Ciências Veterinárias
Estudo fenotípico da microbiota bacteriana
do trato respiratório superior do
golfinho-roaz (Tursiops truncatus)
Raquel Maria Pereira Fernandes
Orientadora:
Professora Doutora Ana Cláudia Correia Coelho Co-Orientadora:
Doutora Carla Anne Flanagan
UNIVERSIDADE DE TRÁS-OS-MONTES E ALTO DOURO VILA REAL, 2012
Estudo fenotípico da microbiota bacteriana
do trato respiratório superior do
golfinho-roaz (Tursiops truncatus)
Raquel Maria Pereira Fernandes
Orientadora:
Professora Doutora Ana Cláudia Correia Coelho Co-Orientadora:
Doutora Carla Anne Flanagan
UNIVERSIDADE DE TRÁS-OS-MONTES E ALTO DOURO VILA REAL, 2012
Challenges are what make life interesting; overcoming them is what makes life meaningful. Joshua J. Marine
IV
Resumo
A microbiota é o termo que inclui bactérias, vírus, protozoários e fungos que habitam em diferentes regiões anatómicas. Estes microrganismos estão adaptados para viver dentro do hospedeiro sem causar doenças ou danos. A interação entre o hospedeiro e o meio ambiente estabelece a microbiota normal, sendo esta determinada por muitos fatores, tais como, a idade, raça, níveis hormonais, dieta, stress, comportamento sexual, medicação (caso esteja a ser feita), estação do ano, localização geográfica, habitat, densidade populacional, contato entre animais e, até, procedimentos de limpeza do habitat.
Devido à crescente importância das doenças respiratórias em cetáceos este trabalho pretende obter resultados que permitam caracterizar a microbiota do trato respiratório superior de vários exemplares da espécie Tursiops truncatus (Golfinho-roaz) que vivem sob cuidados humanos no Zoomarine, bem como comparar esta com estudos anteriores realizados pelo Zoomarine com dados recolhidos entre o ano de 2005 e o de 2011.
Para tal, realizaram-se colheitas assépticas de material do espiráculo destes animais e, posteriormente, o seu processamento laboratorial.
Os resultados mostraram uma microbiota variada: na primeira colheita, a bactéria mais isolada foi Escherichia coli, seguido de Morganella morganii, Enterococcus spp. e Pseudomonas aeruginosa, Staphylococcus spp. e, com menor frequência, Streptococcus spp. e Flavobacterium spp.. Na segunda colheita, o género de bactérias mais isolado foi Enterococcus spp., seguido de Morganella morganii e Escherichia coli, Staphylococcus spp. e Aeromonas salmonicida. Além disso, verificou-se a presença de Leveduras, nomeadamente Candida albicans em vários animais.
Sendo a pneumonia bacteriana uma das principais causas de morte nestes animais torna-se urgente e essencial andar um passo à frente da doença, de modo a que esta possa ser detetada de forma precoce. Como tal, torna-se fundamental conhecer a sua microbiota respiratória normal.
Palavras-chave: golfinho-roaz; Tursiops truncatus; mamíferos marinhos; microbiota
V
Abstract
The microbiota is the term that includes bacteria, viruses, protozoa and fungi that live in different anatomical regions. These microorganisms are adapted to live within the host without causing disease or injury. The interaction between the host and the environment establishes the normal microbiota, which is determined by many factors, such as age, race, hormone levels, diet, stress, sexual behavior, medication (if that is the case), season of the year, geographical location, habitat, population density, contact between animals and even the cleaning procedures of the habitat.
Due to the great importance of respiratory diseases in cetaceans, the aim of this investigation was to study the microbiota of the respiratory tract of several specimens of Tursiops truncatus (bottlenose dolphin) that live under human care in Zoomarine, and compare these with previous studies performed by Zoomarine with data collected between 2005 and 2011.
To perform the study, samples were collected from the blowhole of these animals and were subsequently processed in laboratory.
The results showed a varied flora: in the first collection, the most commonly isolated bacteria was Escherichia coli, followed by Morganella morganii, Enterococcus spp. and Pseudomonas aeruginosa, Staphylococcus spp. and, less frequently, Streptococcus spp. and Flavobacterium spp.. In the second collection, the most isolated bacteria was Enterococcus spp., followed by Morganella morganii and Escherichia coli, Staphylococcus spp. and Aeromonas salmonicida. Moreover, we verified the presence of yeasts, particularly Candida albicans in various animals.
Pneumonia due to bacteria is one of the leading causes of death in these animals, so it is essential to be one step ahead of the disease so that it can be detected early. Therefore, it is fundamental to know the normal respiratory microbiota.
Keywords: bottlenose dolphin; Tursiops truncatus; marine mammals; respiratory
VI
Índice
1 Introdução ... 1
2 Revisão bibliográfica ... 2
2.1 Golfinho-roaz ... 2
2.1.1 Características gerais e classificação ... 2
2.1.2 Biologia e fisiologia reprodutiva ... 3
2.1.3 Dieta ... 4
2.1.4 Suplementação vitamínica ... 5
2.2 Sistema respiratório dos mamíferos marinhos ... 6
2.2.1 Adaptações para uma vida marinha ... 6
2.2.2 Pulmões, pleura e linfonodos relacionados nos cetáceos ... 8
2.2.3 Doenças do trato respiratório dos cetáceos: a importância relacionada da recolha de amostras do espiráculo ... 9
2.3 Bactérias da microbiota do trato respiratório superior ... 10
2.3.1 Streptococcus spp. ... 10 2.3.2 Aeromonas salmonicida ... 11 2.3.3 Staphylococcus spp. ... 12 2.3.4 Enterococcus spp. ... 13 2.3.5 Flavobacterium spp. ... 14 2.3.6 Morganella morganii... 15 2.3.7 Escherichia coli ... 16 2.3.8 Pseudomonas aeruginosa... 17 3 Objetivos ... 18 4 Material e métodos ... 19
4.1 Isolamento de bactérias provenientes da microbiota do trato respiratório superior do golfinho-roaz ... 19
4.1.1 Origem das amostras ... 19
4.1.2 Material utilizado e colheita das amostras ... 20
4.1.3 Isolamento e identificação das bactérias ... 22
VII
4.1.5 Métodos morfofisiológicos para identificação dos isolados... 22
4.1.6 Conservação dos isolados ... 23
4.1.7 Análises das amostras de água realizadas por rotina no Zoomarine ... 23
4.1.8 Colheita das amostras de água ... 23
4.1.9 Análise das amostras de água ... 24
4.1.10 Galerias API® (Biomérieux) ... 25
4.2 Parâmetros físico-químicos dos habitats dos animais do estudo ... 25
5 Resultados ... 26
5.1 Caracterização da população ... 26
5.2 Caracterização da microbiota respiratória da 1ª recolha ... 26
5.3 Caracterização da microbiota respiratória da 2ª recolha ... 27
5.4 Identificação individual da microbiota respiratória ... 28
5.5 Evolução de isolamentos bacterianos no Zoomarine entre 2005 e 2011 ... 29
5.6 Caracterização microbiana das águas dos habitats ... 32
6 Discussão ... 34 7 Considerações finais ... 38 8 Bibliografia ... 39 9 Anexos ... 45 9.1 Anexo I ... 45 Meios seletivos ... 45
Meio de MacConkey® (Liofilchem 610028) ... 45
Meio de GSP® (Merck 15727) ... 45
Meio de Chapman® (Merck 1.05469.0500) ... 46
Meio de Cromocult® (Merck 1.10426) ... 46
Meio de Enterococcosel® (BD 212205) ... 46
Meio de CNA® (BD 212104) ... 47
Caldo de BHI® (Himedia M210I) com 6,5%de NaCl ... 47
Meio de BHI® (Oxoid CM225) com sangue ... 47
VIII
9.2 Anexo II ... 48
Métodos morfofisiológicos para identificação dos isolados ... 48
Coloração de Gram ... 48
Prova da citocromo-oxidase ... 49
Prova da catalase ... 49
Prova da coagulase ... 50
Prova do hidróxido de potássio a 3% (KOH) ... 50
Inoculação no meio de TSI® (Triple Sugar Iron) (BD 226540) ... 51
