UNIVERSIDADE FEDERAL DE MINAS GERAIS
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE BIOLOGIA GERAL
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA
DISSERTAÇÃO DE MESTRADO
Influência de polimorfismos da catecol-O-metiltransferase no
desempenho da memória de trabalho de crianças saudáveis
ORIENTADA: Máira Pedroso de Almeida
ORIENTADORA: Profa. Dra. Maria Raquel Santos Carvalho
BELO HORIZONTE
Máira Pedroso de Almeida
Influência de polimorfismos da catecol-O-metiltransferase no
desempenho da memória de trabalho de crianças saudáveis
Dissertação apresentada ao programa de Pós-Graduação em Genética do Departamento de Biologia Geral do Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, como requisito parcial para obtenção do grau de Mestre em Genética.
Área de Concentração: Genômica e Bioinformática
Orientadora: Profa. Dra. Maria Raquel Santos Carvalho.
BELO HORIZONTE
043 Almeida, Máira Pedroso de
Influência de polimorfismos da catecol-O-metiltransferase no desempenho da memória de trabalho de crianças saudáveis [manuscrito] / Máira Pedroso de Almeida. – 2015.
82 f. : il. ; 29,5 cm.
Orientador: Maria Raquel Santos Carvalho.
Dissertação (mestrado) – Universidade Federal de Minas Gerais, Departamento de Biologia Geral.
1. Memória imediata - Teses. 2. Dopamina – Teses. 3. Haplótipos. 4. Catecol-O-metiltransferase. 5. Polimorfismo (Genética) - Teses. 6. Genética – Teses. I. Carvalho, Maria Raquel Santos. II. Universidade Federal de Minas Gerais.
Departamento de Biologia Geral. III. Título.
AGRADECIMENTOS
À Deus, por me dar forças e confiança para sempre seguir a diante.
Aos meus pais pelo carinho e apoio incondicional.
À Universidade Federal de Minas Gerais, em especial ao Departamento de
Biologia Geral e ao Programa de Pós-Graduação em Genética, pela oportunidade de
realização de mais esta etapa na minha vida.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
pela concessão da bolsa de estudos.
À minha orientadora, Professora Maria Raquel Santos Carvalho, pela
orientação, confiança, oportunidades, incentivos e ensinamentos.
Ao Laboratório de Neuropsicologia do Desenvolvimento, em especial a
Annelise Júlio Costa e ao Prof. Dr. Vitor G. Haase, pela parceria no projeto.
Ao Laboratório de Genética Celular e Molecular, em especial ao Rodrigo Dias
e a Mariana Santana, pelo auxílio nas análises de genotipagem.
Ao Peter Laspina pelo auxílio na correção de trabalhos, resumos e artigos em
inglês, bem como pelos conselhos.
Aos membros da banca de defesa.
Aos meus colegas do Laboratório de Genética Humana e Médica pelo apoio e
amizade.
Sumário
Lista de figuras... vii
Lista de tabelas... ix
Lista de abreviaturas ... x
Resumo ... 1
Abstract ... 3
Introdução ... 5
Memória de trabalho ... 5
Síndromes que cursam com déficits na MT ... 8
Gene COMT ... 10
Estrutura e função ... 10
Polimorfismos ... 13
Estudos de associação ... 16
Necessidades atuais ... 18
Justificativa e relevância ... 19
Referências bibliográficas ... 20
Objetivos ... 29
Objetivo geral ... 29
Objetivo específico ... 29
Artigo: Influence of Catechol-O-methyltransferase Polymorphisms on Working Memory Performance of Healthy Children ... 30
Introduction ... 30
Material and methods ... 32
Results ... 35
Discussion ... 38
Conclusion ... 41
References ... 41
Supplementary material ... 45
Considerações finais ... 53
Referências bibliográficas ... 55
Anexos ... 56
Produção bibliográfica ... 56
Parecer do Comitê de Ética ... 57
LISTA DE FIGURAS
Introdução
Figura 1. Modelo de memória de trabalho baseado em quatro componentes. Este modelo compreende um sistema de controle, o executivo central, dois sistemas de armazenamento, os componentes viso-espacial e fonológico, bem como um buffer episodic, o qual permite que informações dos dois sistemas de armazenamento sejam combinadas com informações da memória de longo prazo.
FONTE: adaptada de Baddeley, 2003...6
Figura 2. O gene COMT e seus transcritos. O gene COMT é formado por seis éxons (representados pelas caixas cinza) e dois promotores, P2 e P1, os quais são responsáveis por controlar a expressão dos transcritos de 1.5 e 1.3 kb, respectivamente. O gene COMT contém alguns polimorfismos de interesse, como por exemplo, o rs2075507 e o rs4680. As posições de início da tradução da MB-COMT e S-COMT, bem como o códon de terminação da tradução e a cauda poli-A (pA) estão indicados.
FONTE:adaptada de Zhu, 2002...11
Figura 3. Transmissão dopaminérgica no córtex pré-frontal (CPF) e corpo estriado. No CPF a COMT é a principal responsável pela remoção da dopamina na fenda sináptica, enquanto em outras regiões cerebrais, como o corpo estriado, a proteína transportadora de dopamina (DAT- dopamine transporter) exerce esta função.
FONTE: adaptada de Tunbridge et al., 2006...12
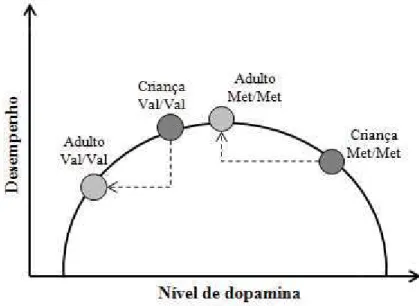
Figura 4. Relação em forma de U invertido entre nível de dopamina no córtex pré-frontal e desempenho da memória de trabalho.
FONTE: adaptada de Tunbridge et al., 2006...13
Figura 5. Efeito da atividade da COMT na relação em forma de U invertido entre nível de dopamina e função do córtex pré-frontal (CPF). Homozigotos Val estão associados a níveis subótimos de dopamina, enquanto homozigotos Met apresentam níveis próximos aos ideais. Fonte: adaptada de Tunbridge et al., 2006...14
Figura 6. Efeito da atividade da COMT na relação em forma de U invertido entre nível de dopamina e função do córtex pré-frontal (CPF). Homozigotos Valpodem ser deslocados para próximo do ápice da curva em razão da redução da atividade da COMT (presença do alelo G do SNP rs2075507). Em contrapartida, a redução da atividade da COMT em homozigotos Met resulta em níveis de dopamina exageradamente altos, prejudicando assim a função do CPF.
FONTE: adaptada de Tunbridge et al., 2006...16
Figura 7. Alterações na relação entre genótipos da COMT e desempenho cognitivo em função da idade. Na infância, devido à presença de maiores níveis de dopamina no córtex pré-frontal (CPF), homozigotos Val apresentam melhor desempenho cognitivo que homozigotos Met.
