Voluntary wheel running: patterns and physiological effects in mice
Texto
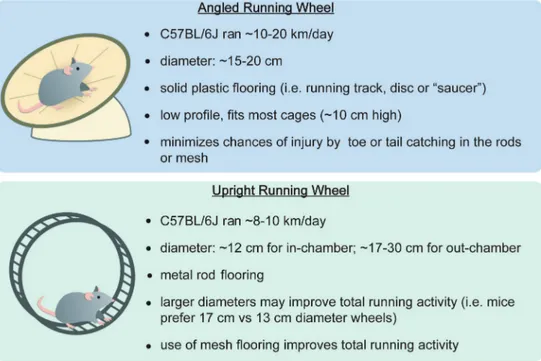
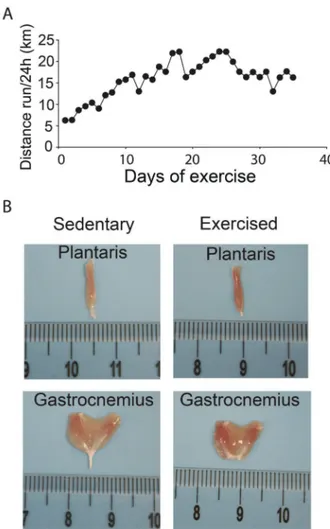
Imagem
Documentos relacionados
Neste trabalho o objetivo central foi a ampliação e adequação do procedimento e programa computacional baseado no programa comercial MSC.PATRAN, para a geração automática de modelos
Além disso, o Facebook também disponibiliza várias ferramentas exclusivas como a criação de eventos, de publici- dade, fornece aos seus utilizadores milhares de jogos que podem
Afinal, se o marido, por qualquer circunstância, não puder assum ir a direção da Família, a lei reconhece à mulher aptidão para ficar com os poderes de chefia, substituição que
Na hepatite B, as enzimas hepáticas têm valores menores tanto para quem toma quanto para os que não tomam café comparados ao vírus C, porém os dados foram estatisticamente
É nesta mudança, abruptamente solicitada e muitas das vezes legislada, que nos vão impondo, neste contexto de sociedades sem emprego; a ordem para a flexibilização como
Quanto ao grupo SB, as participantes que possuíam a mastigação bilateral alternada permaneceram, apesar de a ativação muscular ter aumentado e ter sido possível
Carollia perspicillata had the high- est recapture rate, especially in the banding plots of the riparian forest, suggesting the highest fidelity to the sample area of all
The main goal of the ALERT-ES project (“Sistema de Alerta Sísmica Temprana: Aplicación al Sur de España”) is to study the feasibility of an Earthquake Early Warning System (EEWS)