Provas combinadas ... 51
9.3 Anexo III ... 54
Atividades desenvolvidas durante o estágio decorrente deste trabalho de investigação ... 54 9.4 Anexo IV ... 57 O Zoomarine ... 57 9.5 Anexo V ... 59 Meios de cultura ... 59 9.6 Anexo VI ... 64
IX
Índice de figuras
Figura 1- Golfinho-roaz. ... 2 Figura 2- Golfinho-roaz (barbatanas peitorais). ... 2 Figura 3- Principais características anatómicas do Golfinho-roaz. ... 3 Figura 4- Pormenor das diferenças ao nível dos genitais do golfinho-roaz fêmea e macho. ... 3
Figura 5- Representação esquemática das vias respiratória e digestiva de um golfinho-roaz. ... 6
Figura 6- Anatomia comparada das vias respiratória e digestiva de um mamífero terrestre e um mamífero marinho (golfinho-roaz). ... 7
Figura 7- Candida spp. numa preparação de azul de metileno (40x). ... 9 Figura 8- Pormenor de uma célula epitelial, leucócitos e muco numa prepraração de azul de metileno (40x)... 9 Figura 9- Pormenor de uma célula epitelial com várias bactérias numa preparação de azul de metileno (40x). ... 9
Figura 10- Pormenor de estruturas fúngicas numa coloração de Gram feita a partir de uma amostra do espiráculo. ... 10 Figura 11- Material necessário para a realização de uma citologia com amostras do espiráculo. ... 20
Figura 12- Representação esquemática da metodologia de colheita da amostra. .. 21 Figura 13- Copos esterilizados durante a filtração das amostras de água na câmara de fluxo laminar ... 24 Figura 14- Pormenor do filtro® (Whatman) a ser retirado de um dos copos esterilizados após a filtração. ... 24 Figura 15- Depois de retirar do copo esterilizado o filtro® (Whatman) é colocado numa placa com um dos meios de cultura. ultilizados ... 25 Figura 16- A placa é posteriormente fechada ainda na câmara de fluxo laminar e levada a incubar na estufa. ... 25 Figura 17- Percentagem de bactérias Gram Negativas (57%) e Gram Positivas (43%) encontradas na primeira colheita. ... 27
Figura 18- Percentagem de bactérias Gram Negativas (60%) e Gram Positivas (40%) encontradas na segunda colheita. ... 28
Figura 19- Placa com meio de MacConkey® (Liofilchem 610028) inoculado com uma bactéria fermentadora de lactose. ... 45
Figura 20- Comparação de uma placa com meio de GSP® (Merck 15727) não inoculado com duas placas do mesmo meio com crescimento de Aeromonas. ... 45
X Figura 21- Pormenor de uma colónia isolada a crescer no meio de GSP® (Merck
15727). ... 45
Figura 22- Três placas de GSP® (Merck 15727) com crescimento de Aeromonas e alteração da cor do meio. ... 45
Figura 23- Placa com meio de Chapman® (Merck 1. 05469. 0500) com crescimento de Staphylococcus. ... 46
Figura 24- Placa com meio de Cromocult® (Merck 1.10426) com crescimento de E. coli em colónias de cor azul. ... 46
Figura 25- Placa com meio de Enterococcosel® (BD 212205) com crescimento de Enterococcus e alteração da cor do meio. ... 46
Figura 26- Placas com meio de Sabouraud® (Liofilchem 610103) com crescimento de leveduras. ... 47
Figura 27- Pormenor das colónias de leveduras... 47
Figura 28- Prova da oxidase- a tira de papel de filtro superior demonstra uma reação negativa e a tira de papel inferior demonstra uma reação positiva. ... 49
Figura 29- Prova da catalase- Ambas gotas demonstram uma reação positiva. ... 49
Figura 30- Prova da coagulase- o tubo da esquerda demonstra uma reação negativa e o da direita uma reação positiva. ... 50
Figura 31- Meio de TSI® (Triple Sugar Iron) (BD 226540) inoculado... 51
Figura 32- Presença de um halo de indol. ... 52
Figura 33- Tubos com estirpes positivas à prova do Vermelho de Metilo. ... 52
Figura 34- Leão-marinho-californiano. ... 57
Figura 35- Foca-cinzenta. ... 57
Figura 36- Foca-vitulina. ... 57
Figura 37- Otária-Sul-Africana (fêmea e macho). ... 58
Figura 38- Golfinho-roaz (2). ... 58
Índice de Tabelas Tabela 1- Espécies de peixes e cefalópode consumidos pelos animais do parque. . 5
Tabela 2- Parâmetros físico-químicos dos habitats dos animais do estudo (T- temperatura e HR- humidade relativa). ... 25
Tabela 3- Características dos animais em estudo. ... 26
Tabela 4- Isolamento bacteriano obtido no estudo (1ª colheita). ... 27
Tabela 5- Isolamento bacteriano obtido no estudo (2ª colheita). ... 28
Tabela 6- Identificação da microbiota de cada animal do estudo. ... 29
Tabela 7- Evolução das bactérias isoladas ao longo do tempo, utilizando registos desde 2005 (fonte: Zoomarine). ... 30
XI Tabela 8- Bactérias previamente isoladas nos animais do estudo com base em registos desde 2005 (fonte: Zoomarine). ... 31
Tabela 9- Antibioterapia utilizada nos animais do estudo. ... 31 Tabela 10- Bactérias isoladas noutros golfinhos-roazes do parque que não os incluídos no estudo. ... 32 Tabela 11- Caracterização microbiana das águas dos habitats 1 e 2 na 1ª colheita1. ... 32
Tabela 12- Caracterização microbiana das águas dos habitats 1 e 3 na 2ª colheita2. ... 33
XII
Lista de siglas e abreviaturas
®- Marca registada ºC- Graus Celsius µm- Micrómetro º- Graus α- Alfa β- Beta γ- Gama %- Por cento
BHI- Brain Heart Infusion BID- Duas vezes ao dia CNA- Agar Colistina Nalidíxico DNA- Ácido Desoxirribonucleico Ex- Exemplo
GSP- Glutamato Amido Vermelho de Fenol HR- Humidade relativa H2S- Sulfureto de hidrogénio KOH-Hidróxido de potássio mg- Miligrama mL- Mililítro MR- Vermelho de Metilo m3- Metros cúbicos NaCl- Cloreto de sódio PCA- Plate Count Agar
PCR- Polymerase Chain Reaction PO- Por via oral
SA-Sociedade Anónima T- Temperatura
TID- Três vezes ao dia TSI- Triple Sugar Iron
UTAD- Universidade de Trás-os-Montes e Alto Douro VP- Voges Proskauer
XIII
Agradecimentos
Nesta fase de conclusão do Mestrado Integrado em Medicina Veterinária, gostaria de agradecer a todos que de alguma forma estiveram presentes ao longo deste percurso.
Ao Reitor da Universidade de Trás-os-Montes e Alto Douro, Professor Doutor Carlos Alberto Sequeira, e à instituição que representa, pela disponibilidade e facilidades concedidas.
À minha orientadora, Professora Ana Cláudia Coelho, pelo auxílio, incansável paciência e gosto que me incutiu pela Microbiologia.
À minha co-orientadora, Doutora Carla Flanagan, pelo acompanhamento e preocupação, pela efetiva orientação na prática clínica de animais exóticos e mamíferos marinhos, pelo incentivo constante, por estimular a perseguição das minhas ambições. Por acreditar nas minhas capacidades.
Aos Professores que, de alguma forma, ajudaram e incentivaram na perseguição do meu objetivo, nomeadamente, Professor Celso Santos, Professora Cristina Saraiva, Professor Luís Cardoso, Professora Madalena Vieira-Pinto, Professor Francisco Brito.
Às técnicas do Laboratório de Microbiologia Médica da UTAD, Fátima Fraga, Sónia Dias e Anabela Gonçalves, pela enorme paciência, ajuda e todo o trabalho que tiveram comigo.
Aos amigos que me acompanharam nesta jornada fantástica que é a vida académica, todos os finalistas do ano 2010/2011. Os momentos que passamos juntos foram inesquecíveis e estarão sempre no meu coração. À Filipa Mota, pela amizade, boas conversas. Ainda custa a acreditar que já não estás aqui connosco.
À Família Araucária, por me ter adotado, e a todos os seus membros: Aldo Andrade, Cláudia Soares, Daniela Cardoso, Hélia Ferreira, Inês Ferreira, Isabel Sanchez, Joana Campos, Joana Cardoso, Joana Grácio, Vanessa Medeiros. Obrigada pela espetacular vida académica (e não só), por cada um dos momentos que tive a honra de partilhar convosco.
Aos meus padrinhos de curso, Carla Teixeira e Luís Ferreira, por tudo o que fizeram por mim e por estarem sempre lá.
