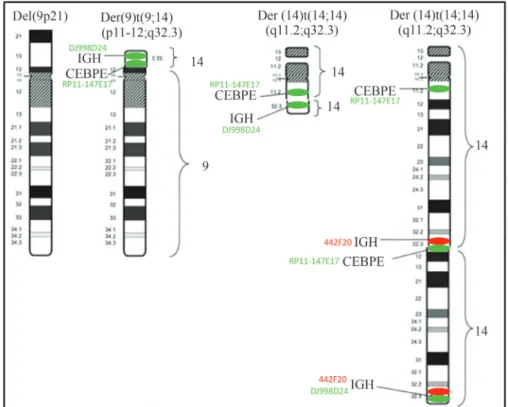
The complex translocation (9;14;14) involving
Texto
Imagem

Documentos relacionados
Qualquer participante da Pajero TR4ER Master em 2014 Qualquer participante das categorias Triton ER e Triton RS 3 primeiros colocados de qualquer categoria em.. campeonatos de rallye
Ele fugiu para Roma, e a irmã mais velha de Cleópatra, Berenice, tornou-se rainha.. Em 55 a.C., Ptolomeu retornou
Pressione o botão para medir a distância “a”, em seguida repita o passo para a medir a distância “b”, e novamente para a medida “c”, o resultado da área “S”
Flutuação populacional de adultos capturados com atrativos florais e lagartas capturadas em rede de varredura e em plantas e posturas das espécies Physocleora
Ao administrador público cabem as medidas de aprimoramento da qualidade da gestão do gasto público. As avaliações do Tribunal de Contas do Estado do Rio de Janeiro a
Os fragmentos florestais podem funcionar como corredores para deslocamento de anuros entre habitats de reprodução e áreas onde realizam outras atividades como
Tratando-se de uma Unidade Curricular de Ensino Prático e Laboratorial, todos os objetivos focados no desenvolvimento de competências (orais, expositivas, de apresentação, de
Candidatos autodeclarados pretos, pardos ou indígenas, com renda familiar bruta per capita igual ou inferior a 1,5 salário mínimo e que tenham cursado integralmente o ensino médio