LISTA DE FIGURAS
Artigo
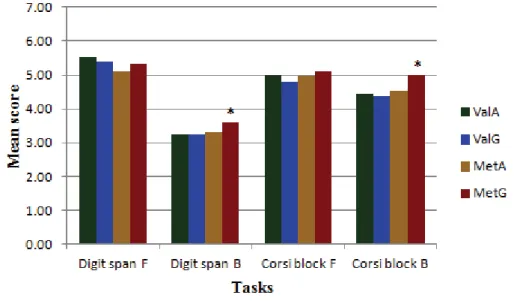
Figure 1. Performance of girls on working tasks, by rs4680 genotype. *P≤ 0.05. F = forward,
B = backward………36
Figure 2. Performance of boys on working memory tasks, by rs4680 genotyope. F = forward,
B = backward………37
Figure 3. Performance of girls on working memory tasks, by rs4680 - rs2075507 haplotype. *
P≤ 0.05. F = forward, B = backward. For estimated haplotyte frequencies see Supplementary
Table 1………..……….38
Figure 4. Performance of boys on working memory tasks, by rs4680 - rs2075507 haplotype. *
P≤ 0.05. F = forward, B = backward. For estimated haplotyte frequencies see Supplementary
Table 1………..………...…..39
Supplementary figure 1. Distribution of the test scores……….47
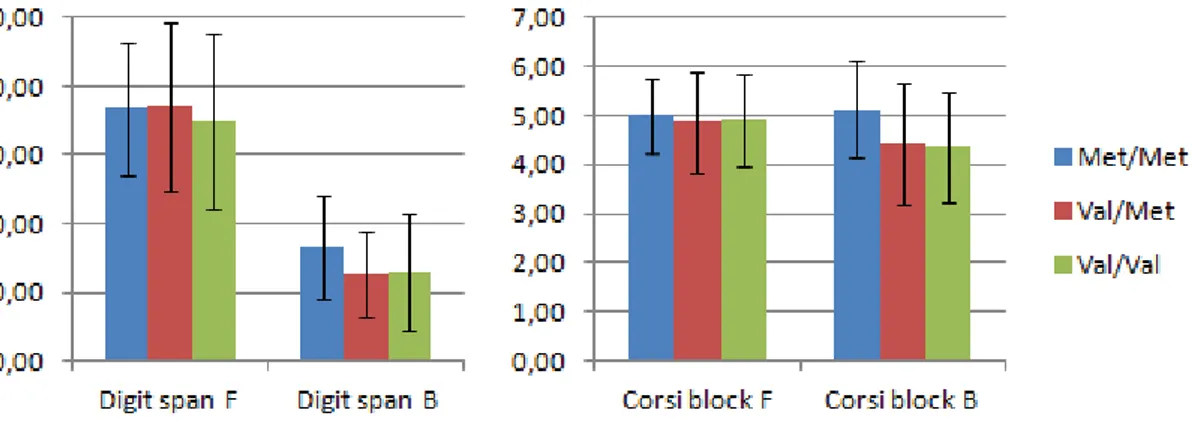
Supplementary figure 2. Performance of whole sample on working memory tasks, by rs4680
genotype. F = forward, B = backward ………...…..47
Supplementary figure 3. Performance of younger children (7-9-year-old) on working memory tasks, by rs4680 genotype. F = forward, B = backward………48
Supplementary figure 4. Performance of older children (10-12-year-old) on working memory tasks, by rs4680 genotype. F = forward, B = backward……….…48
Supplementary figure 5. Performance of whole sample on working memory tasks, by rs2075507 genotype. F = forward, B = backward……….49
Supplementary figure 6. Performance of girls on working memory tasks, by rs2075507
genotype. F = forward, B = backward………..49
Supplementary figure 7. Performance of boys on working memory tasks, by rs2075507
genotype. F = forward, B = backward………..50
Supplementary figure 8. Performance of younger children (7-9-year-old) on working memory tasks, by rs2075507 genotype. F = forward, B = backward……….………….50
Supplementary figure 9. Performance of older children (10-12-year-old) on working memory tasks, by rs2075507 genotype. F = forward, B = backward….………51
Supplementary figure 10. Performance of whole sample on working memory tasks, by rs4680 - rs2075507 haplotype. *P≤ 0.05. F = forward, B = backward. For estimated haplotyte
frequencies see Supplementary Table 1………..………..51
Supplementary figure 11. Performance of younger children (7-9-year-old) on working memory tasks, by rs4680 - rs2075507 haplotype. F = forward, B = backward. For estimated haplotyte frequencies see Supplementary Table 1………52
LISTA DE TABELAS
Artigo
Table 1. Demografic statistics for different genotypes………35
Table 2. Estimated haplotyte frequencies……….………35
Table 3. Genotype effects on working memory tasks for sex groups……….…………36
Table 4. Haplotype effects on working memory tasks for sex groups………38
Supplementary table 1. Estimated haplotyte frequencies for sex and age groups….………45
Supplementary table 2. Genotype effects on working memory tasks for the whole sample
………...45
Supplementary table 3. Genotype effects on working memory tasks for age groups……....46
Supplementary Table 4. Haplotype effects on working memory tasks for the whole sample ...46
LISTA DE ABREVIATURAS
Introdução e Considerações finais
COMT catechol-O-methyltransferase CPF córtex pré-frontal
DAT dopamine transporter DL dificuldade de leitura
DM dificuldade matemática
FMRP fragile X mental retardation protein FMR1 fragile X mental retardation 1
Met metionina
MT memória de trabalho
QI quociente de inteligência
SNP polimorfismo de nucleotídeo único
Val valina
Artigo
COMT catechol-O-methyltransferase LD linkage disequilibrium
Met methionine
PFC prefrontal cortex
SNP single nucleotide polymorphism
Val valine
Resumo
Memória de trabalho refere-se ao armazenamento temporário e a manipulação
de informações, os quais são essenciais para os processos de aprendizagem. Nos
últimos anos, muitos estudos identificaram o gene catechol-O-methyltransferase (COMT) como um importante modulador da memória de trabalho. Este gene,
localizado no cromossomo 22q11.2, codifica a enzima COMT, que possui o papel de
metabolizar a dopamina presente nas fendas sinápticas do córtex pré-frontal. Como
consequência deste papel, a COMT tem sido apontada como a principal determinante
das funções dependentes do córtex pré-frontal, sendo a memória de trabalho uma
delas. Múltiplos polimorfismos já foram descritos no gene COMT, mas o
polimorfismo rs4680, também chamado de Val158Met, é o mais investigado na
literatura. Este polimorfismo de nucleotídeo único (SNP), localizado no éxon 4,
consiste em uma substituição G → A, a qual por sua vez acarreta na substituição de
uma valina (Val) por uma metionina (Met). O alelo Met, por apresentar menor
termoestabilidade, resulta em menor atividade da COMT. Assim, quando
comparados a homozigotos Val158Val, homozigotos Met158Met exibem maiores
níveis de dopamina no córtex pré-frontal. Como os alelos são codominantes,
heterozigotos apresentam níveis intermediários de atividade da COMT. Os
homozigotos Met158Met apresentam níveis de dopamina no córtex pré-frontal
próximos aos considerados ideais, por isso, espera-se que eles desempenhem melhor
que heterozigotos e homozigotos Val158Val em tarefas de memória de trabalho. No
entanto, diversos estudos não conseguiram identificar este padrão. Existem
evidências de que os níveis de dopamina decrescem a partir do início da puberdade, o
que acarretaria em diferentes efeitos dos genótipos da COMT em diferentes idades, e
explicaria a falta de concordância entre os estudos. Embora alguns trabalhos
conseguiram identificar estas diferenças de idade, diferenças de efeito dos genótipos
devido ao sexo ainda não foram identificadas. Recentemente, um SNP no promotor
P2 do gene COMT, o rs2075507 (A287G), emergiu como um possível modulador
adicional da memória de trabalho. Este polimorfismo, que apresenta um baixo efeito
sobre a COMT, interage com o Val158Met para modular a eficiência do córtex
pré-frontal. Dependendo com qual dos alelos do polimorfismo Val158Met os alelos do
SNP rs2075507 estejam associados, eles podem beneficiar ou prejudicar a memória
efeitos dos polimorfismos rs4680 e rs2075507 sobre a memória de trabalho, em uma
amostra de crianças entre 6 e 14 anos de idade, bem como averiguar a existência de
interação entre estes SNPs por meio da análise de haplótipos. Além disso, foi
investigada a influência do sexo e da idade sobre os efeitos exercidos por estes
polimorfismos. Ao estudar o SNP rs4680 separadamente, os resultados
demonstraram que para meninos e crianças mais novas (6 a 9 anos), homozigotos
Val158Val apresentaram melhor desempenho da memória de trabalho. Já para
meninas e crianças mais velhas (10 a 14 anos), os homozigotos Met158Met
apresentaram melhor desempenho. Estes dados revelam, pela primeira vez, que além
da idade, o sexo também afeta os padrões de associação do SNP Val158Met. Esta
influência do sexo possivelmente se deve às diferentes idades de início da puberdade
para cada gênero. Não foram detectados efeitos para o polimorfismo rs2075507. Na
análise conjunta dos SNPs, o haplótipo ValG previu melhor desempenho da memória
de trabalho em meninos e crianças mais novas, enquanto o haplótipo MetG previu
melhor desempenho em meninas e crianças mais velhas. A presença do alelo G em
ambos os haplótipos relacionados ao melhor desempenho da memória de trabalho é
um indicativo de que talvez o rs2075507 não seja o real polimorfismo interagindo
com o rs4680, mas sim algum outro polimorfismo próximo a ele. Em geral, este
estudo sugere que mais do que a idade, o estágio puberal possa ser um fator
importante a ser considerado nas análises de associação envolvendo o SNP Val158Met. Além disso, foi apontada a necessidade de se analisar outros
polimorfismos próximos ao rs2075507, uma vez que um deles pode ser o verdadeiro responsável pela modulação da atividade da COMT. Associados a pesquisas futuras,
os resultados aqui obtidos poderão auxiliar no desenvolvimento de estratégias
educacionais e psicológicas eficazes para a melhor assistência aos alunos com
Abstract
Working memory refers to the temporary storage and manipulation of
information, which are essential to the learning processes. In the last few years, a
number of studies have identified the catechol-O-methyltransferase (COMT) gene as
an important working memory modulator. This gene, located on chromosome 22q11.2, codes the COMT enzyme which catabolizes dopamine in the synaptic clefts
of the prefrontal cortex. COMT has been pointed out as one of the major determinant of prefrontal cortex-dependent functions, such as working memory. Multiple
polymorphisms have been described in the COMT gene; but, rs4680, also known as
Val158Met, is the most investigated in the literature. This single nucleotide
polymorphism (SNP), located in exon 4, consists of a G → A substitution which, in
turn, causes a substituon of a valine (Val) to a methionine (Met). Due to its lower
thermostability, the Met allele results in lower COMT activity. Thus, compared to
Val158Val homozygotes, Met158Met homozygotes have higher dopamine levels in
the prefrontal cortex. Since the alleles are codominant, heterozygotes have
intermediate levels of COMT activity. The Met158Met homozygotes have near
optimal prefrontal cortex dopamine levels; hence, they are expected to perform better
than both heterozygotes and Val158Val homozygotes in working memory tasks.