Aos meus afilhados de curso, Tiago Almeida e Artur Nascimento, vocês são o meu orgulho. Aos meus afilhados académicos, João Tomás, Jorge Silva, Maria João Rebelo, Silvana Machado, Carla Costa, Marlene Santos. Foi uma honra praxar-vos e poder estar lá para vos ensinar, apoiar, incutir o amor pelo curso e gritar sempre por ele, até não ter voz.
À fantástica equipa Veterinária do Zoomarine Algarve, Doutora Carla Flanagan (mais uma vez), Doutor Nuno Urbani e Enfermeira Veterinária Joana Silva, por todos os
XIV ensinamentos, oportunidades, carinho, companheirismo. O tempo que passei com vocês não podia ter sido melhor, obrigada por serem os meus maiores exemplos profissionais e, além disso, meus amigos. À Enfermeira Veterinária Antonieta Nunes, por ser o meu Porto d’Abrigo. És muito mais do que uma colega de trabalho, estarás sempre no meu coração. Ao Mestre Miguel Silveira pelo apoio constante e pelo interesse no meu trabalho. À Mia Coelho pela ajuda e pela paciência, por ter sempre respondido aos infindáveis e-mails, por acreditar em mim. À Isabel Gaspar pelos ensinamentos de uma área tão vasta como a dos peixes, pela amizade e pelas conversas. Ao Marco Bragança pela boa disposição e conhecimentos partilhados. À Letizia Fiorucci, com quem construí uma amizade fantástica que nunca vou esquecer, pelas conversas de muitas horas, pela troca de ideias e pela discussão de casos. Grazie sorellina, non ti dimenticherò.
Ao Doutor Luís Roque, pelos ensinamentos valiosíssimos e pelas lembranças do Norte, em particular de Vila Real, apesar das muitas saudades que tinha de casa. À Doutora Ana Salbany que despertou o meu interesse no mundo dos mamíferos marinhos e me fez começar a lutar com um objetivo. Obrigada pelos conselhos na altura certa.
Aos treinadores excecionais que foram ouvindo com paciência todas as dúvidas da estagiária “Nunca Serás”, que espera um dia vir a ser. Pessoal do Aquário, das aves tropicais, das rapinas, mas em especial àqueles que treinam os animais mais mágicos e inteligentes do mundo, por quem eu me apaixonei: os mamíferos marinhos. Márcia, Hugo, Elsa, Lucie, Nadea, Vítor, Xana, Rita, Celine, Viegas, Rodolfo, Vanessa, Tiago, Sandra, Gui, Ticha, Zé. Obrigada pelos conhecimentos partilhados, pelos momentos divertidos, pelo carinho com que sempre me trataram.
Aos amigos que fizeram parte da minha vida algarvia e estiveram sempre lá: Ariana Carmelino, pela amizade que criamos, cumplicidade, momentos zen. Tiago Prata, pelas conversas infindáveis, companhia, por me ouvires sempre. Paula Irving, por me teres recebido tão bem, por me mimares, pelos momentos divertidos e jantares de sushi.
Aos amigos que ficaram longe da vista, mas perto do coração: Tomás Rito, Ana Marçal, João Zenha, Ângelo Jesus, Fábio Cruz.
Aos meus amigos, Sara Martins, Daniela Santos, André Dias e Gilberto Ferreira, sem vocês eu não seria a mesma. Obrigada por me ouvirem sempre, estarem sempre lá para mim, por serem os meus melhores amigos. Mesmo a 600 km nunca deixei de pensar em vocês.
À minha família algarvia, Dona Conceição Matias, Senhor Dino Matias, Andreia André, Ricardo André, Diana André e Simão André. Vocês foram os meus pilares, sem vocês as saudades de casa seriam triplicadas. Obrigada pela força, carinho, amor, paciência, companhia. Por nunca me abandonarem.
XV Ao Dino Matias, pela cumplicidade, pelo conhecimento, pelo carinho, por tudo o que só tu me dás e só tu sabes de mim.
À minha família, Avó Lucinda, Avô Alcídio, Avô António, Bá, Né, Lu, Fernando, Tio Mário, Tia Fernanda, João, Ana. Posso parecer ausente muitas vezes e sei que as saudades foram muitas de ambas as partes, mas estiveram sempre no meu coração.
À Marta, minha afilhada, pelo carinho, pela paixão. Desculpa não estar contigo sempre como gostávamos, mas um dia vais perceber o quão importante é seguir os teus sonhos.
À minha Avó Maria, por todo o mimo e amor que me deu. Sei que, esteja onde estiver, estará feliz por esta minha grande vitória.
Por último, às duas pessoas sem as quais eu não estaria aqui, os meus Pais, Isabel Pereira e Manuel Fernandes, por me terem educado desta forma, por todo o amor, carinho, apoio incondicional, por me deixarem seguir o meu sonho. Não imaginam o quanto vos fico grata por acreditarem em mim e me ajudarem a atingir os meus objetivos, mesmo quando as saudades são do tamanho do mundo.
Obrigado a todos. Nunca desistam dos vossos sonhos, porque eu nunca desistirei do meu.
1
1
Introdução
O estudo da microbiota bacteriana do trato respiratório superior do golfinho-roaz é um tema incontornável devido à crescente importância das doenças respiratórias em cetáceos que vivem sob cuidados humanos. Sendo a pneumonia bacteriana uma das principais causas de morte nestes animais torna-se urgente e essencial andar um passo à frente da doença, de modo a que esta possa ser detetada de forma precoce (Jeraj e Sweeney, 1996).
A microbiota normal é o termo que inclui bactérias, vírus, protozoários e fungos que habitam em diferentes regiões anatómicas. Estes microrganismos estão adaptados para viver dentro do hospedeiro sem causar doenças ou danos. Esta flora pode ser transitória ou permanente. A transitória coloniza por períodos curtos e é originada a partir de microrganismos ambientais, enquanto a de longa duração é a única que persiste por longos períodos de tempo, sendo semelhante entre os indivíduos da mesma espécie. Cada região anatómica cria o seu próprio ambiente seletivo onde alguns microrganismos são favorecidos em detrimento de outros. A interação entre o hospedeiro e o meio ambiente estabelece a microbiota normal, sendo que o próprio sistema imunitário acaba por criar uma certa tolerância para com esta (Avalos-Téllez et al., 2010).
Uma das formas de rastreio mais utilizada em programas de medicina preventiva é a colheita de amostras provenientes do espiráculo, que são coradas e depois observadas ao microscópio ótico e que poderão fornecer o primeiro alerta no caso de haver alguma alteração. Neste caso, o passo seguinte será realizar uma colheita de uma amostra asséptica do espiráculo, para que possa posteriormente ser cultivada e a partir da qual se vão isolar os microrganismos que poderão ser potencialmente patogénicos (Jeraj e Sweeney, 1996; Walsh et al., 2002).
Alguns microrganismos potencialmente patogénicos poderão estar presentes no trato respiratório do animal sem provocarem qualquer tipo de sintomatologia ou doença. Tudo depende do estado imune do animal, pois poderão ser agentes oportunistas (Lima et al., 2011). Assim, é de elevada importância compreender e estudar as bactérias presentes no trato respiratório destes animais de forma a perceber quais são as bactérias comensais, sem esquecer que cada animal é um caso específico e deve ser avaliado em conjunto com vários fatores individuais, tais como idade, raça, sistema hormonal, dieta, stress, comportamento sexual, medicação (caso esteja a ser feita), estação do ano, localização geográfica, habitat, densidade populacional, contacto entre animais e, até, procedimentos de limpeza do habitat (Avalos-Téllez et al., 2010).
2
2
Revisão bibliográfica
2.1 Golfinho-roaz
2.1.1 Características gerais e classificação
Golfinho nariz de garrafa, golfinho-roaz ou golfinho roaz-corvineiro: qualquer uma destas denominações pode ser usada para identificar os espécimes Tursiops truncatus que, na sua origem, significa “com aparência de golfinho” (Tursio
traduz-se por golfinho e o sufixo – ops quer dizer aparência) (Wells e Scott, 2002).
O golfinho-roaz (representado na figura 1) pertence ao Reino Animalia, Filo Chordatta, Sub-filo Vertebrata, Classe Mammalia, Subclasse Eutheria, Ordem Cetacea, subordem Odontoceti, Família Delphinidae, Género Tursiops e Espécie Tursiops truncatus (Reynolds III et al., 2000).
