PROGRAMA DE PÓS-GRADUAÇÃO EM FISIOTERAPIA
“Efeitos da Clorfeniramina na aprendizagem,
memória e ansiedade de teleósteos submetidos à
ablação telencefálica”
Flávia Roberta Faganello Navega
PROGRAMA DE PÓS-GRADUAÇÃO EM FISIOTERAPIA
“Efeitos da Clorfeniramina na aprendizagem, memória e
ansiedade de teleósteos submetidos à ablação
telencefálica”
Flávia Roberta Faganello Navega
Orientadora: Rosana Mattioli
SÃO CARLOS 2006
Ficha catalográfica elaborada pelo DePT da Biblioteca Comunitária/UFSCar
N323ec
Navega, Flávia Roberta Faganello.
Efeitos da clorfeniramina na aprendizagem, memória e ansiedade de teleósteos submetidos à ablação telencefálica. -- São Carlos : UFSCar, 2006.
94 p.
Tese (Doutorado) -- Universidade Federal de São Carlos, 2006.
1. Medicina experimental. 2. Teleósteos. 3. Aprendizagem e memória. 4. Ansiedade. 5. Ablação telencefálica I. Título.
Estudo I
Clorfeniramina facilita a esquiva inibitória em teleósteos submetidos à ablação
telencefálica
Estudo II
Efeito ansiolítico da Clorfeniramina na esquiva inibitória em peixes dourados
submetidos à ablação telencefálica
Estudo III
Papel da CPA na recuperação funcional de teleósteos submetidos à ablação
telencefálica em novo modelo de condicionamento operante para peixes
Orientadora: Prof
a. Dr
aRosana Mattioli
Apoio Financeiro: CAPESAo Marcelo,
Agradecimento Especial
À profa Dra Rosana Mattioli, por ter possibilitado mais essa etapa de minha formação profissional e por tudo o que me ensinou durante esses anos.
À Deus por tantas oportunidades.
À todos os membros da minha banca, que aceitaram o convite para enriquecer esse trabalho.
Aos professores: Azair Liane M.C. Souza, Carla Christina Medalha, e João Eduardo de Araújo por todas as sugestões dadas em minha qualificação.
Ao meu marido, Marcelo, por todo amor, por toda paciência e por ser tão amigo. Eu amo você.
Aos meus pais que sempre incentivaram meus estudos. Às minhas irmãs e ao meu irmão por serem tão amigos e tão importantes em minha vida.
À minha futura afilhada, Maria Eduarda, por já trazer tanta alegria para toda a família.
Às minhas queridas e velhas amigas Ana Cláudia e Cristiane, por tudo.
Aos meus colegas de laboratório, Luciana, Fernanda, Fabiana, Aline e Miagui, por tornarem esses anos tão felizes.
À Luciana, Fernanda e Fabiana por serem amigas e por me ajudarem tanto na realização deste trabalho. Eu adoro vocês.
Ao Miagui, pela velha amizade, pelo empenho em solucionar problemas e por me ajudar com as ilustrações desse trabalho.
À Karina Gramani, por sua amizade tão especial. Você realmente é uma pessoa incrível.
À Andréa, “minha” aluna, por me agüentar e pela ajuda na realização desse trabalho.
Ao professor Nivaldo Parizotto por me introduzir na vida científica, e por ser uma pessoa maravilhosa.
À professora e amiga Vanessa Monteiro Pedro. Eu realmente te adoro.
À Tereza Fatori pela amizade e por toda ajuda na análise histológica.
À Paulinha por ser uma secretária eficiente e amiga
À todo o pessoal dos laboratórios do Departamento de Fisioterapia, pela amizade e pelos bons momentos que passamos juntos.
À professora Tânia Salvini por disponibilizar o criostato. E aos seus alunos, Sabrina e Tiago, por toda a ajuda e paciência.
À todos os meus professores do Programa de Pós-gradução, por terem me ensinado tanto.
The teleost telencephalon provides a good opportunity for studying the function of the
vertebrate limbic system because it is simple and present homologies to the limbic structure of
higher vertebrates, with hippocampus and amygdala. These animals shows only one area of
projection to the telencephalon (Ekström et al., 1995), which has been well preserved through
CONTEXTUALIZAÇÃO 2
ESTUDO I 4
RESUMO 4
ABSTRACT 5
INTRODUÇÃO 6
MATERIAIS E MÉTODOS 8
ANIMAIS 8
ABLAÇÃO TELENCEFÁLICA 9
ANÁLISE HISTOLÓGICA 10
DROGAS E TRATAMENTO 11
AQUÁRIO EXPERIMENTAL 11
PROCEDIMENTOS 12
ANÁLISE ESTATÍSTICA 12
RESULTADOS 13
DISCUSSÃO 15
DESENVOLVIMENTO A PARTIR DO ESTUDO INICIAL 19
ESTUDO II 21
RESUMO 21
ABSTRACT 22
INTRODUÇÃO 23
MATERIAIS E MÉTODOS 25
ANIMAIS 25
ABLAÇÃO TELENCEFÁLICA 25
ANÁLISE HISTOLÓGICA 26
DROGAS E TRATAMENTOS 26
AQUÁRIO EXPERIMENTAL 27
PROCEDIMENTOS 27
ANÁLISE ESTATÍSTICA 28
RESULTADOS 29
DISCUSSÃO 33
DESENVOLVIMENTO A PARTIR DOS ESTUDOS ANTERIORES 37
ESTUDO III 39
RESUMO 39
ABSTRACT 40
MATERIAIS E MÉTODOS 41
ANIMAIS 41
ABLAÇÃO TELENCEFÁLICA E ANÁLISE HISTOLÓGICA 41
AQUÁRIOS EXPERIMENTAIS 42
PROCEDIMENTOS 43
ANÁLISE ESTATÍSTICA 43
RESULTADOS 44
DISCUSSÃO 46
CONSIDERAÇÕES FINAIS 49
REFERENCIAS BIBLIOGRÁFICAS 51
ANEXOS – ESTUDO I 59
ANEXOS – ESTUDO II 71
Contextualização
Peixes, répteis, aves e mamíferos fazem parte de uma mesma escala evolutiva e
possuem um ancestral comum (NORTHCUTT, 1981; CARROL, 1988; BROGLIO, 2003).
Sendo o peixe o mais antigo nesta escala evolutiva, o conhecimento de sua capacidade
cognitiva e suas bases neurais pode contribuir para o entendimento da evolução cerebral e
comportamental dos vertebrados, incluindo dos mamíferos.