However, several studies have failed to identify this pattern. There is some evidence
that dopamine levels begin to decrease with the onset of puberty, which would result
in different effects of COMT genotype at different ages, and, thus, would explain the
lack of agreement among studies. Although some studies were able to identify age
differences, sex differences have not yet been identified. Recently, a SNP in the P2
promoter of the COMT gene, rs2075507 (A287G), emerged as a possible additional
modulator of working memory. This polymorphism, that has a minor effect on
COMT activity, interacts with Val158Met to modulate prefrontal cortex efficiency.
Depending on which of the Val158Met alleles the rs2075507 alleles are associated,
they might benefit or impair working memory. Given this, the aims of this study
were to investigate the effects of the rs4680 and rs2075507 polymorphisms on
working memory in a sample of children aged 6 to 14 years old, as well as to
examine the possibility of interaction between both SNPs using haplotype analysis.
In addition, the influences of sex and age on the effects exerted by these
boys and younger children (6 to 9 years), Val158Val homozygotes presented better
working memory performance. For girls and older children (10 to 14 years),
Met158Met homozygotes presented better performance. These data point to, for the
first time, that in addition to age, sex also affects the association patterns of the
Val158Met SNP. This sex influence is possibly due to the different onset ages of
puberty for males and females. No effects were detected for the rs2075507
polymorphism. Analyzing both SNPs together, the ValG haplotype predicted better
working memory performance in boys and younger children, while the MetG
haplotype predicted better performance in girls and older children. The presence of
the G allele in both haplotypes related to better working memory performance, is an
indication that perhaps the rs2075507 polymorphism is not the one actually
interacting with rs4680; but, instead, another polymorphism near rs2075507 might
actually be interacting with rs4680. In summary, the present study suggests that
puberty, more than age, is an important factor to be considered in Val158Met SNP
association analyses. In addition, the necessity to analyze other polymorphisms near
rs2075507 was pointed out, since one of these other polymorphisms might really be
responsible for modulating COMT activity. Associated with future research, the
results obtained here may help in the development of effective educational and
psychological strategies for improving the assitence of children with learning
1. Introdução
1.1. Memória de trabalho
A habilidade humana de interagir com o ambiente baseia-se principalmente na
capacidade de manter informações ativas e disponíveis para consulta durante um
determinado momento. Tal capacidade é executada por meio da memória de trabalho
(MT), que nada mais é do que um armazenamento temporário e dinâmico de
informações a serem utilizadas ou manipuladas constantemente (Karlsgodt et al.,
2011). A MT apresenta papel fundamental na cognição e no desempenho cognitivo,
permitindo a integração de dados a partir de estímulos percebidos instantaneamente,
memória de longo prazo e processos do pensamento (Vogler et al., 2014).
Informações mantidas na MT podem ser aplicadas em situações rotineiras, utilizadas
na resolução de problemas ou empregadas como gatilho para a memória de longo
prazo. Ela é crítica para o desempenho nas atividades de leitura, escrita,
planejamento e comunicação coerente. Como são requeridas constantemente, danos a
uma ou mais destas habilidades são capazes de levar a graves perturbações na vida
diária de um indivíduo (Karlsgodt et al., 2011).
Ao longo das últimas décadas, o grande número de trabalhos na área gerou um
consenso de que a MT constitui uma característica complexa, onde o volume de
armazenamento de conteúdo transitório varia de um indivíduo para outro devido, em
parte, a fatores genéticos (Vogler et al., 2014). Apesar do desempenho da MT ser
herdável, o mecanismo biológico pelo qual ele é transmitido não é conhecido. A
ampla rede gênica que influencia a MT pode afetar o funcionamento neuronal dentro
de regiões cerebrais específicas ou a coordenação da atividade nestas regiões (Ando
et al., 2001; Chen et al., 2009; Karlsgodt et al., 2011). Utilizando metodologia
baseada no estudo de polimorfismos de nucleotídeo único (single nucleotide
polymorphisms - SNPs), Vogler et al. (2014) obtiveram estimativas de herdabilidade
variando entre 31 e 41% para o fenótipo MT. Trabalhos prévios, com base em
famílias e gêmeos, observaram estimativas de herdabilidade entre 15 e 72%
(Karlsgodt et al., 2010; Vinkhuyzen et al., 2010; Wilson et al., 2011; Lee et al.,
Atualmente, o modelo de MT mais aceito é o baseado em três componentes
descrito por Baddeley e Hitch (1974). Este modelo é formado pelo componente
fonológico, que é responsável por manter informação verbal, pelo componente
viso-espacial, o qual mantém informação viso-espacial, e pelo componente executivo central, o
qual dirige a atenção, integra informações e coordena os dois componentes anteriores
(Baddeley, 2003b; Karlsgodt et al., 2011) (Fig. 1). O elemento fonológico pode ser
decomposto em dois subelementos: o sistema de armazenamento temporário, o qual
mantém informações verbalmente codificadas por alguns segundos, e o sistema de
ensaio subvocal, que atualiza as informações armazenadas no primeiro subelemento
para que elas não sejam perdidas (Baddeley, 2003a; b; Karlsgodt et al., 2011). O
componente viso-espacial tem a função de integrar dados espaciais, visuais e
cinestésicos em uma representação unificada, que pode ser temporariamente
armazenada e manipulada. Por fim, o componente executivo central é responsável
pelo controle da atenção e pelo processamento de tarefas cognitivas. Ele possui a
função de coordenar o desempenho em diferentes tarefas (p. ex. simultaneamente
armazenar e processar a informação), optar por uma tarefa ou estratégia, atentar para
informação relevante e inibir informação irrelevante, e ativar e recuperar informação
da memória de longo prazo (Baddeley, 2003a; Corso e Dorneles, 2012).
Recentemente, este modelo de MT descrito por Baddeley e Hitch (1974) foi
atualizado para incluir um quarto componente, o Buffer episódico. Este novo elemento é capaz de integrar informações da memória de longo prazo e dos
componentes fonológico e viso-espacial (Baddeley, 2000).
Figura 1. Modelo de memória de trabalho baseado em quatro componentes. Este modelo compreende um sistema de controle, o executivo central, dois sistemas de armazenamento, os componentes viso-espacial e fonológico, bem como um buffer episodic, o qual permite que informações dos dois sistemas de armazenamento sejam combinadas com informações da memória de longo prazo.
Informações mantidas na MT são em grande parte transitórias, ao passo que
informações retidas por extensos períodos de tempo são apoiadas por diferentes
processos de memória de longo prazo. O córtex pré-frontal (CPF) é a principal região
cerebral envolvida na MT. Lesão desta região interfere na execução de tarefas que
demandam esta característica (Karlsgodt et al., 2011).
