Functional analysis of Rift Valley fever virus NSs encoding a partial truncation.
Texto
Imagem
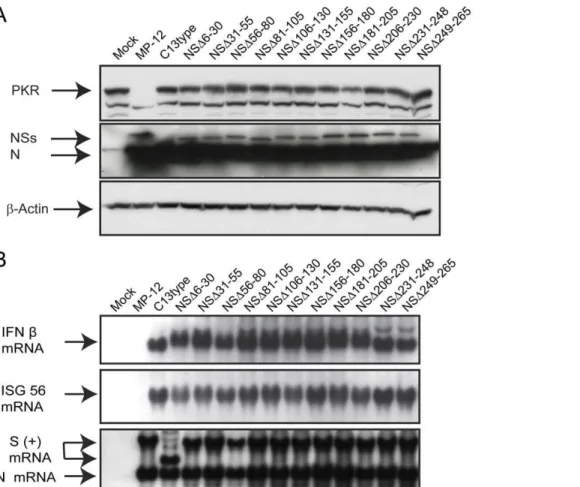

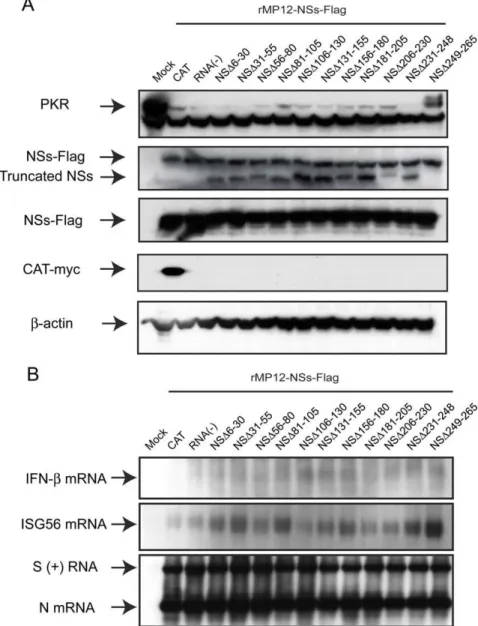
Documentos relacionados
(H) Huh7.5.1 cells were transfected with MEK1 siRNA1, MEK1 siRNA2 or scramble RNA, 6 hr later, cells were infected with JFH-1 (MOI 0.5), 3 days post infection, cells were
HekT cells were transiently transfected with plasmids encoding the indicated V5-tagged UbcH proteins together with either empty vector or FLAG-tagged Hu, Agm or Mo ISG15 or a
HeLa cells were left untreated (Medium, A) or transiently transfected with control RNAi (B) or CK18 RNAi (C).. After 24 h, cells were infected with tis- sue culture trypomastigotes
C and D HEK293T cells were transfected with the indicated HA-tagged Akt1 phosphorylation site mutants with or without co- transfected FLAG-tagged FKBP51 K352A/R356A (TPR_mut).. After
(C) HeLa cells transfected with either non-targeting siRNA or siRNA targeting CPSF6 were infected with VSV-G-pseudotyped GFP carrying CA mutants that lost the cell cycle independence
Reporter cells were cotransformed with a prey vector encoding SEC15 and a bait plasmid encoding either wild type or GTPase deficient mutant Sec4p Q79L with phosphorylated
siRNAs directed against (A) luciferase or XIAP, (B) cIAP-1, (C) cIAP-2, or (D) survivin were transfected in HeLa cells, and the protein levels of all IAPs were determined in infected
(B and C) HEK-293T cells were transfected with expression vectors encoding GST alone or fused to the C proteins of measles virus (MV-C), hPIV3 (hPIV3-C), or Nipah virus (Nipah-C),