Os teleósteos, classe de peixe com esqueleto ósseo, possuem o telencéfalo bem
desenvolvido com áreas homólogas a áreas cerebrais de vertebrados superiores. A área
dorso-lateral do telencéfalo é considerada homóloga ao hipocampo dos mamíferos
(NIEUWENHUYS e MEEK, 1990; BRAFORD, 1995; NORTHCUTT, 1995; VARGAS et
al., 2000, RODRÍGUEZ et al., 2002) e a área dorso-medial, homóloga a amídala (VARGAS
et al., 2000; PORTAVELLA et al., 2002; RODRÍGUEZ et al., 2002).
Nas últimas décadas estudos farmacológicos têm sido realizados a fim de investigar a
função de neurotransmissores sobre os processos de aprendizagem e memória. Dentre eles,
agonistas e antagonistas histaminérgicos têm sido alvo de muita atenção, uma vez que o
sistema histaminérgico está relacionado a uma série de funções no Sistema Nervoso Central
(SNC), incluindo aprendizagem e memória (HASS e PANULA, 2003)
A atuação da histamina no SNC se dá pela estimulação de receptores pós (H1 e H2) e
pré-sinápticos (H3) (ARRANG et al., 1985). Outro tipo de receptor histaminérgico (H4) foi
identificado, porém sua ação, até agora, é considerada restrita ao sistema imunológico (LIM et
al., 2005).
Os resultados dos estudos relacionando o sistema histaminérgico aos processos de
aprendizagem e memória não são conclusivos. Por exemplo, o bloqueio crônico da enzima
sintetizadora da histamina (HA) induziu tanto a facilitação como a inibição da aquisição da
Antagonistas e agonistas inversos do receptor H3 prejudicaram a aprendizagem de medo
condicionado (PASSANI et al., 2001) e melhoraram o desempenho em tarefa de esquiva
inibitória em ratos adultos (FOX et al., 2002, 2003). Em teleósteos a administração de
clorfeniramina (CPA), um antagonista H1, induziu a aprendizagem de preferência
condicionada por lugar (MATTIOLI et al., 1996), e também facilitou a retenção da memória
em um teste de esquiva inibitória (MEDALHA et al., 2000).
Segundo Blandina e colaboradores (2004) o efeito facilitador ou inibidor da histamina
nos processos de aprendizagem e memória pode estar relacionado, dentre outros fatores, à
estrutura cerebral envolvida e a natureza da tarefa realizada. Desta forma a análise das
conseqüências da ablação telencefálica em teleósteos pode contribuir para entendimento das
funções da histamina na aprendizagem e memória uma vez que, o sistema histaminérgico
desses animais apresenta uma única via ascendente ao telencéfalo (EKSTRÖM et al., 1995).
Diante disso, realizamos três estudos. No primeiro, que teve como objetivo verificar o
papel da CPA na esquiva inibitória em teleósteos submetidos à ablação telencefálica,
sugerimos um provável efeito ansiolítico da CPA. No segundo buscamos esclarecer a hipótese
levantada analisando a semelhança entre o efeito da CPA e uma droga benzodiazepínica na
aprendizagem de esquiva inibitória. No terceiro estudo foi desenvolvido um modelo de
condicionamento operante a fim de verificar a ação da CPA na recuperação funcional de
teleósteos submetidos à ablação telencefálica, e conseqüentemente ajudar no esclarecimento
Estudo I
“Clorfeniramina facilita a esquiva inibitória em teleósteos submetidos
à ablação telencefálica”.
Resumo
O objetivo desse estudo foi verificar o papel da CPA na esquiva inibitória em teleósteos submetidos à ablação telencefálica. Os peixes sofriam ablação telencefálica cinco dias antes de iniciado os procedimentos experimentais. Os procedimentos de esquiva inibitória foram realizados em um aquário retangular dividido em dois compartimentos (branco e preto) em 5 dias. Um dia para a adaptação (os peixes eram adaptados por 10 minutos), três dias para o treino composto por 3 tentativas cada (primeiro dia T1, T2 e T3; segundo dia 2T1, 2T2 e 2T3 e terceiro dia 3T, 3T2 e 3T3) e um dia para teste. Nos dias de treino, os peixes eram colocados em um compartimento branco e após 20 segundos a porta, que separava os dois compartimentos, era aberta. Quando o peixe passava para o compartimento preto, um peso de 45 gramas era solto. Esse procedimento era repetido por três vezes consecutivas. Imediatamente após a terceira tentativa, em cada dia de treino, o peixe recebia um dos tratamentos farmacológicos (salina, 8 ou 16 µg/g de CPA). No dia do teste, o tempo de passagem para o compartimento preto era registrado. Os resultados sugerem que a CPA, principalmente na dose de 8 µg/g, apresentou um efeito facilitatório na memória da tarefa de esquiva inibitória. Nós sugerimos que o peixe submetido à ablação telencefálica foi capaz de aprender a tarefa de esquiva inibitória devido a circuitos locais presentes no mesencéfalo que atua para a geração e elaboração de reações defensivas, e que a CPA, devido a seu efeito ansiolítico, interfere nesses circuitos mesencefálicos.
“Chlorpheniramine facilitates the inhibitory avoidance in teleosts submitted to
telencephalic ablation”
Abstract
The aim of this study was verify the role of CPA in inhibitory avoidance in teleosts submitted to telencephalic ablation. The fish underwent telencephalic ablation five days prior to the experimental procedure. The inhibitory avoidance procedure was performed in a rectangular aquarium divided into two compartments (black and white), in 5 days. One day for adaptation (fish were adapted for 10 min.), three days for training composed of 3 trials each (first day T1, T2 and T3; second day 2T1, 2T2 and 2T3 and third day 3T1, 3T2 and 3T3), and one day for test. On training days, the fish were placed in a white compartment and after 20 seconds the door, that separated the compartments, was opened. When the fish crossed to a black compartment, a 45-gram weight was dropped. This procedure was performed three times in a row. Immediately after the third trial, on each day of training, the fish received one of the pharmacological treatments (saline, 8 or 16µg/g of CPA). On test day, the time for crossing to the black compartment was recorded. The results suggest that CPA, mainly at a dosage of 8 µg/g, had a facilitating effect on memory of the inhibitory avoidance task. We suggest that the fish submitted to telencephalic ablation were able to learn the inhibitory avoidance task due to the local circuits of midbrain that act to generate and elaborate defense reactions, and that CPA, probably due to its anxiolytic-like effect, interferes in these midbrain circuits.