Uma ampla variedade de testes psicológicos foi desenvolvida para tornar o
desempenho da MT quantificável. Estes testes tipicamente consistem na
apresentação de estímulos aos indivíduos testados. Adultos/crianças passam por um
breve período de codificação da informação recebida, seguido por um período de
manutenção e/ou processamento na ausência do estímulo original, e por fim, por um
período de recuperação da informação. Como a memória é armazenada
transitoriamente, para que o indivíduo obtenha sucesso ele deve se lembrar da
informação alvo durante o período de ausência do estímulo e, em seguida, descartá-la
(Karlsgodt et al., 2011).
Os principais testes de mensuração da capacidade de MT encontrados na
literatura são: n-back, Count span, Digit span e Corsi block (Case et al., 1982;
Kessels et al., 2000; Figueiredo, 2002; Conway et al., 2005). O teste n-back
apresenta uma lista de itens, na qual o indivíduo testado deve continuamente
informar se cada item é igual àquele que apareceu n itens atrás na sequência (o n
tipicamente varia de 1 a 4) (Conway et al., 2005). Por exemplo, na tarefa two-back, os indivíduos devem manter de forma contínua os últimos dois itens da lista,
atualizando este conjunto de memória a cada novo item e descartando aquele menos recente. No Count span, o sujeito avaliado deve contar em voz alta o número de
pontos vermelhos presentes em uma tela. Após a observação de múltiplas telas, o
sujeito deve informar quais quantidades foram contadas em cada uma (Case et al.,
1982). O teste Digit span apresenta dois tipos de tarefas, de ordem direta e inversa.
Na tarefa de ordem direta, o examinador dita uma sequência de números e, em
seguida, o probando deve repetir os números na mesma ordem. Já na tarefa de ordem
inversa, o probando deve repetir os números na ordem inversa, ou seja, repeti-los a
partir do último. Em ambas as tarefas, a sequência se inicia com dois números e
aumenta progressivamente à medida que o sujeito acerta as respostas (Figueiredo,
2002). Assim como o Digit span, o teste Corsi block também apresenta tarefas de
ordem direta e inversa. Na tarefa de ordem direta, o examinador toca em alguns
mesma ordem. Na tarefa de ordem inversa, o indivíduo deve tocar os blocos na
ordem inversa, iniciando do último até o primeiro. Conforme o indivíduo acerta a
sequência, o número de blocos aumenta, podendo chegar até nove (Kessels et al.,
2000).
A MT é fortemente relacionada às habilidades de aprendizagem e ao progresso
acadêmico, sendo capaz de predizer o nível escolar atual e subsequente de crianças
através dos anos escolares, em leitura e matemática (Alloway, 2009). Diversos
estudos apoiam fortemente a visão de que déficits na MT contribuem para
dificuldades de aprendizado (Alloway, 2009; Peng e Fuchs, 2014). Alloway et al.
(2005) observaram que crianças com graves problemas de aprendizagem
apresentavam maiores déficits na MT do que crianças com problemas mais leves. Em
um estudo semelhante, Alloway (2009) avaliou 64 crianças, entre 7 e 11 anos, para
memória, Quociente de Inteligência (QI), leitura e matemática. Após dois anos, estas
mesmas crianças foram retestadas para as duas últimas características. O objetivo da
pesquisa era investigar o poder preditivo da MT e do QI nas habilidades de leitura e
matemática em crianças com problemas de aprendizagem. Os resultados
demonstraram que a capacidade de MT e os conhecimentos adquiridos
anteriormente, e não o QI, foram os preditores exclusivos dos resultados de
aprendizado dois anos mais tarde.
Estudantes com déficits importantes na MT precisam se esforçar mais na sala de aula, uma vez que eles são incapazes de armazenar em mente informação suficiente
que lhes permita completar uma dada tarefa. A perda de informações cruciais a partir da MT acarreta no esquecimento de instruções que os alunos deveriam seguir
e de detalhes sobre o que eles estão fazendo. Consequentemente, estes alunos
falham em diversas atividades, o que por fim, resulta em um progresso acadêmico
inferior, quando comparado aos demais colegas de classe (Alloway et al., 2009).
1.2. Síndromes que cursam com déficits na MT
Algumas síndromes apresentam o déficit na MT como uma importante
característica fenotípica. Como exemplo, podemos citar a Síndrome de Turner,
Síndrome do X-Frágil e Síndrome Velocardiofacial.
A Síndrome de Turner resulta da perda total ou parcial do cromossomo X. Uma
das consequências desta síndrome é a falha no desenvolvimento ovariano, o que
influencia o desempenho em tarefas de memória verbal e não-verbal, e pode
contribuir para o fenótipo cognitivo associado a Síndrome de Turner. O fenótipo
cognitivo caracteriza-se, principalmente, por dificuldades na função executiva e no
processamento de informações viso-espaciais, as quais são acompanhadas por
relativa preservação das habilidades verbais (Hart et al., 2006; Murphy et al., 2006;
Bray et al., 2011). Embora a deficiência intelectual não seja uma característica
frequente, existe um alto risco de dificuldades de aprendizado dentre as meninas
afetadas. Particularmente, dificuldades de aprendizagem da matemática, as quais são
significativas e persistentes (Rovet, 1993; Mazzocco, 2009). Estudos realizados com
meninas em idade escolar encontraram as seguintes frequências de dificuldade
matemática: 7-10% entre normais e 43-55% entre afetadas (Rovet, 1993; Mazzocco,
2001).
A Síndrome do X-Frágil é causada por expansões do trinucleotídeo CGG na
região 5’-UTR do gene FMR1 (fragile X mental retardation 1) (Verkerk et al.,
1991; Yu et al., 1991). Grandes expansões (acima de 200 repetições) levam a
inibição da produção da proteína FMRP (fragile X mental retardation protein), a
qual é importante para o desenvolvimento neural. Pacientes com Síndrome do
X-Frágil apresentam déficits específicos na função executiva e MT, sendo que o
comprometimento desta última parece ser mais pronunciado em tarefas de MT
fonológicas. Estes déficits estão presentes mesmo em indivíduos assintomáticos com pré-mutação e o número de repetições CGG está diretamente correlacionado
com o grau de prejuízo da função executiva (Karlsgodt et al., 2011). A maioria dos meninos afetados por este distúrbio apresenta deficiência intelectual, que pode
variar de leve à moderada (p. ex. QI médio entre 36 e 70) (Bailey et al., 1998). Por
outro lado, 50% das meninas afetadas possuem deficiência intelectual, enquanto as
50% restantes podem apresentar deficiências cognitivas menos graves, incluindo
dificuldades de aprendizagem, ou podem não manifestar efeitos perceptíveis
(Rousseau et al., 1994; Murphy et al., 2006). A taxa de prevalência da dificuldade
de aprendizagem da matemática entre meninas sem deficiência intelectual excede a
taxa da população geral (Murphy, 2009). Murphy et al. (2006) em um estudo
longitudinal, encontraram que a porcentagem de meninas com Síndrome do
X-Frágil que apresentaram dificuldade matemática pelo menos uma vez durante
avaliações anuais, entre o jardim de infância e a terceira série, foi maior (87%) que a
pesquisas são realizadas com meninas, pouco se sabe sobre a habilidade matemática
em meninos afetados (Murphy, 2009).
A Síndrome Velocardiofacial, também chamada de Síndrome de deleção
22q11.2, envolve a perda de uma das cópias de em torno de 30 genes (Scambler et
al., 1992; Lindsay, 2001; Prescott e Scambler, 2005). As principais características
apresentadas pelas crianças afetadas incluem anormalidades velofaríngeas,
anomalias cardíacas congênitas, dismorfias faciais e dificuldades de aprendizagem.
Dentre os fenótipos neurocognitivos, destaca-se o déficit de MT, especialmente de
MT viso-espacial (Montojo et al., 2014). Durante o ensino primário, as crianças já
exibem dificuldades matemáticas consideráveis, apesar do desempenho em leitura
ser relativamente preservado (De Smedt et al., 2009). As habilidades intelectuais
são geralmente mais baixas que a média (QI igual 74), com cerca de 60% dos
afetados apresentando inteligência variando de normal a limítrofe (QI > 70) e cerca
de 40% apresentando deficiência intelectual leve (Swillen et al., 1997; Woodin et
al., 2001; De Smedt et al., 2007).
