insetos – AIN; folhas secas – FSE.
A posição dos vetores demonstra o nível de influência de um sobre o outro. Quando os vetores têm a mesma direção representam correlações positivas (formam ângulo menor que 90º), quando os vetores têm direções contrárias representam correlações negativas (formam ângulo de 180°), quando os vetores se sobrepõem representam a correlação máxima e quando os vetores são perpendiculares representam a correlação nula (formam ângulo de 90°) (GRANATE et. al., 2001).
Nenhum vetor das variáveis analisadas se correlacionou negativamente. Os vetores de CFO e LFO se correlacionaram positivamente, demonstrando que acessos que apresentavam maior comprimento de folhas também apresentavam maior largura das mesmas. Tanto o vetor de CFO quanto o vetor e LFO se correlacionaram positivamente com TRS, o que faz sentido, pois plantas com folhas mais compridas e mais largas tendem a recobrir mais uniformemente o substrato do vaso.
A qualidade de plantas envasadas é essencialmente definida pela aparência visual, que depende da forma, tamanho, cor de flores e folhas. Para plantas de folhagem, a forma, tamanho e a maneira como as folhas se dispõem no vaso, são as características mais observadas. Plantas com folhas compridas, largas e com bom recobrimento do substrato contido no vaso são as mais apreciadas (FERRANTE et al., 2015).
O vetor de TRS também se correlacionou positivamente com OPD, demonstrando que plantas com maior recobrimento de solo eram também, as menos infestadas por plantas daninhas. Segundo Bolivar e Alvarenga (2000), plantas que possuem crescimento de folhas mais próximos ao solo/substrato, com capacidade maior de recobrimento, oferecem maior pressão de controle sobre as plantas daninhas, uma vez que apresentam maior capacidade de abafamento e agressividade, diminuindo a sua população, devido à competição por fatores de crescimento, especialmente luz.
Houve correlação nula entre os vetores FSE, LFO e CFO, demonstrando que o índice de folhas secas não se relaciona com o comprimento e a largura de folhas. O vetor de AIN apresentou forte correlação positiva com FSE, ou seja, plantas menos atacadas por insetos eram também, as plantas com menos folhas secas. O ataque de insetos deprecia o aspecto visual e a longevidade das folhas em Amaryllidaceae (TOMBOLATO, 2004).
O método de análise por componentes principais (PCA) também apresentou boa concordância com a análise de agrupamento observados no dendrograma (UPGMA), sendo coerentes os resultados obtidos a partir do coeficiente de similaridade considerando os acessos mais e menos similares com as projeções desses acessos no espaço bidimensional (Figura 2).
quadrantes). Os que estavam localizados no 4º quadrante foram assim agrupados por possuírem comprimento e largura de folhas semelhantes, já os acessos localizados no 1º quadrante ficaram próximos da zona de influência dos vetores TRS e OPD, demonstrando que estes acessos têm boa taxa de recobrimento do solo e baixa ocorrência de plantas daninhas.
Todos os acessos do grupo 2 estão localizados no 3º quadrante do gráfico. Esses acessos apresentaram as menores médias de TRS e FSE entre todos os grupos formados no dendrograma, isso explica o fato de estarem distribuídos no sentido oposto dos vetores de
TRS e FSE. Os sete acessos do grupo 3 foram distrbuídos nos 2º e 3º quadrantes. Dois acessos deste grupo (AAS 3193 e GPS 13913) foram dispostos do 3º quadrante por apresentarem os menores valores de TRS em relação ao demais componentes do grupo. Os outros cinco (ACA 54, GPS 11054, JBP (2) 240, JBP (2) 329 E JBP 329) ficaram no 2º quadrante e apresentaram valores similares para todas as variáveis estudadas.