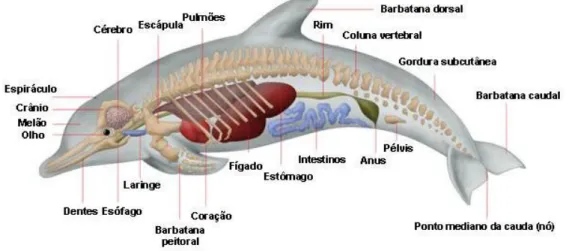
Estes apresentam um tamanho médio e um corpo robusto, com uma barbatana dorsal falciforme e uma coloração escura. Os adultos medem entre dois a aproximadamente quatro metros. A coloração do seu corpo é cinza escuro e apenas a parte ventral apresenta uma coloração mais clara (Wells e Scott, 2002). O seu corpo fusiforme apresenta duas barbatanas peitorais (figura 2), uma barbatana dorsal e uma caudal (figura 3). Os seus ouvidos são internos, sendo apenas visível um pequeno orifício exterior (Reynolds III et al., 2000).
Sob cuidados humanos os golfinhos podem viver mais que 50 anos. Recentemente o Zoomarine perdeu Sam com 50 anos, um dos machos mais velhos do Mundo que vivia sob cuidados humanos. Atualmente, existem duas fêmeas com mais de 50 anos que vivem sob cuidados humanos e, no Zoomarine, vive Cher, uma fêmea com 43 anos.
A sua longevidade tende a aumentar, visto que atualmente existem boas condições para este tipo de animais viveram sob cuidados humanos.
Figura 1- Golfinho-roaz.
[Fonte:albufeira.costasur.com]
Figura 2- Golfinho-roaz (barbatanas peitorais).
3
Figura 3- Principais características anatómicas do Golfinho-roaz.
2.1.2 Biologia e fisiologia reprodutiva
Em relação ao dimorfismo sexual, os machos tendem a ser maiores do que as fêmeas. Eles apresentam duas fendas, uma urogenital e a anal, enquanto elas apresentam uma única fenda, que inclui a urogenital e a anal. Lateralmente a esta, as fêmeas apresentam de ambos os lados duas fendas mamárias onde encontramos os mamilos (Reynolds III et al., 2000). As glândulas mamárias nestes animais são ventrais, mediais e ligeiramente caudais (figura 4) (Rommel e Lowenstine, 2001).
A maturidade sexual é atingida pelos 10 anos nas fêmeas no meio selvagem. Estas são poliéstricas sazonais, ovulando esporádica e
espontaneamente. O seu período de gestação é de aproximadamente um ano e a lactação, no meio selvagem, pode durar até 2 anos ou mais, sendo que é um dos fatores que suprime o ciclo éstrico. O intervalo entre crias poderá rondar os 3-4 anos, sendo superior nas fêmeas jovens e idosas (Rommel e Lowenstine, 2001; Wells e Scott, 2002).
Estudos realizados em fêmeas da espécie Tursiops truncatus aduncus comprovaram que o ciclo éstrico dura, aproximadamente, 30 dias. Podem ainda existir períodos de anestro não associados com a gestação ou lactação nesta família. Em relação à
pseudogestação, de referir que esta pode ocorrer em cetáceos, havendo uma maior frequência em fêmeas que não têm acesso a machos (Rommel e Lowenstine, 2001).
[Fonte: Adaptado de en.wikipedia.org/wiki/File:Dolphin_anatomy.png]
[Fonte:roazesdosado.icnb.pt]
Figura 4- Pormenor das diferenças ao nível dos genitais do golfinho-roaz fêmea e macho.
4 No que toca à monitorização da gestação, a ecografia é o método auxiliar mais utilizado (Rommel e Lowenstine, 2001). No Zoomarine existe um Protocolo Médico criado especificamente para fêmeas gestantes, no qual, através de um acompanhamento ecográfico semanal, é possível observar e medir o corpo lúteo e executar medições dos diâmetros craniano (distância inter-parietal) e torácico do feto bem como observar a sua anatomia em geral, o cordão umbilical e o batimento cardíaco e, posteriormente, introduzir estes dados num programa informático que os utiliza para prever a data do parto. Numa altura mais próxima da data do parto, é feita a medição da temperatura corporal duas vezes por dia e da distância intermamária. Além disso, são feitas colheitas de sangue regulares às fêmeas gestantes para controlar os níveis de progesterona e estradiol. É ainda de referir que o Zoomarine tem um programa de observações que é iniciado antes da data prevista do parto e só termina um mês depois da cria nascer, em que há um acompanhamento diário de 24 horas por um observador, sendo registados vários comportamentos, como: alimentação, quando defeca e urina, contrações e distensões (quando existem e de que grau são), a frequência respiratória, entre outros (comunicação pessoal).
Nos machos, a maturidade sexual é atingida, normalmente, entre os 10 e os 13 anos. Estes podem estar ativos durante todo ano. Normalmente, os picos de testosterona estão associados ao período de reprodução, acontecendo entre Abril e Maio e depois, novamente, em Setembro e Outubro. Estes normalmente não estão associados aos picos de produção de espermatozóides, que acontecem depois disto. Os animais jovens mantidos sob cuidados humanos poderão procriar com sucesso mais cedo em relação aos animais que vivem em estado selvagem (Rommel e Lowenstine, 2001; Wells e Scott, 2002).
2.1.3 Dieta
Uma dieta variada e equilibrada é um fator essencial na manutenção da saúde dos mamíferos marinhos. No caso dos animais que vivem sob cuidados humanos, o arenque (Clupea harengus), a cavala (Scomber japonicus) e a sardinha (Sardinella sp.), como são espécies de peixe com maior teor em gordura são suplementados com peixes mais magros como, por exemplo, o capelim (Mallotus villosus), o verdinho (Micromesistius poutassou) e o eperlano-arco-íris (Osmerus mordax). Nos invertebrados, a lula (Loligo vulgaris) é o exemplar mais usado (Couquiaud, 2005).
Em cada sessão de alimentação os golfinhos devem ingerir pelo menos duas espécies diferentes, para que não haja habituação a uma espécie concreta e comece a haver uma recusa em relação a outras espécies e também por uma questão de enriquecimento. Há uma variação na quantidade e nas espécies oferecidas. A dieta de um golfinho jovem depois do desmame é idêntica à de um adulto (Immerzeel e Lotens, 2005).
5 No caso específico do Zoomarine, a dieta dos mamíferos marinhos é apresentada na tabela seguinte (Tabela 1):
Tabela 1- Espécies de peixes e cefalópode consumidos pelos animais do parque.
Nome comum Nome científico Categoria
Cavala Scomber japonicus Peixes azuis ou gordos
Arenque Clupea harengus Peixes azuis ou gordos
Capelim Mallotus villosus Peixes brancos ou magros
Verdinho Micromesistius poutassou Peixes azuis ou gordos
Carapau Trachurus trachurus Peixes brancos ou magros
Espadilha Sprattus sprattus Peixes azuis ou gordos
Lula Loligo vulgaris Cefalópodes
De referir ainda que no parque a quantidade de alimento a fornecer a cada animal é calculada com base na sua taxa metabólica basal (o que corresponde à energia necessária para manter as funções vitais) e nas quilocalorias fornecidas por cada espécie de peixe. O peixe é enviado para análise nutricional e é possível saber a quantidade exata de quilocalorias por quilo de cada lote. Todo o peixe ingerido por estes animais é submetido a análise de qualidade e está apto para consumo humano (comunicação pessoal).
Em relação à água consumida por estes animais, a maior parte desta provém da alimentação fornecida. No entanto poderá haver necessidade destes ingerirem pequenas quantidades de água “extra” à alimentação (Worthy, 2001).
2.1.4 Suplementação vitamínica
Pelo facto do peixe fornecido aos mamíferos marinhos ser congelado é essencial ter em conta as perdas de nutrientes na congelação, armazenagem e descongelação deste e compensá-las com uma suplementação vitamínica (comunicação pessoal).
No caso específico do Zoomarine, os golfinhos são suplementados com Akwavit®, que contém as vitaminas A, E, C, B1, B2 e B6, ácido pantoténico, ácido fólico e biotina.
Este tipo de suplemento vitamínico deve ser fornecido diariamente, pelo menos uma hora antes da primeira sessão de alimentação, para que haja uma correta absorção pelo organismo. A dose fornecida a cada animal vai depender de vários fatores, tais como o estado reprodutivo, estado de saúde, atividade física e idade (White e Francis-Floyd, 1988)
6
2.2 Sistema respiratório dos mamíferos marinhos
2.2.1 Adaptações para uma vida marinha
Para os mamíferos que estavam originalmente adaptados a viver na terra, a vida aquática impôs muitos desafios. Assim, os mamíferos marinhos apresentam várias características que advêm da sua anterior existência em terra. Os mamíferos marinhos não respiram como os peixes; assim, foi necessário um conjunto de alterações que protegesse um sistema concebido para funcionar ao ar, característico dos mamíferos, que excluísse a água que os rodeia permanentemente. Tanto as narinas valvulares como a laringe que existe dentro da narina são exemplos disso (figura 5). As primeiras não permitem a entrada de água e a última protege a inundação do trato respiratório quando os animais engolem (Reidenberg, 2007).