Introdução
É bem conhecido que o sistema límbico esta envolvido no medo-relacionado ao
condicionamento clássico em mamíferos (KAPP et al., 1979; DAVIS, 2000; MEDINA et al.,
2002). Por outro lado, a ablação do telencéfalo dos teleósteos, que é filogeneticamente
relacionada ao sistema límbico de vertebrados superiores, não impediu o condicionamento
autonômico clássico em um procedimento aversivo (OVERMIER e CURNOW, 1969;
MEDINA et al., 2002), o que sugere que estruturas mesencefálicas podem ser capazes de
promover aprendizagem de respostas emocionais condicionadas.
Teleósteos são apropriados para o estudo da histamina como neurotransmissor no
sistema nervoso central (SNC) porque o sistema histaminérgico em peixes é mais restrito e
possui somente uma área de projeção no telencéfalo (EKSTRÖN et al., 1995) que foi bem
preservada durante a evolução (PEITSARO et al., 2003).
Vários estudos sugerem que a HA participa dos processos de aprendizagem e
formação da memória (HASS e PANULA, 2003; CHOICH et al., 2004). Entretanto os
resultados são freqüentemente contraditórios, visto que efeitos facilitatório e inibitório já
foram descritos (BLANDINA et al., 1996; PASSANI et al., 2001; CANGIOLI et al., 2002).
Estudos anteriores mostraram que a histamina apresenta um efeito inibitório em
diferentes tarefas de esquiva inibitória. Eidi e colaboradores (2003) sugeriram que a
administração pós-treino de histamina atenuou a retenção da memória e potencializou o
prejuízo na memória induzido pela escopolamina, em ratos. Em um estudo realizado por
Rubio e colaboradores (2001) o aumento nos níveis de histamina cerebral em ratos prejudicou
a aquisição da resposta de esquiva, ao passo que níveis reduzidos facilitaram essa aquisição.
Em teleósteos, a clorfeniramina (CPA), um antagonista histaminérgico H1, melhorou a
Uma vez que, em alguns estudos, a histamina apresentou um efeito inibitório em
tarefas de aprendizagem e a ablação telencefálica em teleósteos não interrompeu o
condicionamento autonômico clássico usando um procedimento aversivo, o objetivo desse
estudo foi verificar o papel da CPA na esquiva inibitória em teleósteos submetidos à ablação
Materiais e métodos
Animais
Foram utilizados quarenta e seis peixes dourados (Carassius auratus) (figura 1) de
sexo indeterminado pesando de 4 a 8g. Os animais eram mantidos em aquários de 30 litros
(15 peixes por aquário) com filtro e oxigenação constante, a uma temperatura de 20-22º com
ciclo natural de luz.
Os peixes eram alimentados cinco vezes por semana com ração flutuante (Wardly
Corporation, New Jersey, U.S.A.). Um intervalo de duas semanas de aclimatação foi dado
entre a compra do peixe e os procedimentos cirúrgicos. Os animais eram individualmente
identificados por suas características físicas, como cor e tipo de cauda.
Ablação Telencefálica
Os peixes foram anestesiados por imersão em uma solução de 0,8 g/l de tricaína
metasulfonado (TMS)-(3-aminobenzoic acid ethyl ester methasulfonate, Sigma Chemical Co.)
até a atividade motora espontânea e os movimentos branquiais cessarem. Os peixes eram
retirados da solução anestésica, envolvidos em gaze úmida e posicionados adequadamente em
um aparato (figura 2) para a realização dos procedimentos cirúrgicos. Uma solução anestésica
de 0,3 g/l de TMS foi perfundida continuamente pela boca, durante todo o procedimento
cirúrgico, por perfusão pela boca.
A parte dorsal do crânio era removida para a exposição dos lobos telencéfalicos. Os
lobos telencefálicos eram então retirados por aspiração através de uma pipeta conectada a um
sistema de vácuo. Após a ablação do telencéfalo o crânio era fechado com acrílico dental de
secagem rápida (Dental Vipi Ltda, Pirassununga, São Paulo – Brazil). A solução anestésica
era então substituída por água até reiniciarem os movimentos branquiais, e os peixes eram
recolocados nos aquários de manutenção por um período de recuperação de 5 dias antes do
início dos procedimentos experimentais.
Análise Histológica
Após a conclusão dos procedimentos experimentais os animais eram novamente
anestesiados, em seguida decapitados, os cérebros foram removidos dos crânios e
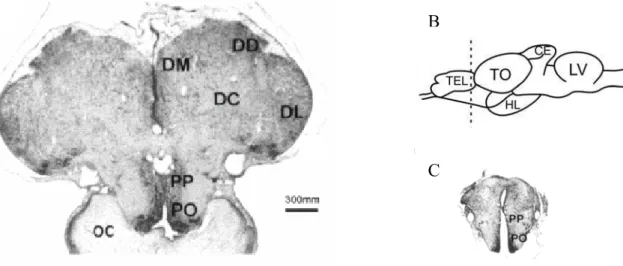
inspecionados para uma avaliação preliminar da ablação (figura 3). Os cérebros eram então
fixados em formalina 10% por pelo menos quatro dias. Depois disso, eram seccionados a uma
espessura de 50 µm usando um criostato. Por meio da análise histológica verificamos que os
cérebros submetidos à ablação exibiram completa remoção do tecido telencefálico ao passo
que as áreas pré-ópticas e tratos ópticos não foram lesados (figura 4).
A
B B
C
Fig 4: Corte transverso do cérebro de teleósteos. A: cérebro normal. C: cérebro após ablação telencefálica. B: Esquema do cérebro mostrando o local de secção. Abreviaturas: TEL: telencéfalo; TO: trato óptico; CE: cerebelo; LV: lobo vagal; HL: hipotálamo lateral, DC: região dorso-central do telencéfalo, DD: região dorso-dorsal; DM: região dorso-medial; DL: região dorsolateral; PP: núcleo pré-óptico periventricular; PO: núcleo pré-óptico. Adaptado de Portavela et al.,2003.
Fig 3: A: foto de um cérebro íntegro de teleósteo. B: foto de um cérebro de teleósteo após ablação telencefálica.