Os acessos do grupo 4 foram dispostos na área superior do 2º quadrante, no sentido contrário dos vetores TRS e LFO, ou seja, são acessos que foram agrupados por possuírem folhas estreitas e com baixa taxa de recobrimento do solo. O acesso EG 1131, único componente do grupo 5 foi disposto mais próximo da extremidade esquerda do 3º quadrante, distante dos eixos, se caracterizando por possuir o menor CFO e baixa TRS. O acesso AAS 3215, único representante do grupo 6, foi disposto na extremidade do 1º quadrante, próximo ao eixo do 1º componente, sendo influenciado pelos vetores de TRS e CFO.
De forma geral, os acessos de Habranthus possuem uma alta variabilidade
5.2 Caracterização e divergência genética de Hippeastrum
Os acessos MCA 416, ACA (4) 58 e GPS (1) 7440 não foram considerados nas análises estatisticas e de similaridade, pois não tinham folhas no momento da avaliação,
contudo, os bulbos estavam “vivos”. O que pode explicar o fato é que o gênero Hippeastrum
está incluído no grupo de plantas chamadas geófitas, essas plantas apresentam completa separação entre as fases de produção e armazenamento de assimilados, onde as folhas e flores ocorrem em peródos diferentes e os bulbos passam por período de dormência. Esses bulbos aparentemente dormentes podem estar iniciando o processo de florescimento (TOMBOLATO, 2004).
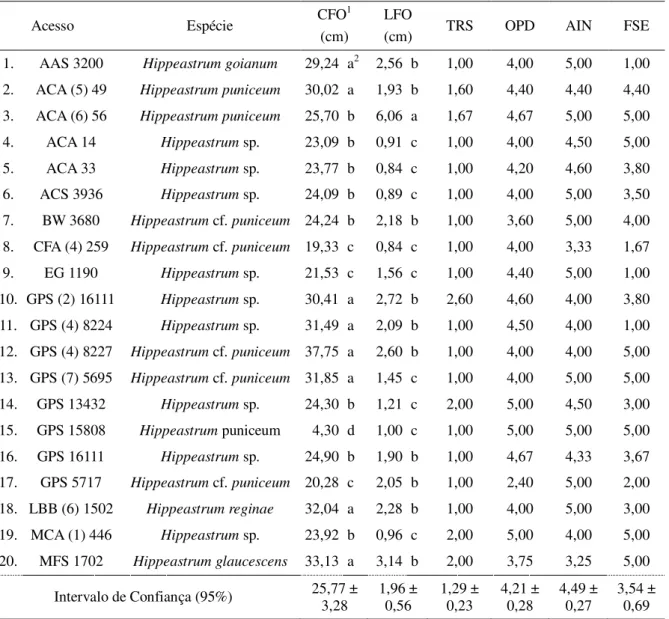
Os demais acessos apresentaram variação para todos os descritores avaliados (Tabela 6). O comprimento das folhas (CFO) variou de 4,30 cm a 37,75 cm apresentando diferença estatística. Os que apresentaram o maior comprimento foram GPS (4) 8227 (37,75 cm), LBB (6) 1502 (32,04 cm) e MFS 1702 (33,13 cm) e os acessos que apresentaram os menores foram CFA (4) 259 (19,33 cm), GPS 15808 (4,30 cm) e GPS 5717 (20,28 cm). A largura de folhas (LFO) variou de 0,84 cm a 6,06 cm, os acessos que apresentaram folhas mais largas foram: ACA (6) 56 (6,06 cm), GPS (2) 16111 (2,72 cm) e MFS 1702 (3,14 cm). Já os que apresentaram folhas mais estreitas foram: ACA 14 (0,91 cm), ACA 33 (0,84 cm) e CFA (4) 259 (0,84 cm).
A taxa de recobrimento do solo (TRS) não apresentou grande variabilidade entre os acessos, variando de 1,00 a 2,60, demonstrando que os acessos deste gênero não tem bom recobrimento do substrato em vasos. A ocorrência de plantas daninhas (OPD) variou de 2,40 a 5,00, contudo apenas três acessos apresentaram valores abaixo de 4,00 (BW 3680, GPS 5717, MFS 1702). O ataque de insetos (AIN) variou de 3,25 a 5,00, sendo que a maioria dos acessos obteve valores de 4,00 a 5,00, demonstrando que as plantas do gênero Hippeastrum são pouco
atacadas por insetos. O índice de folhas secas (FSE) foi o que demonstrou maior variabilidade entre os acessos, tendo valores de 1,00 a 5,00.