Tem sido sugerido que um ou vários genes contidos na deleção 22q11.2 possam
estar envolvidos no desenvolvimento e na função cerebral, possivelmente levando às
alterações morfofuncionais típicas desta síndrome (Maynard et al., 2003). Um dos
genes da região deletada que tem chamado bastante atenção dos pesquisadores é o
catechol-O-methyltransferase (COMT). O grande interesse neste gene provém da sua contribuição para as diferenças individuais no funcionamento cerebral, bem
como para o desempenho cognitivo na população geral (Diamond et al., 2004).
1.3. Gene COMT
1.3.1. Estrutura e função
O gene COMT possui seis éxons (sendo os dois primeiros não codantes) e é
capaz de codificar dois transcritos, de 1,3 e 1,5 Kb, a partir de dois promotores
(Tenhunen et al., 1994). As variantes transcricionais codificam aloenzimas distintas,
uma solúvel (S-COMT) e uma ligada à membrana (MB-COMT), a partir de dois
códons de início ATG. O menor transcrito, de 1,3 kb, é regulado pelo promotor P1.
Este promotor sobrepõe parcialmente a sequência codificadora da forma MB-COMT. Por sua vez, a síntese do transcrito de 1,5 kb é regulada pelo promotor P2,
associada à membrana, enquanto o transcrito de 1,3 kb codifica somente a forma
solúvel (Fig. 2) (Männistö e Kaakkola, 1999; Zhu, 2002).
A S-COMT apresenta 221 aminoácidos, ao passo que a MB-COMT possui, além
destes, uma extensão de 50 aminoácidos hidrofóbicos. Destes 50 aminoácidos extra,
20 funcionam como âncoras de membrana (Bertocci et al., 1991; Lundström et al.,
1991; Männistö e Kaakkola, 1999; Zhu, 2002). A maioria dos tecidos humanos
expressa os dois transcritos, no entanto, no cérebro, somente o maior transcrito pode
ser prontamente detectado. Como resultado, a MB-COMT é a forma enzimática
predominante no cérebro (70%), sendo a S-COMT encontrada em níveis reduzidos
(30%) (Tenhunen et al., 1994; Hong et al., 1998; Zhu, 2002; Chen et al., 2004).
Apesar da forma associada à membrana ser expressa em todas as regiões do sistema
nervoso central, dados mostram uma predominância no CPF (Hong et al., 1998;
Matsumoto et al., 2003).
Figura 2. O gene COMT e seus transcritos. O gene COMT é formado por seis éxons (representados pelas caixas cinza) e dois promotores, P2 e P1, os quais são responsáveis por controlar a expressão dos transcritos de 1.5 e 1.3 kb, respectivamente. O gene COMT contém alguns polimorfismos de interesse, como por exemplo, o rs2075507 e o rs4680. As posições de início da tradução da MB-COMT e S-MB-COMT, bem como o códon de terminação da tradução e a cauda poli-A (pA) estão indicados.
FONTE: adaptada de Zhu, 2002.
A COMT é importante para o catabolismo de uma gama de substratos catecol,
incluindo catecolaminas (dopamina, norepinefrina e epinefrina) e
substratos, por meio da utilização de uma molécula S-adenosil-L-metionina como
doador de grupo metil (Tunbridge et al., 2006).
No cérebro, a COMT pode modular as funções de neurotransmissão da dopmina
e norepinefrina em vários processos, através da alteração da taxa de inativação
metabólica destas substâncias (Zhu, 2002). A COMT é responsável por degradar
mais de 60% da dopamina presente nas fendas sinápticas do CPF (Karoum et al.,
1994). Em geral, o CPF expressa baixos níveis de proteínas transportadoras de
dopamina (DAT – dopamine transporter), sendo que dentro das fendas sinápticas
não é possível detectar sua expressão. Como o DAT é o mecanismo mais eficiente
para degradar a dopamina liberada no espaço extracelular, o CPF é dependente de
um mecanismo secundário, a COMT, para encerrar a ação da dopamina liberada
(Fig. 3) (Dauvilliers et al., 2014).
Figura 3. Transmissão dopaminérgica no córtex pré-frontal (CPF) e corpo estriado. No CPF a COMT é a principal responsável pela remoção da dopamina na fenda sináptica, enquanto em outras regiões cerebrais, como o corpo estriado, a proteína transportadora de dopamina (DAT - dopamine transporter) exerce esta função. FONTE: adaptada de Tunbridge et al., 2006.
Os níveis de dopamina nesta região cerebral são críticos para a modulação da
(Tunbridge et al., 2006; Kukshal et al., 2013). Fortes evidências sugerem que existe
uma relação em forma de U invertido entre atividade da dopamina no CPF e
desempenho da MT. Nesta relação, níveis intermediários de dopamina são
considerados ótimos para a performance da MT (Fig. 4) (Tunbridge et al., 2006).
1.3.2. Polimorfismos
Devido à importância da dopamina na regulação da cognição no CPF,
polimorfismos do gene COMT foram identificados como possíveis contribuintes
para a variação individual em funções dependentes nesta região. Evidências
sugerem que o gene COMT está associado com desempenho da MT (Goldberg et
al., 2003; Júlio-Costa et al., 2013).
Atualmente, existem vários SNPs descritos no gene COMT, como por exemplo:
rs2075507 localizado no promotor P2, rs6267 localizado no éxon 3, rs737865
localizado no íntron 1 e rs4680 localizado no éxon 4. Destes, o mais investigado na
literatura é o rs4680, também chamado de polimorfismo Val158Met (Fig. 2). Este
SNP, localizado na região codificadora do gene, consiste em uma substituição G → A, a qual por sua vez resulta na substituição de uma valina por uma metionina, na
posição 158 da MB-COMT (108 da S-COMT) (Bertocci et al., 1991; Lundström et
al., 1991; Swift-Scanlan et al., 2014).
Figura 4. Relação em forma de U invertido entre nível de dopamina no córtex pré-frontal e desempenho da memória de trabalho.
FONTE: adaptada de Tunbridge et al., 2006.
Em razão da presença deste SNP funcional, a COMT apresenta uma distribuição
(Floderus et al., 1981; Lachman et al., 1996). A forma Met apresenta menor
termoestabilidade e, portanto, menor atividade em temperatura fisiológica (Chen et
al., 2004). Assim, indivíduos homozigotos para o alelo valina exibem maior
atividade da COMT e menores concentrações de dopamina nas fendas sinápticas do
CPF. Já homozigotos para o alelo metionina exibem menor atividade da COMT
(taxa de degradação entre 1/3 e 1/4 mais lenta) e, consequentemente, maior
disponibilidade de dopamina nas fendas sinápticas. Como os alelos são
codominantes, heterozigotos apresentam níveis intermediários de atividade da
COMT, explicando assim a distribuição trimodal observada (Lachman et al., 1996;
Chen et al., 2004).
Figura 5. Efeito da atividade da COMT na relação em forma de U invertido entre nível de dopamina e função do córtex pré-frontal (CPF). Homozigotos Val estão associados a níveis subótimos de dopamina, enquanto homozigotos Met apresentam níveis próximos aos ideais.
FONTE: adaptada de Tunbridge et al., 2006.
Como mencionado anteriormente, em razão de o gene COMT ser importante
para a modulação dos níveis de dopamina no CPF, ele representa um excelente
candidato para modular a função desta região (Tunbridge et al., 2004). Egan et al.
(2001) demonstraram que o alelo Val esta associado com baixa performance em
tarefas dependentes do CPF, com homozigotos Met desempenhando melhor que
homozigotos Val, e heterozigotos apresentando desempenho intermediário. O
mesmo grupo de pesquisa constatou uma associação entre genótipo e atividade
pré-frontal. Utilizando ressonância magnética funcional durante tarefas que demandam
em relação a homozigotos Met, e que heterozigotos, mais uma vez, exibiram padrão
intermediário (Egan et al., 2001). A partir deste e de outros estudos, conclui-se que
indivíduos homozigotos para o alelo Val utilizam seus CPF de forma menos
eficiente que portadores do alelo Met, ou seja, indivíduos Val158Val requerem
maior ativação pré-frontal para um dado nível de desempenho. Portanto, a maior
atividade da enzima codificada a partir do alelo Val está relacionada à baixa
performance e à ativação pré-frontal menos eficiente, presumivelmente, devido aos
menores níveis de dopamina no CPF (Tunbridge et al., 2006). A baixa atividade da
enzima COMT, como resultado do alelo Met, leva a uma maior concentração de
dopamina pré-frontal, permitindo que ela se difunda mais através da fenda sináptica,
e assim, ative receptores extrassinápticos (dopamine receptor D1; ativação mais
eficiente). De maneira oposta, a alta atividade da COMT codificada a partir do alelo
Val leva a uma menor concentração de dopamina pré-frontal, o que limita a difusão
deste neurotransmissor e favorece a ativação de receptores intrassinápticos
(dopamine receptor D2; ativação menos eficiente; Fig. 3) (Winterer e Weinberger,
2004). Os receptores D1 estão relacionados à ativação mais eficiente do CPF devido
à habilidade de estimular um efeito excitatório global no interior da célula
(Kebabian e Calne, 1979).