Drogas e tratamento
Clorfeniramina (CPA) sal de maleato (Sigma, MO, USA) foi dissolvida em salina
(0.9%) nas concentrações de 8 e 16 µg/µl. CPA 8 e 16 µg/g ou salina (SAL) foram
administrado intraperitonealmente no volume de 1µl/g. As substâncias foram codificadas e os
códigos não eram conhecidos pelo experimentador.
Os peixes foram divididos em três grupos. Grupo SAL (n=20) composto por animais
tratados com salina, grupo CPA8 (n=12) e grupo CPA16 (n=13) compostos, respectivamente,
por animais que receberam tratamento de CPA nas doses de 8 e 16 µg/µl.
Aquário experimental
Foi utilizado um aquário, de vidro, retangular dividido em dois compartimentos sendo
um branco e o outro preto, delimitados por uma porta do tipo guilhotina. Cada compartimento
possuía dimensões de 15 centímetros de altura, 10 centímetros de largura e 22,5 centímetros
de comprimento (figura 5).
Procedimentos
O experimento foi realizado em 5 dias consecutivos. Um dia de adaptação, três dias de
treino (compostos por 3 tentativas cada: primeiro treino T1, T2 e T3, segundo treino 2T1, 2T2
e 2T3 e terceiro treino 3T1, 3T2 e 3T3) e um dia de teste.
No dia de adaptação os animais eram colocados individualmente no aquário por 10
minutos. Nos dias de treino os animais eram colocados individualmente no compartimento
branco do aquário e após 20 segundos a porta era retirada. Quando o peixe atravessava a linha
divisória entre os compartimentos um peso de 45 gramas era solto a sua frente. O peixe
voltava ou era imediatamente recolocado no compartimento inicial, o procedimento era
repetido por mais 2 vezes. A passagem para o compartimento preto foi definida como o
momento em que a nadadeira dorsal do animal entrava no compartimento preto.
Imediatamente após a terceira tentativa, nos três dias de treino os animais recebiam o
tratamento farmacológico (SAL, CPA8 ou CPA16). Vinte e quatro horas após, os animais
eram testados e o tempo de entrada no compartimento preto era registrado novamente.
A determinação de colocar o animal inicialmente no compartimento branco se deve a
sua preferência natural pelo ambiente escuro (SANTANGELO e MATTIOLI, 1999). Todo
procedimento experimental foi gravado em vídeo-tape.
Análise estatística
Todos os resultados foram inicialmente submetidos ao teste Levene para verificar a
homogeneidade dos dados. Quando apropriado, os dados foram transformados em log10 e
então analisados pela análise de variância (ANOVA) seguida pelo teste de comparações
múltiplas de Dunn.
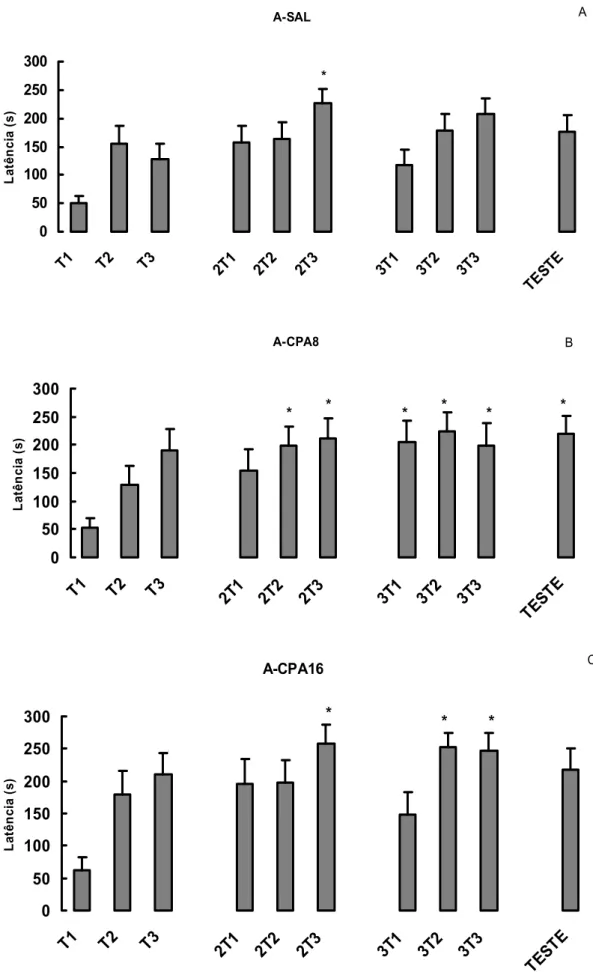
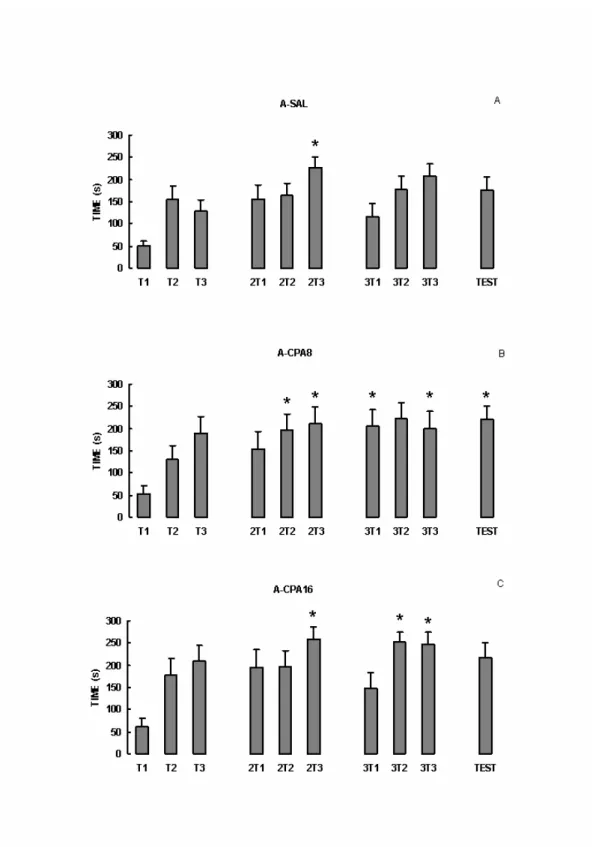
Resultados
A figura 6-A mostra as médias e erro padrão das médias (+EPM) das latências dos
dias de treino e teste do grupo SAL. A latência foi significativamente maior somente na
terceira tentativa do segundo dia de treino (ANOVA: d.f.=9, F=2.19, p=0.0249 e Dunn
p<0.05). Sugerindo que os animais tratados com salina foram capazes de aprender a tarefa,
mas não conseguiram reter a informação aprendida.