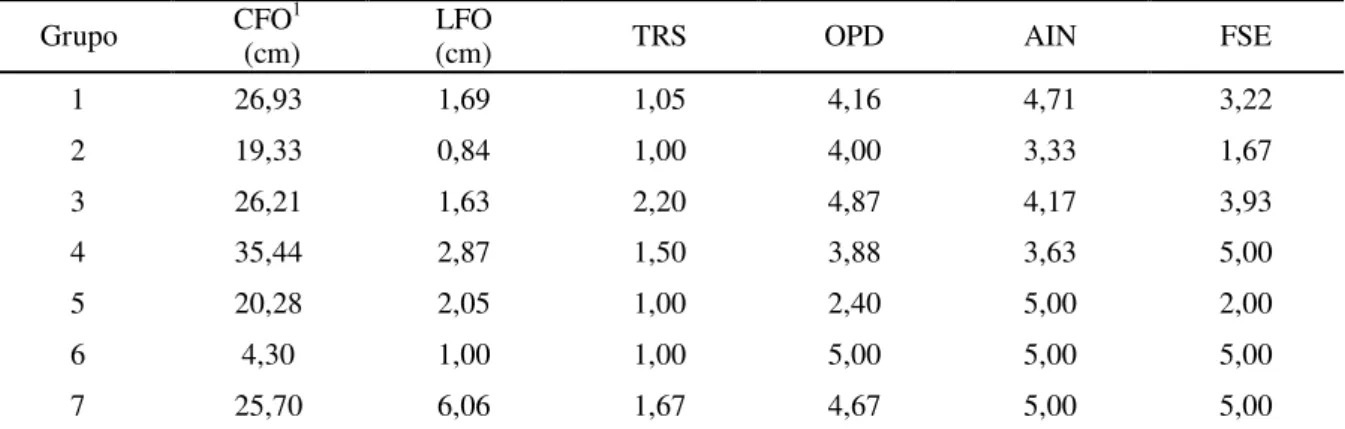
Através do método de agrupamento UPGMA ocorreu a formação de sete grupos entre os acessos do gênero Hippeastrum (Figura 3). O primeiro grupo foi formado por 11
acessos: AAS 3200, ACA (5) 49, ACA 14, ACA 33, ACS 3936, BW 3680, EG 1190, GPS (4) 8224, GPS (7) 5695, GPS 16111 e LBB (6) 1502. Este grupo reuniu acessos com AIN variando de 4,33 a 5,00 e OPD variando de 3,60 a 4,50 evidenciando que são plantas pouco atacadas por insetos e com baixa ocorrência de daninhas.
cf. puniceum. Se comparado aos valores médios dos caracteres de cada grupo, este acesso apresenta o segundo menor comprimento de folhas (19,33 cm) e o menor valor para o indice de folhas secas (1,67) (Tabela 7).
Tabela 6 – Médias e valores referentes aos descritores morfológicos avaliados em 20 acessos de Hippeastrum,
Fortaleza-CE, 2018.
Fonte: própria autoria.
1/comprimento de folhas – CFO; largura de folhas – LFO; taxa de recobrimento do solo – TRS; ocorrência de
plantas daninhas – OPD; ataque de insetos – AIN; folhas secas – FSE. 2/ Médias seguidas da mesma letra na
coluna não diferem entre si, pelo teste Scott-Knott, a 5% de significância.