Embora a maioria dos estudos de associação envolvendo a COMT esteja focada
no polimorfismo Val158Met, nos últimos anos, diversas evidências emergiram sugerindo que outros loci dentro do gene COMT podem constituir polimorfismos
capazes de influenciar na atividade da enzima. Sendo assim, alguns trabalhos têm utilizado haplótipos, uma vez que eles podem demonstrar efeitos mais confiáveis do
que o SNP Val158Met sozinho (Tunbridge et al., 2006). Por exemplo, o
polimorfismo A287G (rs2075507) localizado na região 5’ do gene (dentro do
promotor P2), possui um baixo efeito sobre a atividade da COMT e interage com o
polimorfismo Val158Met para modular a eficiência do CPF (Fig. 2) (Chen et al.,
2004; Meyer-Lindenberg et al., 2006). A presença do alelo G reduz os níveis de
atividade da COMT, comparado ao alelo A. Esta redução, em combinação com o
alelo Val, é benéfica para a função do CPF, uma vez que direciona a atividade da
COMT para próximo dos níveis ótimos. No entanto, esta redução é prejudicial
combinada com o alelo Met, dado que ela direciona a atividade da enzima para
níveis subótimos, refletindo assim em um aumento da dopamina pré-frontal acima
(2006) confirmaram estes achados por meio de estudos de neuroimagem durante
tarefas que demandam MT. A resposta pré-frontal mais eficiente foi observada para
o haplótipo MetA, enquanto a mais ineficiente foi observada para o haplótipo ValA.
As respostas para os haplótipos contendo o alelo G foram intermediárias.
1.3.3. Estudos de associação
Apesar do padrão mencionado acima ser bastante aceito (desempenho superior =
Met, desempenho inferior = Val), pesquisas mais recentes, considerando diferentes
grupos étnicos e faixas etárias têm gerado resultados discordantes.
Normalmente, os estudos realizados com adultos caucasianos confirmam que o
alelo Met está associado com melhor função cognitiva, especialmente em MT. No
entanto, diversos estudos com adultos chineses demonstraram que portadores do
alelo Val desempenharam melhor em tarefas que demandam MT que homozigotos
Met (Wang et al., 2013). Isto sugere que o efeito de diferentes haplótipos ou
backgrounds genéticos, modificam o impacto do polimorfismo Val158Met.
Figura 6. Efeito da atividade da COMT na relação em forma de U invertido entre nível de dopamina e função do córtex pré-frontal (CPF). Homozigotos Val podem ser deslocados para próximo do ápice da curva em razão da redução da atividade da COMT (presença do alelo G do SNP rs2075507). Em contrapartida, a redução da atividade da COMT em homozigotos Met resulta em níveis de dopamina exageradamente altos, prejudicando assim a função do CPF.
FONTE: adaptada de Tunbridge et al., 2006.
6 e 14 anos, identificaram que aquelas com genótipo Met158Met apresentaram
performance significativamente maior em tarefa que requer MT e inibição,
comparadas a crianças com genótipo Val158Val. As crianças heterozigotas exibiram
desempenho intermediário. Wahlstrom et al. (2007), em um estudo com crianças e
adolescentes, entre 9 e 17 anos, observaram que indivíduos heterozigotos e
homozigotos para o alelo Valdesempenharam melhor que homozigotos para o alelo
Met em tarefas de MT e atenção. Júlio-Costa e colaboradores (2013), a partir de
uma amostra de 155 crianças entre 8 e 12 anos, recrutadas a partir de escolas
públicas e privadas de Belo Horizonte, puderam observar que crianças com pelo
menos um alelo Met apresentaram desempenho superior em relação às homozigotas
Val158Val, em tarefas numéricas e aritméticas. No entanto, nenhuma diferença foi
detectada entre os genótipos em tarefas de MT.
Esta falta de concordância entre pesquisas com crianças e adolescentes pode ser
devido a diferentes efeitos dos genótipos da COMT em diferentes idades. Diversos
estudos realizados ao longo das últimas décadas demonstraram que a adolescência é
um período de contínuo desenvolvimento do CPF. A adolescência é caracterizada
pelo amadurecimento de uma variedade de processos mediados pelo CPF, como por
exemplo, a MT (De Luca et al., 2003; Luna et al., 2004; Luciana et al., 2005;
Conklin et al., 2007). A partir da puberdade até a vida adulta, os níveis de dopamina
decrescem no CPF (Wahlstrom et al., 2007). Este decréscimo nos níveis basais de dopamina entre a infância e a vida adulta pode afetar a posição dos genótipos
Val158Met na curva em forma de U invertido e, consequentemente, levar a diferentes efeitos do SNP rs4680 durante o desenvolvimento (Fig. 7) (Dumontheil et
al., 2011).
Durante a infância, devido à presença de maiores níveis de dopamina, a relação
entre os genótipos da COMT e o funcionamento cognitivo pode ser oposta a
observada em adultos. Os maiores níveis de dopamina podem interagir com o
genótipo Met158Met e causar níveis excessivos de dopamina no CPF, deslocando
os indivíduos portadores deste genótipo para uma posição mais a direita da curva em
U invertido. Posição esta caracterizada por funcionamento cognitivo ineficiente. Já
os indivíduos Val158Val e Val158Met se beneficiariam destes maiores níveis de
dopamina, uma vez que eles seriam deslocados para mais próximo do ápice da
curva. Este deslocamento resultaria em melhor funcionamento cognitivo comparado
Os resultados controversos entre diferentes estudos podem ser devido à ampla
gama de idades avaliadas. Muitos estudos investigando associação entre o
polimorfismo Val158Met e funções cognitivas fazem uso de amostras onde estão
presentes crianças e adultos (Wahlstrom et al., 2010). Além disso, mesmo em
estudos com amostras contendo apenas crianças, a diferença entre meninas e
meninos quanto à idade de início da puberdade, 10 e 12 anos, respectivamente, pode
levar a diferentes resultados.
Figura 7. Alterações na relação entre genótipos da COMT e desempenho cognitivo em função da idade. Na infância, devido à presença de maiores níveis de dopamina no córtex pré-frontal (CPF), homozigotos Val apresentam melhor desempenho cognitivo que homozigotos Met.
FONTE: adaptada de Wahlstrom et al., 2010.
1.4. Necessidades atuais
Muitos dos trabalhos prévios investigando os mecanismos subjacentes à MT e,
consequentemente, à aprendizagem, baseiam-se principalmente no estudo de
populações caucasianas adultas. Há uma carência em pesquisas, principalmente com
crianças, buscando averiguar polimorfismos distribuídos na população brasileira,
capazes de influenciar o amplo espectro fenotípico observado para a MT. Pesquisas
voltadas para a nossa população se fazem extremamente necessárias, uma vez que
nossa alta taxa de miscigenação pode acarretar em padrões de associação gene – MT
Compreender as modulações genéticas que regem as diferentes habilidades
cognitivas, como por exemplo, a MT, pode revelar passos essenciais para suplantar
a dificuldade de aprendizagem, principalmente entre crianças em idade escolar.
2. Justificativa e relevância
Aproximadamente 7% das crianças apresentam dificuldade matemática (DM),
enquanto 5 a 12% sofrem de dificuldade de leitura (DL) (De Weerdt et al., 2013).
Apesar de adequadas oportunidades de aprendizagem, normalmente, ambas as
dificuldades costumam persistir ou aumentar ao longo da vida escolar, o que pode
resultar em consequências negativas, tais como ansiedade, baixa motivação e alta
frustração acadêmica, bem como isolamento social (Schuchardt et al., 2008;
Willcutt et al., 2013).