Figura 5- Representação esquemática das vias respiratória e digestiva de um golfinho-roaz.
No caso dos cetáceos, esta narina trata-se do espiráculo, que sofreu uma migração da parte rostral para a parte dorsal, ao longo da evolução, como é possível observar na figura 5. Desta forma, torna-se mais fácil expor o espiráculo à superfície da água durante a natação. Assim, estes animais nunca respiram pela boca. Na realidade, a traqueia e o esófago estão separadas por um órgão chamado bico de ganso (goose beak). O bico de ganso é uma laringe evolutivamente modificada que preenche as lacunas entre as passagens nasais e a traqueia e
7 existe para manter apenas e só ar nos pulmões. Este órgão separa o trato respiratório do digestivo. Desta forma, não existe qualquer incursão nas vias aéreas durante a alimentação (Reidenberg e Laitman, 2008).
Em relação à natação, é importante também ter em conta que a pressão que está ligada à profundidade que estes atingem é bastante elevada e que o sistema respiratório teve que se adaptar para que os pulmões não sofressem qualquer tipo de dano. Essas adaptações podem ser verificadas na cavidade nasal óssea, que se encontra revestida com tecidos moles e flexíveis, que podem aumentar de volume para ocupar o espaço criado pelo ar colapsado em profundidade, sendo que também facilitam a troca rápida de um grande volume de ar de cada vez que o animal vai à superfície. Além disso, as vias aéreas estão reforçadas com anéis circulares de cartilagem que podem sofrer uma deformação parcial em profundidade. A caixa torácica destes mamíferos diferencia-se da dos mamíferos terrestres pois é articulada e compressível e permite a diminuição do volume pulmonar consoante a profundidade (Reidenberg e Laitman, 2008). Na figura 6 é possível observar algumas destas alterações e compará-las com o sistema respiratório de um mamífero terrestre.
Figura 6- Anatomia comparada das vias respiratória e digestiva de um mamífero terrestre e um mamífero marinho (golfinho-roaz).
Cada passagem de ar termina num frágil alvéolo, onde de facto ocorrem as trocas gasosas, que está reforçado com tecido muscular e/ou cartilaginoso que o mantém aberto mesmo quando a profundidade do mergulho é suficiente para colapsar os pulmões. No entanto, torna-se benéfica a paragem de trocas gasosas durante o mergulho para que não haja uma grande quantidade de nitrogénio a sair dos pulmões e a entrar em circulação (Reynolds III et al., 2000).
8 Outra modificação, e talvez a mais radical, é a ausência de seios ósseos paranasais. A sua presença no caso dos mamíferos marinhos poderia ser um risco estrutural para o seu crânio, graças às altas profundidades e pressões que estes atingem em mergulho. Devido à incapacidade dos tecidos ósseos de se contrair ou expandir o crânio poderia fraturar. Assim, alguns mamíferos marinhos, como os cetáceos, desenvolveram um crânio sem seios ósseos paranasais, desenvolvendo em compensação sacos aéreos que possuem paredes moles que apesar de estarem relacionados com a via aérea permanecem fora do crânio (Reidenberg e Laitman, 2008).
2.2.2 Pulmões, pleura e linfonodos relacionados nos cetáceos
Os pulmões dos cetáceos são pequenos, simétricos e sem lobulação externa (Geraci e Lounsbury, 1993). No caso específico do golfinho-roaz, a porção crânio-ventral do pulmão esquerdo (que cobre o coração) é muito estreita. Nestes animais verifica-se a presença do brônquio apical (traqueal) (Rommel e Lowenstine, 2001). A pleura é usualmente espessa, bem vascularizada (Geraci e Lounsbury, 1993).
Em relação aos linfonodos, existem três grupos constantemente associados ao sistema respiratório: os linfonodos hilares (hilar nodes), o linfonodo da margem anterior livre (anterior free margin node) e o linfocentro do diafragma (diaphragmatic node mass). Em estudos recentes foi reconhecida uma complexa estrutura linfoglandular na laringe de animais da espécie Tursiops truncatus que ocorre em muitas espécies de cetáceos e parece ser análoga da adenóide (Cowan e Smith, 1999).
Os linfonodos hilares (linfonodos traqueobronquiais craniais) estão associados aos brônquios principais na sua inserção no pulmão. Estes são cobertos pela pleura do pulmão e não são facilmente visíveis, mas podem sobressair ligeiramente na superfície do pulmão (Cowan e Smith, 1999).
O linfonodo marginal do pulmão existe na margem (ventral) anterior livre de cada pulmão, onde se cruza com a superfície do diafragma. É grande e triangular. Existem uma série de vasos linfáticos bastante proeminentes que convergem, sobre a superfície do pulmão, para este linfonodo. Na maioria das vezes, os linfonodos marginais de ambos os pulmões estão aderentes um ao outro através da linha média ou pela pleura parietal do esterno (Cowan e Smith, 1999).
O linfocentro do diafragma é plano, em forma de U e está ligado à face diafragmática de ambos os pulmões e ao diafragma, situando-se atrás do pericárdio, centrado em torno do esófago e do hiato diafragmático. Normalmente, os linfocentros de ambos os lados unem-se formando um aglomerado de linfonodos parcialmente fundidos. Este aglomerado é usualmente
9 plano, mas caso estejamos na presença de uma reação inflamatória torna-se espesso e nodular (Cowan e Smith, 1999).
2.2.3 Doenças do trato respiratório dos cetáceos: a importância relacionada da recolha de amostras do espiráculo
As doenças do trato respiratório são um fator importante a ter em conta na saúde dos cetáceos. No caso particular da pneumonia bacteriana, esta representa uma das maiores causas de morte em golfinhos mantidos sob cuidados humanos. Estas infeções estão normalmente associadas a uma predisposição da parte do animal, podendo ser uma afeção aguda ou crónica, dependendo da bactéria em causa e do estado imune do animal. Os sinais
clínicos podem incluir apatia, anorexia, tosse, cheiro fétido do ar expirado, taquipneia, dispneia, ruídos respiratórios, intolerância ao exercício, depressão e alterações de comportamento (Jeraj e Sweeney, 1996; Walsh et al., 2002). Desta forma, torna-se imprescindível diagnosticar a doença respiratória o mais cedo possível para que se possa instituir rapidamente uma terapêutica adequada (Jeraj e Sweeney, 1996).
Torna-se, desta forma, essencial ter conhecimento da flora normal do trato respiratório para se perceber se alguns destes microrganismos são oportunistas e poderão ser os agentes causadores de afeções respiratórias que virão a desenvolver sinais clínicos dentro de um curto espaço de tempo (Lima et al., 2011). Assim, usando uma técnica não invasiva,
podemos facilmente
recolher amostras representativas do trato respiratório superior destes animais através de um comportamento para o qual estes são treinados para expirar fortemente pelo espiráculo para um recipiente como, por exemplo, uma placa de Petri esterilizada. É, no entanto, necessário limpar e desinfetar a zona do espiráculo antes da colheita e secá-la para evitar possíveis contaminações. Quanto à
Figura 7- Candida spp. numa preparação de azul de metileno (40x).
[Fonte: Zoomarine]
Figura 8- Pormenor de uma célula epitelial, leucócitos e muco numa prepraração de azul de metileno (40x).
[Fonte: Zoomarine]
Figura 9- Pormenor de uma célula epitelial com várias bactérias numa preparação de azul de metileno (40x).
10 obtenção de uma amostra do trato respiratório inferior a lavagem broncoalveolar via broncoscopia é recomendada, apesar de ser uma técnica invasiva (Varela et al., 2007).
Os achados normais numa citologia do espiráculo incluem células escamosas e colunares epiteliais (figura 8) e quantidades variáveis de bactérias (figura 9), leveduras e fungos (figura 7 e 10). Na ausência de inflamação, a presença de uma pequena quantidade de protozoários ciliados e ovos de tremátodes poderão ser considerados normais. Caso estejam presentes vários leucócitos ou eritrócitos, estes achados já deverão ser considerados anormais. É de salientar que a presença de uma pequena quantidade de leucócitos é considerada normal. Apenas se estes existirem em quantidade superior às células epiteliais poderá estar presente uma reação inflamatória. Esta será considerada na presença de neutrófilos degenerados e em banda, de grandes quantidades de bactérias ou parasitas (Varela et al., 2007).