Acesso Espécie CFO
1
(cm)
LFO
(cm) TRS OPD AIN FSE 1. AAS 3200 Hippeastrum goianum 29,24 a2 2,56 b 1,00 4,00 5,00 1,00
2. ACA (5) 49 Hippeastrum puniceum 30,02 a 1,93 b 1,60 4,40 4,40 4,40
3. ACA (6) 56 Hippeastrum puniceum 25,70 b 6,06 a 1,67 4,67 5,00 5,00
4. ACA 14 Hippeastrum sp. 23,09 b 0,91 c 1,00 4,00 4,50 5,00
5. ACA 33 Hippeastrum sp. 23,77 b 0,84 c 1,00 4,20 4,60 3,80
6. ACS 3936 Hippeastrum sp. 24,09 b 0,89 c 1,00 4,00 5,00 3,50
7. BW 3680 Hippeastrum cf. puniceum 24,24 b 2,18 b 1,00 3,60 5,00 4,00
8. CFA (4) 259 Hippeastrum cf. puniceum 19,33 c 0,84 c 1,00 4,00 3,33 1,67
9. EG 1190 Hippeastrum sp. 21,53 c 1,56 c 1,00 4,40 5,00 1,00
10. GPS (2) 16111 Hippeastrum sp. 30,41 a 2,72 b 2,60 4,60 4,00 3,80
11. GPS (4) 8224 Hippeastrum sp. 31,49 a 2,09 b 1,00 4,50 4,00 1,00
12. GPS (4) 8227 Hippeastrum cf. puniceum 37,75 a 2,60 b 1,00 4,00 4,00 5,00
13. GPS (7) 5695 Hippeastrum cf. puniceum 31,85 a 1,45 c 1,00 4,00 5,00 5,00
14. GPS 13432 Hippeastrum sp. 24,30 b 1,21 c 2,00 5,00 4,50 3,00
15. GPS 15808 Hippeastrum puniceum 4,30 d 1,00 c 1,00 5,00 5,00 5,00
16. GPS 16111 Hippeastrum sp. 24,90 b 1,90 b 1,00 4,67 4,33 3,67
17. GPS 5717 Hippeastrum cf. puniceum 20,28 c 2,05 b 1,00 2,40 5,00 2,00
18. LBB (6) 1502 Hippeastrum reginae 32,04 a 2,28 b 1,00 4,00 5,00 3,00
19. MCA (1) 446 Hippeastrum sp. 23,92 b 0,96 c 2,00 5,00 4,00 5,00
20. MFS 1702 Hippeastrum glaucescens 33,13 a 3,14 b 2,00 3,75 3,25 5,00
3 15 17 20 12 19 14 10 8 13 7 6 5 4 16 2 11 9 18 1 25.65 50.43 75.22 100.00 Acessos S im il a ri d a d e
UPGMA - Distância Euclideana
Figura 3 – Dendrograma obtido com base nas medidas de similaridade genética e método de agrupamento UPGMA a partir da distância Euclidiana, considerando os seis descritores morfoagronômicos avaliados nos 20 acessos* de Hippeastrum. Fortaleza-CE, 2018.
*/Legenda dos acessos consta na Tabela 1.
Tabela 7 - Médias e valores referentes aos descritores morfológicos avaliados em grupos de Hippeastrum
formados a partir de dendrograma obtido pelo método UPGMA. Fortaleza-CE, 2018.
Fonte: própria autoria.
1/comprimento de folhas – CFO; largura de folhas – LFO; taxa de recobrimento do solo – TRS; ocorrência de
plantas daninhas – OPD; ataque de insetos – AIN; folhas secas – FSE.
Grupo CFO (cm) 1 LFO (cm) TRS OPD AIN FSE
1 26,93 1,69 1,05 4,16 4,71 3,22
2 19,33 0,84 1,00 4,00 3,33 1,67
3 26,21 1,63 2,20 4,87 4,17 3,93
4 35,44 2,87 1,50 3,88 3,63 5,00
5 20,28 2,05 1,00 2,40 5,00 2,00
6 4,30 1,00 1,00 5,00 5,00 5,00
7 25,70 6,06 1,67 4,67 5,00 5,00
O grupo 3 foi composto por três acessos da espécie Hippeastrum sp. (GPS (2)
16111, GPS 13432 e MCA (1) 446) que demonstraram similaridade fenotípica quanto a TRS (2,00 a 2,60), agrupando quase todos os acessos que tiveram os maiores valores nesse descritor; e quanto a OPD apresentando valores de 4,00 a 5,50.