Os animais tratados com CPA8 mostraram um aumento nas latências, tentativa após
tentativa, no segundo dia de treino e permaneceu alta nas três tentativas do terceiro dia de
treino e no dia do teste (ANOVA: d.f.=9, F=3.15, p= 0.0022 e Dunn p<0.05) (Figura 6-B). O
grupo CPA16 apresentou maiores latências no segundo e terceiro dia de treino (ANOVA
d.f.=9, F=3.67, p=0.0005 e Dunn p<0.05) (figura 6-C). Esses resultados sugerem que a CPA,
A-CPA16 0 50 100 150 200 250 300
T1 T2 T3 2T1 2T2 2T3 3T1 3T2 3T3
TESTE La tê nc ia ( s ) * * * C A-CPA8 0 50 100 150 200 250 300
T1 T2 T3 2T1 2T2 2T3 3T1 3T2 3T3
TEST E La tê nc ia ( s ) * * * * * * B A-SAL 0 50 100 150 200 250 300
T1 T2 T3 2T1 2T2 2T3 3T1 3T2 3T3
TEST E La tê nc ia ( s ) * A
Discussão
Os resultados desse estudo sugerem que a CPA melhora a retenção da tarefa de
esquiva inibitória em peixes submetidos à ablação telencefálica. Resultados similares foram
encontrados por Medalha e colaboradores (2000) em um estudo realizado em peixes dourado
intactos no mesmo modelo experimental. Em seu estudo o grupo tratado com CPA na dose de
8 µg/g apresentou um aumento significativo na latência para deixar o compartimento de saída.
Assim, nós sugerimos que a ação da CPA no condicionamento de esquiva inibitória ocorre via
estruturas mesencefálicas e/ou diencefálicas.
Fanselow (1981) e Blanchard e colaboradores (1989) propuseram que padrões de
comportamento defensivos são organizados em uma série hierárquica de repostas. Alguns
estudos sugerem que a estimulação de estruturas do teto do mesencéfalo produz alerta,
“freezing” e reações de fuga. Sugerem também que a estimulação elétrica da substância
cinzenta periaquedutal e do colículo inferior induz efeito aversivo, reforça a reposta de fuga
operante e também permite aprendizagem de repostas emocionais condicionadas (JENCK et
al., 1983, 1995; BRANDÃO et al., 1997).
O teto do mesencéfalo apresenta circuitos locais para a geração e elaboração de
reações defensivas, entretanto estruturas cerebrais superiores são necessárias para conseguir o
controle de comportamentos mais complexos relacionados ao medo. Por exemplo, a conexão
do colículo inferior com a amídala atua como um importante filtro para as informações
sensoriais de natureza aversiva (MAISONNETTE et al., 1996). Sabendo-se que a esquiva é
um tipo de comportamento defensivo (MISSLIN, 2003), nós podemos sugerir que os peixes
submetidos à ablação telencefálica são capazes de aprender a tarefa de esquiva inibitória
devido aos circuitos locais do mesencéfalo, e que a CPA facilita a retenção da experiência
Outros estudos sugerem também uma ação do sistema histaminérgico no diencéfalo,
principalmente em tarefas que envolvam componentes emocionais como aversão, estresse e
medo. Ito e colaboradores (1999) submeteram ratos a uma situação de estresse agudo, em que
os animais eram mantidos imóveis em uma gaiola apertada por uma hora. A análise
histoquímica mostrou um aumento nos níveis de histamina no diencéfalo.
Ferretti e colaboradores (1998) relataram que o estresse induz a liberação de histamina
no hipotálamo. Da mesma forma, no estudo de Tasaka e colaboradores (1985) após a
administração de histamina intraventricular, maior radioatividade foi determinada no
hipotálamo. Neste estudo a histamina causou uma inibição dose-dependente na resposta de
esquiva em ratos, e os autores sugeriram que esse efeito inibitório está preferencialmente
ligado ao receptor H1.
Regiões do teto do mesencéfalo contêm substratos para o comportamento defensivo,
aversão e medo (BRANDÃO et al., 2003). Existem boas evidências que a aprendizagem de
esquiva é baseada na aquisição de um estado “antecipatório” de medo em peixes dourados
(OVERMIER e CURNOW, 1969; GALONN, 1972; PORTAVELLA et al., 2003). De acordo
com Brandão e colaboradores (2003) os processos preparatórios para a orientação do perigo,
medo e esquiva são ligados a ansiedade. Assim, o efeito da CPA na memória encontrado em
nosso estudo pode ser também devido a um efeito ansiolítico da CPA mediado pelo
diencéfalo e/ou teto do mesencéfalo.
O envolvimento do sistema histaminérgico no estado de ansiedade de animais já foi
sugerido por alguns autores. Malberg-Aiello e colaboradores (2002) mostraram que
substâncias capazes de aumentar a transmissão histaminérgica reduziram o tempo de
permanência no compartimento claro em camundongos, indicando um provável efeito
ansiogênico. Em um estudo fisiológico foi relatado que a destruição da sub-região E2
rostroventral do núcleo tuberomamilar em ratos, que é o local de origem do sistema
parece estar relacionado à diminuição da atividade histaminérgica causado pela lesão (FRISH
et al., 1998). Em um estudo realizado por Medalha et al. (2003) em peixes dourados, o grupo
tratado com CPA apresentou valores mais baixos no nível de um metabólito da serotonina
(5-HIAA- ácido 5-hidroxihindileacetico) no diencéfalo quando comparado ao grupo tratado com
salina, sugerindo que a CPA pode apresentar um efeito ansiolítico uma vez que a serotonina
está envolvida nos processos de ansiedade (GRAEFF, 2002).