2.3 Bactérias da microbiota do trato respiratório superior
São várias as bactérias que podem colonizar o trato respiratório superior do golfinho-roaz. Algumas delas são apresentadas seguidamente, bem como as suas principais características.
2.3.1 Streptococcus spp.
Streptococcus spp. são cocos Gram positivos que aparecem aos pares ou em cadeias de vários tamanhos, sendo que cada coco mede aproximadamente 1.0 µm de diâmetro. São bactérias anaeróbias facultativas, negativas às provas da catalase, oxidase e mobilidade, pois são imóveis (Quinn et al., 2002). O nome Streptococcus foi utilizado pela primeira vez por Rosenbach em 1884 para descrever um microrganismo de forma esférica e que apresentava
Figura 10- Pormenor de estruturas
fúngicas numa coloração de Gram feita a partir de uma amostra do espiráculo.
11 um crescimento em cadeias, tendo sido isolado de lesões supurativas de humanos (Hardie e Whiley, 1997).
As bactérias deste género são fastidiosas, ou seja, exigem mais do que os meios base para crescerem. Com efeito, crescem em meios com sangue ou soro adicionado (Quinn et al., 2002). Uma das características taxonómicas mais importantes passa pela sua capacidade de lisar os eritrócitos quando crescem em agar sangue, produzindo dois tipos de hemólise: β onde há a produção de uma zona clara de hemólise à volta da colónia, com marcada mudança de cor (por exemplo, S. pneumoniae e S. viridans) e α onde há uma zona de hemólise parcial de cor esverdeada a circundar a colónia (por exemplo, S. pyogenes e S. agalactiae). Quando não se verifica hemólise diz-se que a colónia é γ-hemolítica (Prescott et al., 1999). O teste de sensibilidade a 0,04 unidades de bacitracina distingue Streptococcus do grupo A (por exemplo, S. pyogenes) de outros Streptococcus que produzem β-hemólise adicionado (Quinn et al., 2002).
A maior parte das espécies de interesse Veterinário pertencentes a este género são comensais da mucosa do trato respiratório superior e trato urogenital inferior. Estes são sensíveis à dessecação e, como tal, não resistem muito tempo longe do hospedeiro. Tal como o género Staphylococcus, estão frequentemente associados à supuração e formação de abcessos (Quinn et al., 2002). Duas das espécies mais patogénicas são Streptococcus pyogenes e S. pneumoniae (Prescott et al., 1999).
Streptococcus pneumoniae é um dos mais comuns causadores de afeções do trato respiratório superior e inferior. Inicialmente, estas infeções eram facilmente resolvidas com penicilina. No entanto, várias estirpes começaram a apresentar resistências, não só à penicilina como a múltiplos antibióticos. Isto poderá acontecer por um uso excessivo prévio à infecção (Kaplan e Mason, 1998).
No caso de Streptococcus pyogenes este é um dos agentes patogénicos mais comuns em humanos, pois provoca várias doenças, sendo as mais frequentes a faringite aguda e a febre reumática aguda. Este microrganismo pode causar uma variedade de infeções da pele e tecidos moles, muitos dos quais que podem mesmo vir a ser fatais (Bisno e Stevens, 1996).
2.3.2 Aeromonas salmonicida
As bactérias Aeromonas são bastonetes Gram negativos que tanto se podem apresentar em linha reta como em curva. São bactérias anaeróbias facultativas, positivas à prova da catalase e da oxidase e móveis devido aos seus flagelos polares, exceto A. salmonicida que é imóvel. Todas as espécies crescem em agar MacConkey onde fermentam a glucose com produção de ácido e algumas espécies também produzem gás. A temperatura ótima de crescimento é inferior a 35ºC (Quinn et al., 2002).
12 Aeromonas spp. está presente na água, esgotos e solo, sendo que a sua quantidade aumenta com o aumento da presença de matéria orgânica. Os animais podem ser portadores de Aeromonas spp. nas fezes. São organismos oportunistas dos peixes e répteis e, raramente, dos mamíferos (Quinn et al., 2002).
Aeromonas salmonicida é um parasita obrigatório de peixes salmonídeos, podendo também afetar as carpas. Esta causa furunculose e úlceras. Em agar sangue, forma colónias pequenas que produzem hemólise após 48 horas. Microscopicamente, tende a ser cocos organizados em pares, cadeias ou aglomerados. Em agar Furunculose® (Difco) cresce com um pigmento castanho típico, não apresenta mobilidade, produz gás da fermentação da glucose, é negativa à prova do indol, positiva às provas da catalase e oxidase e hidrolisa a esculina (Quinn et al., 2002).
Foi demonstrado que algumas espécies de bactérias Gram negativas conseguem entrar num estado fisiológico em condições fracas em nutrientes, sendo ainda viáveis mais incapazes de se reproduzirem. Uma delas é Aeromonas salmonicida. A existência de bactérias neste estado na água pode explicar a infeção de populações de peixes com furunculose que, aparentemente, nunca teriam estado em contacto com o agente patogénico. No entanto, é de referir que diversos estudos comprovaram a sobrevivência apenas por curtos períodos em amostras de água, havendo maior capacidade de sobreviver em grãos de areia ou sedimentos dos rios (Morgan et al., 1991).
Como já foi referido anteriormente, A. salmonicida é o agente causador de furunculose, uma das doenças bacterianas mais destrutivas em peixes salmonídeos de aquacultura (Barnes et al., 1990). Os sinais clínicos apresentados são septicemia hemorrágica com necrose dos tecidos e esplenomegalia (Vaughan et al., 1993).
2.3.3 Staphylococcus spp.
Staphylococcus spp. são cocos Gram positivos que se organizam em clusters irregulares com aspeto de cachos de uvas, sendo o seu diâmetro médio 1.0 µm (Quinn et al., 2002). Este género foi pela primeira vez reconhecido por Pasteur, sendo associado ao pus (Hartmann, 1997).
As bactérias deste género são anaeróbias facultativas, catalase-positivas, oxidase-negativas, não móveis e fermentam a glucose em anaerobiose (Prescott et al., 1999).
O seu crescimento acontece em agar nutritivo e também em agar sangue, mas não em agar MacConkey. As espécies mais patogénicas (Staphylococcus aureus, S. intermedius, S. hyicus, entre outros) são positivas à prova da coagulase, enquanto as espécies comensais e que se encontram no ambiente, como S. epidermidis e S. saprophyticus, são negativas à mesma prova (Quinn et al., 2002). Algumas das espécies deste género já foram descritas em
13 vários meios, nomeadamente solo, ar, água doce e salgada e superfície das plantas (Bartolomeu et al., 2011).
Os membros desta família colonizam a pele e o trato respiratório superior dos mamíferos. São bactérias piogénicas e estão associadas à supuração e formação de abcessos (Quinn et al., 2002). A capacidade de induzir infeção em qualquer órgão ou tecido do organismo passa pela sua multiplicação e disseminação, bem como, pela produção de várias substâncias extracelulares, bastando para isso que haja um comprometimento dos mecanismos de defesa do organismo em causa (Prescott et al., 1999).
Pode também ser associado a afeções gastrointestinais, apresentando potentes toxinas que funcionam como antigénios, pois estimulam a produção de células T não específicas. Estas enterotoxinas são resistentes ao calor e manifestam-se após um curto período de tempo da sua ingestão produzindo náuseas, vómito, dor abdominal e diarreia (Harris et al., 1993; Balaban e Rasooly, 2000).
Em 1946, Spink e Ferris referiram que investigadores deram conta de estirpes de Staphylococcus coagulase-positivas que seriam naturalmente resistentes à penicilina. Isto acontece devido à existência de plasmídeos codificados para a penicilinase. No entanto, Staphylococcus estariam entre as espécies de bactérias Gram positivas que seriam sensíveis à ação antibacteriana da penicilina. A tolerância a esta acontece quando há falha das enzimas autolíticas da parede das células (Spink e Ferris, 1946; Quinn et al., 2002).
Em relação aos cetáceos, Staphylococcus delphini foi isolado da pele de golfinhos (Quinn et al., 2002).