O valor de CFO foi a principal semelhança apresentada entre os acessos que compuseram o grupo 4 (GPS (4) 8227 e MFS 1702). As folhas destas plantas tinham comprimentos variando de 33 a 37,75 cm, foram as folhas de maior comprimento entre todos os acessos. Somente o acesso GPS 5717 da espécie Hippeastrum cf. puniceum compôs o
grupo 5, sua principal diferença dos demais grupos, foi o baixo valor para o o caractere OPD (2,40), que foi menor entre todos os acessos.
O grupo 6 foi formado pelo acesso GPS 15808 da espécie Hippeastrumpuniceum
e se mostrou fenotipicamente diferente dos demais principalmente por apresentar folhas muito curtas (4,30 cm), contudo demonstrou que não sofre ataques de insetos, nem apresenta ocorrência de plantas daninhas. O acesso ACA (6) 56 formou o último grupo diferenciando-se pela maior lagura de suas folhas (6,06 cm), também apresentou valor 5,00 para os caracteres AIN e FSE (Tabela 7).
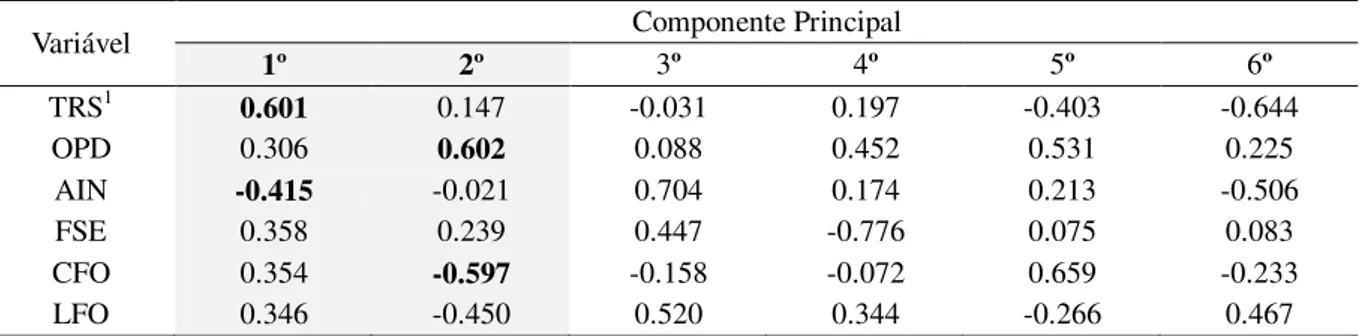
A partir do método de análise por componentes principais (PCA) foi possível a visualização da relação entre os acessos e variáveis (Figura 4). Os pesos nas duas primeiras componentes principais representaram aproximadamente 73,10% das informações contidas nos valores médios das seis variáveis morfoagronômicas.
As variáveis que tiveram maior influência no 1º componente (eixo horizontal) foram TRS e AIN e as que tiveram maior influência no 2º componente (eixo vertical) foram OPD e CFO. Esse fato também pode ser ressaltado quando se observa o peso de cada um dos seis caracteres nos componentes principais (Tabela 8). As variáveis que apresentaram os maiores comprimentos dos vetores foram TRS, AIN, OPD e CFO isto é, foram os caracteres que apresentaram maior variabilidade entre os acessos e foram justamente o que mais pesaram no 1º e 2º componentes principais. Os vetores de menor tamanho foram FSE e LFO, demonstrando menor variabilidade entre os acessos nessas variáveis.
Figura 4 – Gráfico de componente principal englobando os acessos de Hippeastrum e o biplot das características
avaliadas. Fortaleza-CE, 2018.