Uma hipótese adicional pode ser a participação do sistema colinérgico. Estudos
clínicos (BEATTY e BUTTERS, 1986; EAGGER et al., 1991; JONES et al., 1992) e
experimentais (DUNNETT et al.,1985) têm mostrado que o sistema colinérgico desempenha
um importante papel na aprendizagem, memória e atenção, e outros estudos sugerem que o
sistema histaminérgico influencia a aprendizagem e memória por modulação da liberação da
acetilcolina (PASSANI et al., 2000; BACCIOTTINI et al., 2001). Em um trabalho realizado
por Eidi et al. (2003) a melhora na memória induzida pela acetilcolina ou nicotina pode ser
prejudicada pela histamina. Em uma análise bioquímica, Miyazaki et al. (1995) sugeriram que
a histidina (precursora da histamina) diminuiu os níveis de acetilcolina no córtex cerebral, no
diencéfalo e mesencéfalo de ratos.
Uma vez que células colinérgicas foram descritas no teto do mesencéfalo de teleósteos
(GONZALEZ et al., 2002) e que o sistema histaminérgico tem origem no diencéfalo
(EKSTRÖN et al., 1995), e regiões do mesencéfalo apresentam altos níveis de histamina em
peixes (e em mamíferos) (ALMEIDA e BEAVEN, 1981), o efeito facilitatório da CPA em
nosso estudo também pode ser devido a uma relação entre os sistemas histaminérgico e
colinérgico no mesencéfalo e em estruturas diencefálicas. Entretanto estudos adicionais, sobre
a relação da histamina/acetilcolina na aprendizagem de esquiva inibitória, devem ser
realizados.
Em conclusão, podemos concluir que os peixes submetidos à ablação telencefálica são
mesencéfalo que atuam para a geração e elaboração de reações defensivas, e que a CPA
Desenvolvimento a partir do estudo inicial
No primeiro estudo verificamos que os animais com ablação telencefálica foram
capazes de aprender o condicionamento de esquiva inibitória, apesar do telencéfalo ser
considerado homólogo a estruturas límbicas do cérebro de vertebrados superiores. Esse fato,
provavelmente se deu devido à capacidade de estruturas mesencefálicas gerarem reações
comportamentais que são primordiais para a sobrevivência da espécie, denominadas reações
defensivas.
As reações defensivas ocorrem frente à situações de perigo, e desencadeiam respostas
incondicionadas de medo geradas no teto mesencefálico (BRANDÃO et al., 2003). Segundo
Misslin (2003) respostas de esquiva fazem parte do repertório de comportamentos defensivos
dos animais assim como fuga, luta, imobilidade, agressão e ajustes fisiológicos em geral.
Essas reações apresentam íntima relação com a ansiedade e estresse (RAU et al., 2005).
Fortes experiências emocionais podem prejudicar tarefas previamente adquiridas, por
isso drogas ansiolíticas, quando administradas após o treino de tarefas com fortes
componentes emocionais, podem auxiliar na retenção da tarefa (DIAMOND et al, 2004). O
que reforça a hipótese de efeito ansiolítico da CPA levantada em nosso primeiro estudo, uma
vez que os animais tratados com essa droga apresentaram melhor desempenho na tarefa
proposta.
Entretanto, Izquierdo e colaboradores (1990) sugerem que algumas tarefas precisam
de certo grau de estresse e ansiedade para serem aprendidas, como é o caso da esquiva
inibitória. Em um estudo realizado por de Souza-Silva e colaboradores (1993) o tratamento
prévio com diazepam prejudicou a retenção da tarefa de esquiva inibitória em ratos.
pré-treino de clonazepam e diazepam, que são drogas ansiolíticas, apresentaram um efeito
prejudicial na aquisição do condicionamento de esquiva inibitória em ratos.
Vários estudos já relacionaram o sistema histaminérgico com a ansiedade, e apontam
principalmente para uma relação diretamente proporcional, ou seja, que maiores
concentrações de histamina seriam responsáveis por maiores índices de ansiedade (OISHI et
al., 1992; FRISH et al., 1998; MALBERG-AIELLO et al., 2002; BONGERS et al., 2003).
Com base nas informações apresentadas verificamos que drogas ansíoliticas podem
apresentar resultados opostos no condicionamento de esquiva inibitória, dependendo do
momento de sua administração. Quando administradas antes do treino apresentam um efeito
inibitório, e quando administradas após o treino podem causar um efeito facilitatório. Diante
do exposto, decidimos realizar um segundo estudo a fim de verificar se há semelhança entre o
efeito do antagonista histaminérgico H1 (CPA) e uma droga benzodiazepínica (Diazepam),
Estudo II
“
Efeito ansiolítico da Clorfeniramina na esquiva inibitória em peixes
dourados submetidos à ablação telencefálica”
Resumo
O objetivo desse estudo foi verificar as conseqüências da ablação telencefálica na aprendizagem de esquiva inibitória em peixes dourados e verificar se há semelhança entre o efeito do antagonista histaminérgico H1 (CPA) e uma droga benzodiazepínica (Diazepam). Os animais eram submetidos á ablação telencefálica ou a lesão fictícia cinco dias antes dos procedimentos experimentais. O procedimento de esquiva inibitória foi realizado em três dias usando um aquário retangular dividido em dois compartimentos (preto e branco) com uma porta central. No primeiro dia os animais eram adaptados por 10 minutos. No segundo e terceiro dias, eles eram injetados com salina (SAL), 16µg/g de Clorfeniramina (CPA), propilenoglicol 40% (PPG) ou µg/g de diazepam (DZP) vinte minutos antes do treino. Então os animais eram colocados no compartimento branco, a porta central era aberta e o tempo para atravessar a linha entre os compartimentos era registrado. Após o peixe passar a linha entre os compartimentos um peso de 45 gramas era solto. Esse procedimento era repetido por três vezes consecutivas. Os grupos submetidos ou não à ablação telencefálica e tratados com salina apresentaram uma diferença entre as sessões de treino; entretanto, os grupos tratados com CPA, PPG ou DZP não apresentaram nenhuma diferença entre eles. Esses resultados sugerem que o tratamento com CPA, PPG ou DZP prejudicou a aquisição do condicionamento de esquiva inibitória nos animais independente da ablação telencefálica. Em conclusão, a ablação telencefálica não retira a capacidade do animal aprender a tarefa de esquiva inibitória, e baseado no fato de que a CPA mostrou efeitos similares ao DZP, nos animais submetidos ou não à ablação telencefálica, nós sugerimos que a CPA apresenta um efeito ansiolítico, mediado pelo diencéfalo e/ou mesencéfalo em peixes dourados.