2.3.4 Enterococcus spp.
Tal como no género Streptococcus, Enterococcus spp. são cocos Gram positivos que normalmente ocorrem em pares ou cadeias curtas. São também anaeróbios facultativos, negativos às provas da catalase e oxidase. Em relação à mobilidade há exceções, pois nem todos são imóveis (Quinn et al., 2002).
Estes microrganismos apresentam um crescimento rápido em agar sangue, sendo visíveis colónias brancas grandes após 24 horas de incubação. Tipicamente estas colónias são não hemolíticas, mas também poderão ocasionalmente apresentar hemólise α ou β. Além disso, crescem em meios com 6,5% de NaCl e em agar MacConkey com 40% de sais biliares, a 10 e 45 ºC e podem resistir até trinta minutos a 60 ºC (Manero e Blanch, 1999).
As bactérias deste género são bactérias entéricas, ou seja, colonizam o trato gastrointestinal, podendo também aparecer ocasionalmente noutras partes do corpo, como a cavidade oral ou o trato vaginal (Devrise et al., 1992; Muller et al., 2001; Pabich et al., 2003). Por exemplo, E. faecalis, são oportunistas e vivem no trato intestinal de vários animais (Quinn
14 et al., 2002). Estas podem causar infeções do trato urinário e endocardite (Prescott et al., 1999) apresentando grande capacidade para adquirirem resistência a agentes microbianos (Caballero-Granado et al., 1998). A maioria das infeções clínicas nos humanos são originadas por E. faecalis e E. faecium (Lautenbach et al., 1999; Giacometti et al., 2000; Higaki et al., 2002).
Ambos os géneros, Streptococcus e Enterococcus, têm vindo a ser propostos como indicadores de contaminação fecal da água, pois existem em abundância nas fezes dos animais e a sua capacidade de sobreviver no ambiente durante longos períodos de tempo (Manero e Blanch, 1999).
A resistência aos antibióticos é um dos fatores mais importantes que poderão contribuir indiretamente para a patogenicidade das estirpes desta família. No entanto, isso não explica a sua virulência, que se relaciona com fatores como a adesão, invasão e evasão à resposta imunitária. Desta forma, o microrganismo consegue resistir aos mecanismos de defesa do hospedeiro, o que permite a sua colonização (Johnson, 1994; Gilmore et al., 2002).
2.3.5 Flavobacterium spp.
Este género de bactérias pode ser observado como coco-bacilos Gram negativos que não fermentam a glucose. Crescem em agar MacConkey onde produzem α-hemólise, sendo positivas às provas da oxidase e da catalase e negativas à da mobilidade. Estes organismos fastidiosos e de crescimento lento (Crump et al., 2001). São geralmente encontrados no solo e na água, em lagos, lagoas, rios e mar (Ratner, 1984; Quinn et al., 2002).
Estes microrganismos são colonizadores ubiquitários do ambiente hospitalar, provavelmente, devido à sua capacidade de crescer em meios não nutrientes. Flavobacterium já foi isolado a partir de máquinas de gelo, frascos de drogas intravenosas, nebulizadores, desinfetantes tópicos e tubos de alimentação (Ratner, 1984).
Apesar de não ser comumente considerado um agente patogénico humano, Flavobacterium meningosepticum causa várias infeções nosocomiais, que incluem meningite, bacteriemia pós-operatória e endocardite (Ratner, 1984).
Algumas espécies deste género são patogénicas para os peixes como, por exemplo, Flavobacterium psychrophilum, que causam “doença bacteriana de água fria” (bacterial cold water disease) e “síndrome da truta arco-íris frita” (rainbow trout fry syndrome) nos salmonídeos (Nematollahi et al., 2003). A terapia antimicrobiana continua a ser a única opção visto que ainda não se encontra disponível uma vacina específica, sendo defendido o uso de sulfonamidas e oxitetraciclina (Nematollahi et al., 2003; Kim et al., 2010).
No entanto, algumas resistências a agentes antimicrobianos começaram a ser descritas, tais como a existência de estirpes resistentes às quinolonas e à rifampicina (Izumi et al., 2007;
15 LaFrentz et al., 2008). Assim, torna-se urgente encontrar terapias alternativas para estas doenças, podendo uma das opções passar pelo isolamento e identificação de bacteriófagos (Kim et al., 2010).
2.3.6 Morganella morganii
Esta bactéria pertence à família Enterobacteriaceae um bacilo Gram negativo de tamanho médio que cresce em agar MacConkey e não fermenta a lactose. É positiva à prova da catalase e negativa à da oxidase (Quinn et al., 2002). É ainda positiva à prova do vermelho de metilo e apresenta mobilidade (Janda et al., 1996). Cresce rápida e otimamente entre 15 ºC e 25 ºC, sendo que abaixo dos 15 ºC o seu crescimento é dramaticamente reduzido (Klausen e Huss, 1987).
Inicialmente, esta espécie foi colocada no género Proteus. A maior parte dos membros do conjunto Proteus-Providencia-Morganella partilham várias caraterísticas bioquímicas, algumas delas já referidas acima, tais como reação positiva ao vermelho de metilo e à prova da urease e o fato de apresentarem mobilidade. Fenotipicamente, são três as principais características comuns: a produção de fenilalanina deaminase, a degradação de cristais de tirosina e a produção de um pigmento castanho avermelhado na presença de DL-triptofano. É de salientar que todas estas características raramente são observadas noutros membros da família das Enterobacteriaceae (Janda et al., 1996).
Estudos posteriores de DNA revelaram que não havia uma relação mais estreita com o género Proteus do que com qualquer outro membro desta família. Como tal, separaram para o seu próprio género: Morganella (Janda et al., 1996).
Em geral, é encontrada em fezes de animais, podendo infetar o trato urinário e auditivo de cães e gatos (Quinn et al., 2002). Em 1990, Hu et al. constataram que Morganella morganii era uma causa crescente de bacteriúria nosocomial associada ao uso de cateter, sendo que a maioria das isoladas neste caso eram ureases positivas e com múltiplas resistências antibióticas.
No caso específico do trato urinário dos humanos esta capacidade de Morganella morganii hidrolisar a ureia tem sido associada à formação de cálculos de xantina e estruvite. O tratamento baseia-se no uso de um inibidor da urease e também de um antibiótico de largo espetro (Hu et al., 1990).
Em 1996, Young et al. concluíram que Morganella morganii sobrevive em condições de pH ácido, ao contrário de outras bactérias Gram negativas e urease positivas, como Escherichia coli e Pseudomonas aeruginosa. Assim, a urease é ativada in vitro a um pH muito baixo, tendo a sua atividade ótima a um pH 5,5, o que é pouco comum.
16 2.3.7 Escherichia coli
Tal como Morganella morganii, E. coli pertence à família Enterobacteriaceae e apresenta, portanto, características similares. É um bacilo Gram negativo de tamanho médio que cresce em agar MacConkey em anaerobiose facultativa e fermentadora de lactose. Em agar de TSI (triple sugar iron) fermenta todos os hidratos de carbono, dando ao meio uma cor amarela uniforme. É positiva à prova da catalase, ao vermelho de metilo e ao indol, negativa à da oxidase e a maioria são móveis, possuindo flagelos periféricos (Quinn et al., 2002; Thielman e Guerrant, 1999).
A maioria das estirpes desta bactéria sintetiza a vitamina K e B12 e vive comensalmente no trato intestinal de todos os mamíferos, sendo que está presente em maior quantidade nos carnívoros e omnívoros do que nos herbívoros. A sua excreção ocorre nas fezes e tem a capacidade de sobreviver em partículas fecais, pó e água durante semanas a meses (Quinn et al., 2002; Thielman e Guerrant, 1999). No intestino, E. coli desempenha um papel protetor contra infeções e mesmo outras bactérias comensais, pois compete com estas, tais como Shigella e Salmonella, que podem ser patogénicas intestinais. Apesar de comensal, E. coli é responsável por várias toxinfeções alimentares e também afeções extra-intestinais, sejam abdominais ou do trato urinário, meningite, ou no caso de entrar na corrente sanguínea, septicemia (Orskov e Orskov, 1985; Guerra et al., 2003; Costa, 2006). As estirpes patogénicas desta bactéria apresentam fatores de virulência que não são comumente encontrados nas estirpes comensais (Sáenz, 2004). A sua capacidade de causar doença pode passar por vários fatores, como imunidade passiva insuficiente (no caso dos neonatos) ou práticas de pouca higiene (Quinn et al., 2002).