Fonte: própria autoria.
Comprimento de folhas – CFO; largura de folhas – LFO; taxa de recobrimento do solo – TRS; ocorrência de plantas daninhas – OPD; ataque de insetos – AIN; folhas secas – FSE.
Tabela 8 - Peso de cada um dos 6 descritores no primeiro e segundo componente principal para Hippeastrum.
Fortaleza-CE, 2018.
Variável Componente Principal
1º 2º 3º 4º 5º 6º
TRS1 0.601 0.147 -0.031 0.197 -0.403 -0.644
OPD 0.306 0.602 0.088 0.452 0.531 0.225
AIN -0.415 -0.021 0.704 0.174 0.213 -0.506
FSE 0.358 0.239 0.447 -0.776 0.075 0.083
CFO 0.354 -0.597 -0.158 -0.072 0.659 -0.233
LFO 0.346 -0.450 0.520 0.344 -0.266 0.467
Fonte: própria autoria.
1/comprimento de folhas – CFO; largura de folhas – LFO; taxa de recobrimento do solo – TRS; ocorrência de
plantas daninhas – OPD; ataque de insetos – AIN; folhas secas – FSE.
Os vetores de CFO e LFO correlacionaram-se positivamente, assim como aconteceu em Habranthus. Os acessos que apresentavam folhas mais compridas também
possuíam folhas mais largas, os principais exemplos desta correlação são os acessos GPS (4) 8227, LBB (6) 1502 e MFS 1702 (Tabela 6). O vetor de OPD apresentou correlação inexistente com os vetores CFO e LFO, demonstrando que o tamanho das folhas não se relaciona com a ocorrência de plantas daninhas.
O método de análise por componentes principais (PCA) também apresentou concordância com a análise de agrupamento observados no dendograma (UPGMA), sendo possível observar a proximidade entre os acessos que fazim parte do mesmo grupo. Assim, os acessos foram distribuídos de acordo com variáveis que mais influenciaram os mesmos, sendo outra maneira de observar a variablidade fenotipica entre os acessos.
Os vetores correspondentes aos caracteres OPD, FSE e TRS influenciaram a distribuição dos acessos GPS 13432, MCA (1) 446, GPS (2) 16111 e ACA (5) 49 no 1º quadrante. O vetor correspondente ao caractere AIN influenciou a distribuição dos acessos nos 2º e 3º quadrantes, com excesão dos acessos GPS 15808 e GPS 5717. O acesso GPS 15808 da espécie Hippeastrum puniceum foi disposto na extremidade superior do 2º
quadrante por possui o menor CFO entre todos os acessos de Hippeastrum do estudo. Então
sua disposição foi oposta a direção do vetor correspondente ao comprimento de folhas.
O acesso GPS 5717 da espécie Hippeastrum cf. puniceum foi disposto na
extremidade inferior do 3º quadrante, pois apesar de não possuir folhas atacadas por insetos, apresenta considerável ocorrência de plantas daninhas e folhas secas, por isso está localizado no sentido oposto dos vetores OPD e FSE.
Os vetores correspondentes aos caracteres CFO e LFO influenciaram a distribuição dos acessos ACA (6) 56, MFS 1702, GPS (4) 8227 no 4º quadrante. Esses acessos foram os que possuiam as folhas de maior comprimento ou largura (Tabela 6).
De forma geral, os acessos de Hippeastrum possuem uma alta variabilidade
6 CONCLUSÕES
Existe divergência genética entres os acessos de Habranthus e os descritores taxa de
recobrimento do solo, comprimento de folhas, índice de folhas secas e largura de folhas são os que melhor caracterizam este gênero.
Existe divergência genética entres os acessos de Hippeastrum e os descritores taxa de
REFERÊNCIAS
ALVES-ARAÚJO, A.; SANTOS, F. A.; ALVES, M. Caracterização palinológica de espécies de Amaryllidaceae sensu stricto ocorrentes no nordeste brasileiro. Acta Botanica Brasilica, v. 21, p. 967-976, 2007.