“Anxiolytic-like effect of Chlorpheniramine in Inhibitory Avoidance in
goldfish submitted to Telencephalic Ablation”
Abstract
The aim of the present study was to verify the consequences of telencephalic ablation on the learning of inhibitory avoidance goldfish and verify the similarity between CPA and benzodiazepnic drug (Diazepam). The animals were submitted to telencephalic ablation or sham operations five days prior to the experimental procedure. The inhibitory avoidance procedure was performed in 3 days using a rectangular aquarium divided into two compartments (black and white) with a central door. On the first day, the animals were adapted for 10 min. On the second and third days, they were injected with Saline (SAL), 16µg/g Chlorpheniramine (CPA), 40% Propylene glycol (PPG) or 1µg/g Diazepam (DZP) twenty min. before training. Then the animals were placed in the white compartment, the central door was opened and the time spent for crossing between compartments was recorded. After the fish crossed the line between the compartments a 45-g weight was dropped. This procedure was performed three times in a row. The groups submitted or not to telencephalic ablation and treated with SAL presented a difference between training sessions; however, the groups treated with CPA, PPG or DZP did not show any differences between them. These results suggest that the treatment with CPA, PPG or DZP impaired the acquisition of inhibitory avoidance conditioning in animals regardless of telecenphalic ablation. In conclusion, telencephalic ablation does not disrupt the animals’ capacity to learn the inhibitory avoidance task, and based on the fact that CPA showed similar effects to those of DZP on the animals submitted or not to telencephalic ablation, we suggest that the CPA presents an anxiolytic-like effect mediated by the diencephalons and/or mesencephalon in goldfish.
Introdução
O telencéfalo dos teleósteos fornece uma boa oportunidade para o estudo da função do
sistema límbico dos vertebrados porque ele é simples e homólogo a estruturas límbicas de
vertebrados superiores (DROOGLEEVER-FOUTUYN, 1961; NORTHCUTT e
BRADFORD, 1980; LOPEZ et al., 2000).
Portavella e colaboradores (2004) estudaram a resposta de esquiva após lesão de áreas
telencefálicas e sugeriram a presença de dois diferentes sistemas de memória em peixes
dourados. Um sistema de memória emocional relacionado à região medial do telencéfalo, e
um sistema de memória espacial e temporal relacionado à região lateral do telencéfalo. Além
disso, os efeitos diferenciais causados pelas lesões telencefálicas são similares àqueles
produzidos por lesão amídalar e hipocampal em mamíferos.
Teleósteos são apropriados para o estudo da Histamina como neurotransmissor no
Sistema Nervoso Central (SNC), uma vez que o sistema histaminérgico em peixes apresenta
somente uma aérea de projeção para o telencéfalo (EKSTRÖN et al., 1995) que foi bem
preservada durante a evolução (PEITSARO et al., 2003).
Desde a descoberta que antihistamínicos clássicos apresentavam uma ação sedativa,
tornou-se claro que a histamina desempenha importante ação no SNC (HASS e PANULA,
2003). Vários estudos sugerem que a histamina está envolvida em muitas funções
neurológicas, como o controle do despertar, atenção, processos sensoriais e formação da
memória (CHOICH et al., 2004).
Os dados disponíveis sobre os efeitos da histamina central no comportamento animal,
especialmente na aprendizagem e memória, ainda são contraditórios. Alguns estudos
descrevem um efeito inibitório da histamina (BEARDSLEY, 1992; SANNERUD et al., 1995;
da histamina na aprendizagem e memória (De ALMEIDA e IZQUIERDO, 1986; KAMEI e
TASAKA, 1993; MEGURO et al., 1995; MIYAZAQUI et al.,1995; COELHO et al., 2001).
Vários estudos também revelaram um papel da histamina no medo e na ansiedade
(OISHI et al., 1986; IMAIZUMI e ONODERA, 1993; FRISCH et al., 1998).
Considerando que o telencéfalo dos teleósteos é homólogo a estruturas límbicas de
vertebrados superiores, que o sistema histaminérgico central está envolvido na aprendizagem,
memória e ansiedade (De ALMEIDA e IZQUIERDO, 1986, MATTIOLI et al., 1998) e sua
maior área de projeção está localizada no telencéfalo, o objetivo deste estudo foi verificar a
relação do sistema histaminérgico na aprendizagem de esquiva inibitória em peixes
submetidos á ablação telencefálica e investigar se há semelhança entre o efeito do antagonista
histaminérgico H1 (CPA) e uma droga benzodiazepínica.
Materiais e métodos
Animais
Cento e cinqüenta e seis peixes dourados (Carassius auratus) (figura 1) de sexo
indeterminado e pesando de 4 a 8 g foram usados. Os animais eram mantidos em um aquário
de 30 litros (15 peixes por aquário) com filtro e oxigenação constante a uma temperatura de
20-22º com ciclo natural de luz.
Eles foram alimentados cinco vezes por semana com ração flutuante (Wardly
Corporation, New Jersey, U.S.A.). Um intervalo de duas semanas de aclimatação foi admitido
entre a compra do peixe e os procedimentos cirúrgicos. Os animais eram individualmente
identificados por suas características físicas, como cor e tipo de cauda.
Ablação Telencefálica
Os peixes foram anestesiados por imersão em uma solução de 0,8 g/l de tricaína
metasulfonado (TMS)-(3-aminobenzoic acid ethyl ester methasulfonate, Sigma Chemical Co.)
até a atividade motora espontânea e os movimentos branquiais cessarem. Os peixes eram
envolvidos em gaze e colocados em um aparato com suportes laterais que estabilizavam o
corpo do animal para a realização dos procedimentos cirúrgicos. Anestesia foi mantida
continuamente, durante todo o procedimento cirúrgico, por perfusão de uma solução 0,3 g/l de
TMS pela boca (figura 2).
A parte dorsal do crânio era removida para a exposição dos lobos telencefálicos. Os
lobos telencefálicos eram então retirados por aspiração através de uma pipeta conectada a um
sistema de vácuo. Após a ablação do telencéfalo o crânio foi fechado com acrílico dental de
fictícia (Sham) eram submetidos aos mesmos procedimentos cirúrgicos exceto pela aspiração
dos lobos telencefálicos. Depois de o crânio ser fechado a solução anestésica era substituída
por água até reiniciarem os movimentos branquiais. Os peixes eram então recolocados nos
aquários de manutenção para um período de recuperação de cinco dias antes do início dos
procedimentos experimentais.