Déficits na memória de trabalho (MT) são características comuns tanto da DL
quanto da DM. Diversas pesquisas demonstraram que crianças com DL exibem
danos nos componentes fonológico e executivo central da MT. Para a DM, déficits
em todos os três componentes da MT já foram descritos (De Weerdt et al., 2013).
Apesar da grande importância da MT, especialmente para o processo de
aprendizagem, pouco se conhece sobre suas bases genético-moleculares e,
consequentemente, como estas bases influenciam no fenótipo final em diferentes grupos étnicos e diferentes idades. Há uma carência em pesquisas buscando
investigar polimorfismos genéticos entre indivíduos brasileiros de desenvolvimento
típico, principalmente crianças, que possam estar associados ao grau de performance
da MT. Estudos voltados para a população brasileira se fazem necessários, pois a
alta taxa de miscigenação da mesma pode acarretar em padrões de associação
diferentes dos encontrados nas pesquisas com populações caucasianas. Diferenças
estas já comprovadas para as populações asiáticas.
Sendo assim, este presente trabalho objetiva identificar as contribuições de
polimorfismos da COMT (SNPs rs4680 e rs2075507) para o desempenho da MT,
em crianças provenientes de escolas públicas e privadas de Belo Horizonte. Além de
dar continuidade a linha de pesquisa desenvolvida pelo Laboratório de Genética
Humana e Médica em parceria com o Laboratório de Neuropsicologia do
Desenvolvimento, ambos da UFMG, busca-se aprofundar os resultados encontrados
Analisando o contexto atual, este trabalho é inovador, pois procura compreender
a influência de alterações genéticas comuns e, portanto, de seus mecanismos
moleculares subjacentes, na performance de MT, em crianças da população
brasileira. Uma vez que a MT é uma medida relativamente pura da capacidade de
aprendizagem, os resultados deste trabalho poderão auxiliar no desenvolvimento de
estratégias educacionais compensatórias eficazes para o combate das dificuldades de
leitura e Matemática nas escolas.
Referências Bibliográficas
ALLOWAY, T. P. Working memory, but not IQ, predicts subsequent learning in
children with learning difficulties. European J of Psychological
Assessment, v. 25, p. 92–98, 2009.
ALLOWAY, T. P.; GATHERCOLE, S. E.; KIRKWOOD, H.; ELLIOTT, J. The
cognitive and behavioral characteristics of children with low working
memory. Child Dev, v. 80, n. 2, p. 606-21, 2009 Mar-Apr 2009.
ALLOWAY, T. P.; GATHERCOLE, S. E.; WILLIS, C.; ADAMS, A. M.
Working memory and special educational needs. Edu and Child
Psychol, v. 22, p. 56–67, 2005.
ANDO, J.; ONO, Y.; WRIGHT, M. J. Genetic structure of spatial and verbal
working memory. Behav Genet, v. 31, n. 6, p. 615-24, Nov 2001.
BADDELEY, A. The episodic buffer: a new component of working memory?
Trends Cogn Sci, v. 4, n. 11, p. 417-423, Nov 1 2000.
BADDELEY, A. Working memory and language: an overview. J Commun
Disord, v. 36, n. 3, p. 189-208, 2003 May-Jun 2003a.
BADDELEY, A. Working memory: looking back and looking forward. Nat Rev
Neurosci, v. 4, n. 10, p. 829-39, Oct 2003b.
BADDELEY, A. D.; HITCH, G. J. Working memory. Recent advances in
learning and motivation, v. 8, n. New York: Academic Press, p. 44 - 90,
1974.
BAILEY, D. B.; HATTON, D. D.; SKINNER, M. Early developmental
trajectories of males with fragile X syndrome. Am J Ment Retard, v.
BERTOCCI, B.; MIGGIANO, V.; DA PRADA, M.; DEMBIC, Z.; LAHM, H.
W.; MALHERBE, P. Human catechol-O-methyltransferase: cloning
and expression of the membrane-associated form. Proc Natl Acad Sci U
S A, v. 88, n. 4, p. 1416-20, Feb 1991.
BRAY, S.; DUNKIN, B.; HONG, D. S.; REISS, A. L. Reduced functional
connectivity during working memory in Turner syndrome. Cereb
Cortex, v. 21, n. 11, p. 2471-81, Nov 2011.
CASE, R.; KURLAND, D. M.; GOLDBERG, J. Operational efficiency and the
growth of short-term memory span. J Experimental Child Psychol, v.
33, p. 386-404, 1982.
CHEN, J.; LIPSKA, B. K.; HALIM, N.; MA, Q. D.; MATSUMOTO, M.;
MELHEM, S.; KOLACHANA, B. S.; HYDE, T. M.; HERMAN, M.
M.; APUD, J.; EGAN, M. F.; KLEINMAN, J. E.; WEINBERGER, D.
R. Functional analysis of genetic variation in
catechol-O-methyltransferase (COMT): effects on mRNA, protein, and enzyme
activity in postmortem human brain. Am J Hum Genet, v. 75, n. 5, p.
807-21, Nov 2004.
CHEN, L. S.; RICE, T. K.; THOMPSON, P. A.; BARCH, D. M.;
CSERNANSKY, J. G. Familial aggregation of clinical and
neurocognitive features in sibling pairs with and without schizophrenia. Schizophr Res, v. 111, n. 1-3, p. 159-66, Jun 2009.
CONKLIN, H. M.; LUCIANA, M.; HOOPER, C. J.; YARGER, R. S. Working memory performance in typically developing children and adolescents:
behavioral evidence of protracted frontal lobe development. Dev
Neuropsychol, v. 31, n. 1, p. 103-28, 2007.
CONWAY, A. R.; KANE, M. J.; BUNTING, M. F.; HAMBRICK, D. Z.;
WILHELM, O.; ENGLE, R. W. Working memory span tasks: A
methodological review and user's guide. Psychon Bull Rev, v. 12, n. 5,
p. 769-86, Oct 2005.
CORSO, L. V.; DORNELES, B. V. What Role does the Working Memory Play
in Mathematics Learning? Bolema: Boletim de Educação Matemática,
DAUVILLIERS, Y.; TAFTI, M.; LANDOLT, H. P.
Catechol-O-methyltransferase, dopamine, and sleep-wake regulation. Sleep Med
Rev, Oct 2014.
DE LUCA, C. R.; WOOD, S. J.; ANDERSON, V.; BUCHANAN, J. A.;
PROFFITT, T. M.; MAHONY, K.; PANTELIS, C. Normative data
from the CANTAB. I: development of executive function over the
lifespan. J Clin Exp Neuropsychol, v. 25, n. 2, p. 242-54, Apr 2003.
DE SMEDT, B.; DEVRIENDT, K.; FRYNS, J. P.; VOGELS, A.; GEWILLIG,
M.; SWILLEN, A. Intellectual abilities in a large sample of children
with Velo-Cardio-Facial Syndrome: an update. J Intellect Disabil Res,
v. 51, n. Pt 9, p. 666-70, Sep 2007.
DE SMEDT, B.; SWILLEN, A.; VERSCHAFFEL, L.; GHESQUIÈRE, P.
Mathematical learning disabilities in children with 22q11.2 deletion
syndrome: a review. Dev Disabil Res Rev, v. 15, n. 1, p. 4-10, 2009.
DE WEERDT, F.; DESOETE, A.; ROEYERS, H. Working memory in children
with reading disabilities and/or mathematical disabilities. J Learn
Disabil, v. 46, n. 5, p. 461-72, 2013 Sep-Oct 2013.
DIAMOND, A.; BRIAND, L.; FOSSELLA, J.; GEHLBACH, L. Genetic and
neurochemical modulation of prefrontal cognitive functions in children.
Am J Psychiatry, v. 161, n. 1, p. 125-32, Jan 2004.
DUMONTHEIL, I.; ROGGEMAN, C.; ZIERMANS, T.; PEYRARD-JANVID,
M.; MATSSON, H.; KERE, J.; KLINGBERG, T. Influence of the COMT genotype on working memory and brain activity changes during
development. Biol Psychiatry, v. 70, n. 3, p. 222-9, Aug 2011.