ALVES-ARAÚJO, A.; PESSOA, E.; ALVES, M. Caracterização morfoanatômica de espécies de Amaryllidaceae e Alliaceae do Nordeste Brasileiro. Revista Caatinga, v. 25, n. 4, p. 68-81, 2012.
AMARAL et al., 2007. Conservação in vitro de germoplasma indexado de três cultivares de
amarílis (Hippeastrum Herb.). Ornamental Horticulture v. 13, n.2, p. 113-120, 2007
AMARAL, A.C. Habranthus Herb.(Amaryllidaceae) no Brasil: estudo taxonômico,
caracterização morfológica e relações filogenéticas. 167 f., il. Tese (Doutorado em Botânica)— Universidade de Brasília, Brasília, 2012.
AMARAL-LOPES, A. C.; CAVALCANTI, T. B. Habranthus (Amaryllidaceae) of Brasil.
Rodriguésia, v. 66, n. 1, p. 203-220, 2015.
ANDRADE, Jean Paulo de et al. Investigação química em Hippeastrum vittatum (l'hér.)
herbert. Salão de Iniciação Científica (15.: 2003: Porto Alegre). Livro de resumos. Porto Alegre: UFRGS, 2003., 2003.
APG III. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society 161: 105-121.
BOLIVAR Q., Y.; ALVARENGA, R.C. Potencial de plantas de cobertura para recobrimento do solo e supressão de plantas daninhas. In: CONGRESSO BRASILEIRO DE MILHO E SORGO, 23., 2000, Uberlândia, MG. Anais. Uberlândia: SBMS, 2000. I CD-ROM
BURLE, M.L.; OLIVEIRA, M. Manual de Curadores de Germoplasma-Vegetal: Caracterização Morfológica. Embrapa Recursos Genéticos e Biotecnologia-Documentos (INFOTECA-E), 2010.
CAVALCANTE, M.Z.B. et al. Potencial ornamental de espécies do Bioma Caatinga.
Comunicata Scientiae, v.8, n.1, p.43, 2017.
CASTRO, C.E.F; MOREIRA, S.R.; DE CASTRO, C.R.; SOUZA, F.V.D.; LOGES, V.; GONÇALVES, C. Avaliação de espécies de Costaceae para uso ornamental. Ornamental Horticulture, v.17, n.1, p.63-74, 2011.
CASTRO, A.D.; TANIGUCHI, C.A.K.; SOUZA, F.H.; SILVA, T.F.; CAFÉ, F.B.; ARAGÃO, F.A.S.; LOGES, V. Avaliação do germoplasma nativo brasileiro de Paspalum como gramado para fins de paisagismo. Acta horticulturae, n.1104, p.505-510, 2015.
COSTA A.M.; A.; SPEHAR, R.S. Base genética da diversidade. Conservação de recursos genéticos no Brasil. Brasília, DF: Embrapa, p. 28-59, 2012.
COSTA, T.S.; DA SILVA, A.V.C.; LEDÓ, A.S.; DOS SANTOS, A.R.F.; SILVA JÚNIOR, J.F. Diversidade genética de acessos do banco de germoplasma de mangaba em Sergipe. Pesquisa Agropecuária Brasileira, v.46, n.5, p. 499-508, 2011.
CRUZ, C.D.; FERREIRA, F.M.; PESSONI, L.A. Biometria aplicada ao estudo da diversidade genética. Visconde do Rio Branco: Suprema, p. 620, 2011.
CRUZ, C.D. Genes: a software package for analysis in experimental statistics and quantitative genetics. Acta Scientiarum. Agronomy, v. 35, n. 3, p. 271-276, 2013.
DUARTE, J. B. Princípios e utilização de técnicas multivariadas no melhoramento de plantas. 1997. 69 f. Monografia (Bacharelado em Genética) - Escola Superior de Agricultura
‘Luiz de Queiroz’, Piracicaba, 1997.
Herb.(Amaryllidaceae). 1996.