Graças à sua capacidade de sobrevivência na água durante meses, como referido acima, torna-se útil realizar análises microbiológicas de amostras de água para garantir que esta é segura. Existem muitos organismos patogénicos que podem ser associados à água e, não sendo possível testar as amostras de água para cada um desses agentes, usam-se alguns microrganismos indicadores, que acabam por ser marcadores de risco (Barrell et al., 2000; Quinn et al., 2002). A presença de E. coli é um desses marcadores e indica contaminação fecal (Min e Baeumner, 2002).
É importante referir que E. coli possui uma capacidade notável não só para adquirir genes de resistência mas também para os transmitir. Esta capacidade é comum aos Enterococcus e aplica-se mesmo a bactérias filogeneticamente afastadas (Martel et al., 2003; Costa, 2006).
Num estudo realizado por Greig et al. em 2007 foi possível isolar E.coli em fezes de vários animais da espécie Tursiops truncatus, sendo que cerca de metade apresentavam resistência a um ou mais antibióticos.
17 2.3.8 Pseudomonas aeruginosa
Pseudomonas aeruginosa é uma bactéria Gram negativa baciliforme de tamanho médio que é positiva às provas da catalase, oxidase e mobilidade (Quinn et al., 2002). Quanto às exigências de oxigénio estes microrganismos são aeróbios estritos (Prescott et al., 1999), sendo que podem ser encontrados no solo, na água e na vegetação (Delden e Iglewski, 1998), na pele, membranas mucosas e fezes de animais (Quinn et al., 2002).
O surgimento desta bactéria como oportunista pode dever-se à sua resistência a antibióticos e desinfetantes que eliminam outras bactérias. Por essa razão, P. aeruginosa tornou-se uma fonte significativa de bacteriemia em vítimas de queimaduras, infeções do trato urinário, pacientes cateterizados e pacientes com pneumonia adquirida a nível hospitalar, sendo a bactéria Gram negativa mais frequentemente encontrada em infeções nosocomiais, considerada por isso um agente patogénico oportunista cada vez mais prevalente (Stover et al., 2000; Delden e Iglewski, 1998). As causas predisponentes incluem trauma dos tecidos, imunodeficiência ou reduzida quantidade de flora normal, que normalmente é causada pela antibioterapia, que muitas vezes não afeta esta espécie devido à grande quantidade de resistências que apresenta aos antibióticos mais comuns (Quinn et al., 2002). No caso especifico dos doentes com fibrose quística, a situação é bastante grave, visto que o epitélio anormal das vias aéreas permite uma colonização a longo prazo nos pulmões de P. aeruginosa. Este tipo de infeção é impossível de erradicar, em parte devido à resistência natural desta bactéria a antibióticos. Numa última análise, este tipo de infeções leva à insuficiência pulmonar e, posteriormente, à morte (Stover et al., 2000).
Em 1998, O’Toole e Kolter referiram que foi demonstrado que P. aeruginosa formava biofilmes numa grande variedade de superfícies, incluindo tecidos com fibrose quística do pulmão e superfícies abióticas, tais como lentes de contacto. Estes biofilmes são formados por células planctónicas individuais, através de um complexo e controlado processo de desenvolvimento.
Em 1999, Carmeli et al. concluíram que a resistência a agentes antibióticos é um problema clínico em ascensão, sendo que P. aeruginosa possui uma elevada propensão para o desenvolvimento de resistências. Este facto poderá limitar futuras escolhas terapêuticas, estando ainda associado a um aumento dos níveis de mortalidade e morbilidade, bem como dos custos.
18
3
Objetivos
Devido à crescente importância das doenças respiratórias em cetáceos este trabalho pretende obter resultados que permitam caracterizar a microbiota do trato respiratório superior de vários exemplares da espécie Tursiops truncatus (golfinho-roaz) que vivem sob cuidados humanos, bem como comparar esta com estudos anteriores realizados pelo Zoomarine com dados recolhidos entre o ano de 2005 e o de 2011.
Como tal, definiram-se os seguintes objetivos: Isolar agentes da microbiota respiratória;
Comparar os isolados obtidos da microbiota respiratória com os registos de anos anteriores;
Fazer a caracterização bacteriológica da água dos habitats dos animais em estudo;
Comparar os isolados obtidos da microbiota respiratória com as bactérias encontradas nas águas dos habitats.
19
4
Material e métodos
4.1 Isolamento de bactérias provenientes da microbiota do trato respiratório superior do golfinho-roaz
4.1.1 Origem das amostras
As amostras utilizadas neste trabalho foram recolhidas no Zoomarine (Mundo Aquático S.A.). O Zoomarine é um parque oceanográfico de entretenimento educativo privado que foi inaugurado a 3 de Agosto de 1991. Este localiza-se na região do Algarve, mais concretamente na Guia, no Concelho de Albufeira, sendo as suas coordenadas geográficasN 37° 7' 37,9266", W 8° 18' 50,904".
O clima nesta região que é caracterizado por Verões longos e quentes, enquanto os Invernos são amenos e curtos, com a precipitação concentrada no Outono e Inverno (principalmente entre Outubro e Fevereiro), havendo um baixo número anual de dias com precipitação e um alto número de horas de sol. Assim, está-se na presença de um clima temperado, onde a temperatura média anual ronda os 18ºC.
Em relação ao parque, este proporciona o contacto com diferentes espécies, tanto exóticas como selvagens, e fornece informação sobre as mesmas. Além disso, reabilita animais arrojados e procura sensibilizar os visitantes, sendo fiel ao seu tema “Manter a Vida e Preservar a Natureza”.
As amostras do trato respiratório superior utilizadas neste estudo foram provenientes do espiráculo de um grupo de nove golfinhos-roazes que fazem parte de uma população estável que vive há vários anos sob cuidados humanos, sendo seis fêmeas e três machos com idades compreendidas entre os 2 e os 43 anos, que pesam entre os 95 e os 222 quilos e medem entre os 2 metros e 7 centímetros e os 2 metros e 67 centímetros. Estas amostras foram obtidas através de um comportamento médico não invasivo, tendo sido as colheitas feitas pelos treinadores que realizam este comportamento com o animal. Diariamente lidam com estes animais vários treinadores e também, sempre que necessário, membros da equipa Veterinária. As duas principais piscinas que constituem o habitat destes animais ocupam 1958,7 m3 e 2302,6 m3.
No Zoomarine a citologia das amostras do espiráculo é feita rotineiramente e faz parte do plano mensal de Medicina Preventiva. Neste caso, é pedido aos treinadores que realizem as colheitas e as amostras são mantidas em refrigeração até serem observadas ao microscópio ótico.
20 A preparação para observação ao
microscópio ótico é feita com a ajuda de uma ansa descartável esterilizada, com a qual é retirada a amostra e colocada na lâmina, onde é adicionada uma gota do corante azul de metileno (figura 11). Seguidamente coloca-se uma lamela e observa-se na ampliação de 10x, 40x e de 100x com óleo de imersão, que é usado raramente, pois na dúvida é preparada uma nova lâmina e esta é corada com Diff Quick®.
4.1.2 Material utilizado e colheita das amostras
Foram efetuadas duas recolhas de amostras com cerca de dois meses de intervalo. A primeira recolha foi realizada dia 8 de Janeiro de 2012 e a segunda dia 18 de Março do mesmo ano. No primeiro caso, colheram-se duas amostras de cada animal e no segundo apenas uma. Em ambas as ocasiões a par destas foram efetuadas recolhas de duas amostras da água da piscina em garrafas esterilizadas, uma em superfície e outra em profundidade, sendo registada simultaneamente a temperatura da água e do ambiente, bem como a humidade relativa.
O processamento microbiológico das amostras decorreu no Laboratório de Microbiologia do Departamento de Ciências Veterinárias da Universidade de Trás-os-Montes e Alto Douro (UTAD).
A recolha foi efetuada com base na seguinte metodologia:
Fase de preparação da colheita (representada na figura 17):
o Limpeza e desinfeção das mãos do treinador que vai realizar a colheita com clorhexidina sabão 4% (i) e, posteriormente, álcool a 96% (ii);
o O treinador pede ao animal uma expiração, a qual é desperdiçada;
o Secagem da área do espiráculo com papel absorvente (iii). Limpeza e desinfeção da área com clorhexidiina 4% (iv) e secagem posterior com compressa seca esterilizada (v).
Fase de colheita (representada na figura 17):
o O treinador mostra a base da placa de Petri ao animal colocando-a ao lado do olho (vi) e, em seguida, pede as três expirações, aproximando esta do espiráculo (vii). Entre cada expiração este processo repete-se;
Figura 11- Material necessário para a realização de uma citologia com amostras do espiráculo.