EGAN, M. F.; GOLDBERG, T. E.; KOLACHANA, B. S.; CALLICOTT, J. H.;
MAZZANTI, C. M.; STRAUB, R. E.; GOLDMAN, D.;
WEINBERGER, D. R. Effect of COMT Val108/158 Met genotype on
frontal lobe function and risk for schizophrenia. Proc Natl Acad Sci U S
A, v. 98, n. 12, p. 6917-22, Jun 2001.
FIGUEIREDO, V. L. M. WISC-III: Escala de Inteligência Wechsler para
Crianças. Manual, Adaptação e Padronização Brasileira. São Paulo:
FLODERUS, Y.; ROSS, S. B.; WETTERBERG, L. Erythrocyte
catechol-O-methyltransferase activity in a Swedish population. Clin Genet, v. 19, n.
5, p. 389-92, May 1981.
GOLDBERG, T. E.; EGAN, M. F.; GSCHEIDLE, T.; COPPOLA, R.;
WEICKERT, T.; KOLACHANA, B. S.; GOLDMAN, D.;
WEINBERGER, D. R. Executive subprocesses in working memory:
relationship to catechol-O-methyltransferase Val158Met genotype and
schizophrenia. Arch Gen Psychiatry, v. 60, n. 9, p. 889-96, Sep 2003.
HART, S. J.; DAVENPORT, M. L.; HOOPER, S. R.; BELGER, A. Visuospatial
executive function in Turner syndrome: functional MRI and
neurocognitive findings. Brain, v. 129, n. Pt 5, p. 1125-36, May 2006.
HONG, J.; SHU-LEONG, H.; TAO, X.; LAP-PING, Y. Distribution of
catechol-O-methyltransferase expression in human central nervous system.
Neuroreport, v. 9, n. 12, p. 2861-4, Aug 1998.
JÚLIO-COSTA, A.; ANTUNES, A. M.; LOPES-SILVA, J. B.; MOREIRA, B.
C.; VIANNA, G. S.; WOOD, G.; CARVALHO, M. R.; HAASE, V. G.
Count on dopamine: influences of COMT polymorphisms on numerical
cognition. Front Psychol, v. 4, p. 531, 2013.
KARLSGODT, K. H.; BACHMAN, P.; WINKLER, A. M.; BEARDEN, C. E.;
GLAHN, D. C. Genetic influence on the working memory circuitry: behavior, structure, function and extensions to illness. Behav Brain Res,
v. 225, n. 2, p. 610-22, Dec 2011.
KARLSGODT, K. H.; KOCHUNOV, P.; WINKLER, A. M.; LAIRD, A. R.;
ALMASY, L.; DUGGIRALA, R.; OLVERA, R. L.; FOX, P. T.;
BLANGERO, J.; GLAHN, D. C. A multimodal assessment of the
genetic control over working memory. J Neurosci, v. 30, n. 24, p.
8197-202, Jun 2010.
KAROUM, F.; CHRAPUSTA, S. J.; EGAN, M. F. 3-Methoxytryramine is the
major metabolite of released dopamine in the rat frontal
cortex:reassessment of the effects of antipsychotics on the dynamics of
dopamine release and metabolism in the frontal cortex, nucleus
accumbens, and striatum bya simple two pool model. J. Neurochem, v.
KEBABIAN, J. W.; CALNE, D. B. Multiple receptors for dopamine. Nature, v.
277, n. 5692, p. 93-6, Jan 1979.
KESSELS, R. P.; VAN ZANDVOORT, M. J.; POSTMA, A.; KAPPELLE, L.
J.; DE HAAN, E. H. The Corsi Block-Tapping Task: standardization
and normative data. Appl Neuropsychol, v. 7, n. 4, p. 252-8, 2000.
KUKSHAL, P.; KODAVALI, V. C.; SRIVASTAVA, V.; WOOD, J.;
MCCLAIN, L.; BHATIA, T.; BHAGWAT, A. M.; DESHPANDE, S.
N.; NIMGAONKAR, V. L.; THELMA, B. K. Dopaminergic gene
polymorphisms and cognitive function in a north Indian schizophrenia
cohort. J Psychiatr Res, v. 47, n. 11, p. 1615-22, Nov 2013.
LACHMAN, H. M.; PAPOLOS, D. F.; SAITO, T.; YU, Y. M.; SZUMLANSKI,
C. L.; WEINSHILBOUM, R. M. Human catechol-O-methyltransferase
pharmacogenetics: description of a functional polymorphism and its
potential application to neuropsychiatric disorders. Pharmacogenetics,
v. 6, n. 3, p. 243-50, 1996.
LEE, T.; MOSING, M. A.; HENRY, J. D.; TROLLOR, J. N.; AMES, D.;
MARTIN, N. G.; WRIGHT, M. J.; SACHDEV, P. S.; TEAM, O. R.
Genetic influences on four measures of executive functions and their
covariation with general cognitive ability: the Older Australian Twins
Study. Behav Genet, v. 42, n. 4, p. 528-38, Jul 2012.
LINDSAY, E. A. Chromosomal microdeletions: dissecting del22q11 syndrome.
Nat Rev Genet, v. 2, n. 11, p. 858-68, Nov 2001.
LUCIANA, M.; CONKLIN, H. M.; HOOPER, C. J.; YARGER, R. S. The
development of nonverbal working memory and executive control
processes in adolescents. Child Dev, v. 76, n. 3, p. 697-712, 2005
May-Jun 2005.
LUNA, B.; GARVER, K. E.; URBAN, T. A.; LAZAR, N. A.; SWEENEY, J. A.
Maturation of cognitive processes from late childhood to adulthood.
Child Dev, v. 75, n. 5, p. 1357-72, 2004 Sep-Oct 2004.
LUNDSTRÖM, K.; SALMINEN, M.; JALANKO, A.; SAVOLAINEN, R.;
ULMANEN, I. Cloning and characterization of human placental
catechol-O-methyltransferase cDNA. DNA Cell Biol, v. 10, n. 3, p.
MÄNNISTÖ, P. T.; KAAKKOLA, S. Catechol-O-methyltransferase (COMT):
biochemistry, molecular biology, pharmacology, and clinical efficacy
of the new selective COMT inhibitors. Pharmacol Rev, v. 51, n. 4, p.
593-628, Dec 1999.
MATSUMOTO, M.; WEICKERT, C. S.; AKIL, M.; LIPSKA, B. K.; HYDE, T.
M.; HERMAN, M. M.; KLEINMAN, J. E.; WEINBERGER, D. R.
Catechol O-methyltransferase mRNA expression in human and rat
brain: evidence for a role in cortical neuronal function. Neuroscience, v.
116, n. 1, p. 127-37, 2003.
MAYNARD, T. M.; HASKELL, G. T.; PETERS, A. Z.; SIKICH, L.;
LIEBERMAN, J. A.; LAMANTIA, A. S. A comprehensive analysis of
22q11 gene expression in the developing and adult brain. Proc Natl
Acad Sci U S A, v. 100, n. 24, p. 14433-8, Nov 2003.
MAZZOCCO, M. M. Math learning disability and math LD subtypes: evidence
from studies of Turner syndrome, fragile X syndrome, and
neurofibromatosis type 1. J Learn Disabil, v. 34, n. 6, p. 520-33, 2001
Nov-Dec 2001.
MAZZOCCO, M. M. Mathematical learning disability in girls with Turner
syndrome: a challenge to defining MLD and its subtypes. Dev Disabil
Res Rev, v. 15, n. 1, p. 35-44, 2009.
MEYER-LINDENBERG, A.; NICHOLS, T.; CALLICOTT, J. H.; DING, J.;
KOLACHANA, B.; BUCKHOLTZ, J.; MATTAY, V. S.; EGAN, M.; WEINBERGER, D. R. Impact of complex genetic variation in COMT
on human brain function. Mol Psychiatry, v. 11, n. 9, p. 867-77, 797,
Sep 2006.
MONTOJO, C. A.; IBRAHIM, A.; KARLSGODT, K. H.; CHOW, C.;
HILTON, A. E.; JONAS, R. K.; VESAGAS, T. K.; BEARDEN, C. E.
Disrupted working memory circuitry and psychotic symptoms in
22q11.2 deletion syndrome. Neuroimage Clin, v. 4, p. 392-402, 2014.
MURPHY, M. M. A review of mathematical learning disabilities in children
with fragile X syndrome. Dev Disabil Res Rev, v. 15, n. 1, p. 21-7,