DUTILH, J.H.A. Liliaceae sl. Annals of Botany, v. 88, p.1057-1069, 2005.
DUTILH, J.H.A. et al. Amaryllidaceae. In: MARTINELLI, G.; MORAES, M.A. Livro
vermelho da flora brasileira. Rio de Janeiro: Andrea Jakobson Estúdio/Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, p. 126-139, 2013.
DUTILH, J.H.A.; OLIVEIRA, R. S. Amaryllidaceae in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de Janeiro. RJ, 2015.
EUROPEAN COMMISSION. Agriculture and rural development, statistics of live plants and products of floriculture, 2017. Disponível em https://ec.europa.eu/agriculture/fruit-and-vegetables/product-reports/flowers_en. Acesso em 05/05/2018.
FERRANTE, A.; TRIVELLINI, A; SCUDERI, D.; ROMANO, D.; VERNIERI, P. Post-production physiology and handling of ornamental potted plants. Postharvest Biology and Technology, v. 100, p. 99-108, 2015.
FERNÁNDEZ, A.C.; MARINANGELI, P.; CURVETTO, N.; FACCIUTO, G. Reproductive
biology of Habranthus tubispathus. Acta Horticulturae, v.1000, p.183-188, 2013.
FIGÀS, M.R.; PROHENS, J; CASANOVA, C.; FERNÁNDES-DE-CÓRDOBA, P; SOLER, S. Variation of morphological descriptors for the evaluation of tomato germplasm and their stability across different growing conditions. Scientia Horticulturae, v. 238, p. 107–115, 2018.
FLORA DO BRASIL 2020. Flora do Brasil – Hippeastrum Herb. Jardim Botânico do Rio
de Janeiro. Disponível em: <http://reflora.jbrj.gov.br/reflora/floradobrasil/FB4354>. Acesso em: 22 mar. 2018.
GRANATE, M. J.; CRUZ, C. D.; CECON, P. R.; PACHECO, C. A. P. A análise de fatores na predição de ganhos por seleção em milho (Zea mays L.). Acta Scientiarum, v. 23, n. 5, p. 1271-1279, 2001.
IBRALFLOR - INSTITUTO BRASILEIRO DE FLORICULTURA. Números do Setor. Disponível em: < http://www.ibraflor.com/site/wp-content/uploads/2017/11/release-imprensa-ibraflor-10-2017.pdf >. Acesso em: 06 jun. 2018
JUNQUEIRA, A.H; PEETZ, M.S. Flores e plantas ornamentais do Brasil, São Paulo: SEBRAE, v.2, 2015. 100p.
JUNQUEIRA, A.H; PEETZ, M.S. Agronegócio da floricultura na economia brasileira. In: Agrianual 2017- Anuário da Agricultura Brasileira, p.267-268, São Paulo: IEG/FNP, 2016.
JUNQUEIRA, A.H.; PEETZ, M. Brazilian consumption of flowers and ornamental plants: habits, practices and trends. Ornamental Horticulture, v.23, n.2, p.178-184, 2017.
KARASAWA, M.M. O uso de recursos genéticos na produção de cultivares melhoradas com vistas a atender os desafios impostos pela mudança climática. Revista RG News, n.2, v.1, p.95-97, 2016.
KILGORE, M.B.; KUTCHAN, T.M. The Amaryllidaceae alkaloids: biosynthesis and methods for enzyme discovery. Phytochemistry Review, v.15, p. 317-337, 2016.
MAGNUSSON, W.E. et al. O Programa de Pesquisa em Biodiversidade. IN: Peixoto, A.L.;
Luz, J.R.P.; Brito, M.A. Conhecendo a biodiversidade / Organizadores. – Brasília: MCTIC/CNPq, PPBio, 196 p., 2016.
MEEROW, A.W.; SNIJMAN, D.A. Amaryllidaceae. In: Flowering Plants Monocotyledons. Springer, Berlin, Heidelberg, p. 83-110, 1998.