Análise Histológica
Após a conclusão dos experimentos os animais eram decapitados, os cérebros eram
removidos e inspecionados para uma avaliação preliminar da ablação (figura 3). Os cérebros
eram então fixados em formalina 10% por pelo menos quatro dias. Depois disso, eram
seccionados a uma espessura de 50 µm em um criostato. A análise mostrou que os cérebros
submetidos à ablação exibiram completa remoção do tecido telencefálico ao passo que as
áreas pré-ópticas e tratos ópticos não foram lesados (figura 4).
Drogas e tratamentos
Diazepam (DZP) (Roche S/A, São Paulo, Brasil) foi diluído em propilenoglicol 40%
(PPG) na concentração de 1.0 µg/µl. Clorfeniramina (CPA) sal de maleato (Sigma, MO,
USA) foi dissolvida em salina (0.9%) até a concentração de 16 µg/µl. DZP, PPG, CPA ou
salina (SAL) foram administrados intraperitonealmente no volume de 1µl/g. As substâncias
foram codificadas e os códigos não eram conhecidos pelo experimentador, que não sabia que
substâncias eram injetadas em cada animal.
Os peixes foram divididos em oito grupos levando em considereação o procedimento
cirúrgico, submetido à ablação telencefálica (A) ou não (S), e o tratamento farmacológico
Tabela1: grupos experimentais
Grupo N Tratamento
S-Sal Sham 26 Salina
S-PPG Sham 20 Propilenoglicol (40%) S-DZP Sham 20 Diazepam 1µg/g
S-CPA16 Sham 13 CPA 16 µg/g
A-Sal Ablação 20 Salina
A-PPG Ablação 20 Propilenoglicol (40%) A-DZP Ablação 20 Diazepam 1µg/g
A-CPA16 Ablação 17 CPA 16 µg/g
Aquário Experimental
Foi utilizado um aquário de vidro retangular dividido em dois compartimentos sendo
um branco e o outro preto, delimitados por uma porta do tipo guilhotina (figura 5). Cada
compartimento possuía dimensões de 15 centímetros de altura, 10 centímetros de largura e
22,5 centímetros de comprimento.
Procedimentos
O experimento foi realizado em três dias consecutivos, sendo um dia de adaptação e
dois dias de treino. No dia de adaptação os animais eram colocados individualmente no
aquário por 10 minutos.
Os dias de treino eram compostos por três tentativas cada sendo primeiro dia de treino:
T1, T2 e T3 e segundo dia de treino: 2T1, 2T2 e 2T3. Nos dois dias de treino, vinte minutos
antes de iniciado os procedimentos experimentais os animais eram injetados com SAL, CPA,
DZP ou PPG. Então, os animais eram colocados individualmente no compartimento branco
do aquário e após 20 segundos a porta era retirada. Quando o peixe atravessava a linha
divisória entre os compartimentos um peso de 45 gramas era solto a sua frente. O peixe
repetido por mais 2 vezes. A passagem para o compartimento preto foi definida como o
momento em que a nadadeira dorsal do animal passava a linha divisória entre os
compartimentos. Todo procedimento experimental era gravado em vídeo - tape.
A determinação de colocar o animal inicialmente no compartimento branco se deve a
sua preferência natural pelo ambiente escuro (SANTANGELO e MATTIOLI, 1999).
A atividade locomotora do animal na primeira tentativa do primeiro dia de treino (T1)
foi registrada para verificar se as drogas interferiam na locomoção dos animais. Para isso, o
compartimento branco do aquário foi dividido em três partes iguais, e o número de
cruzamentos entre as partes foi contado. O número de cruzamentos dividido pela latência foi
calculado para todos os grupos e utilizado como indicador da atividade locomotora.
Análise estatística
Todos os resultados foram inicialmente submetidos ao teste Levene para verificar a
homogeneidade dos dados. Quando apropriado, os dados foram transformados em log10 e
então analisados pela análise de variância (ANOVA) seguida pelo teste de comparações
múltiplas de Student-Newman-Keuls (SNK).
Resultados
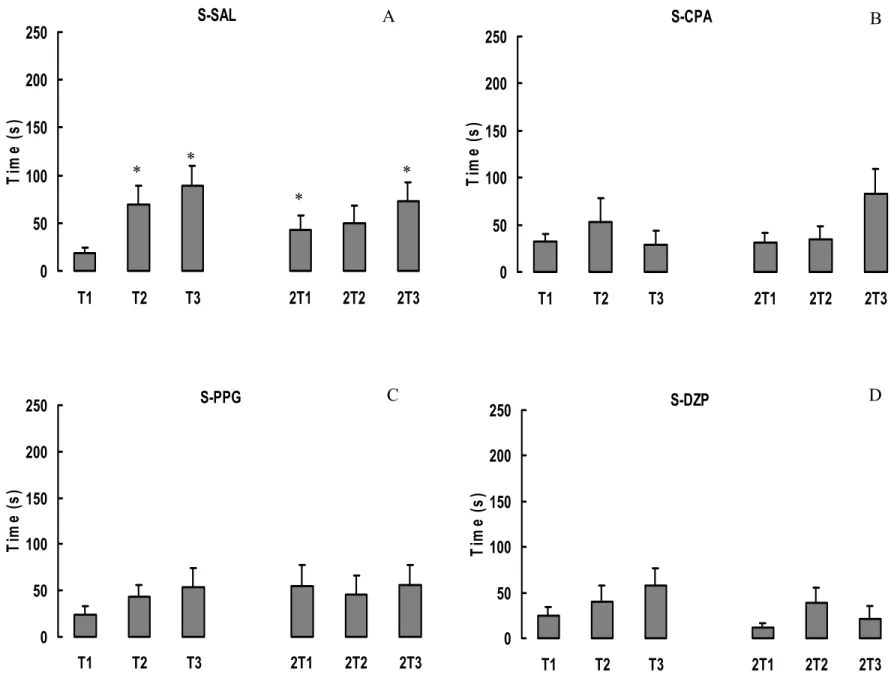
A figura 7 mostra as médias e erro-padrão das médias (+SEM) das latências dos dias
de treino dos grupos Sham. O grupo S-SAL apresentou um aumento significativo nos valores
de latências entre cada tentativa nos dois dias de treino (ANOVA d.f=5; p=0,001 e SNK
p<0,05). Os grupos S-CPA, S-PPG e S-DZP não apresentaram diferença significativa nos
valores de latência em nenhum dia de treino (ANOVA d.f.=5: p=0,23, p=0,73 e p=0,12
respectivamente).
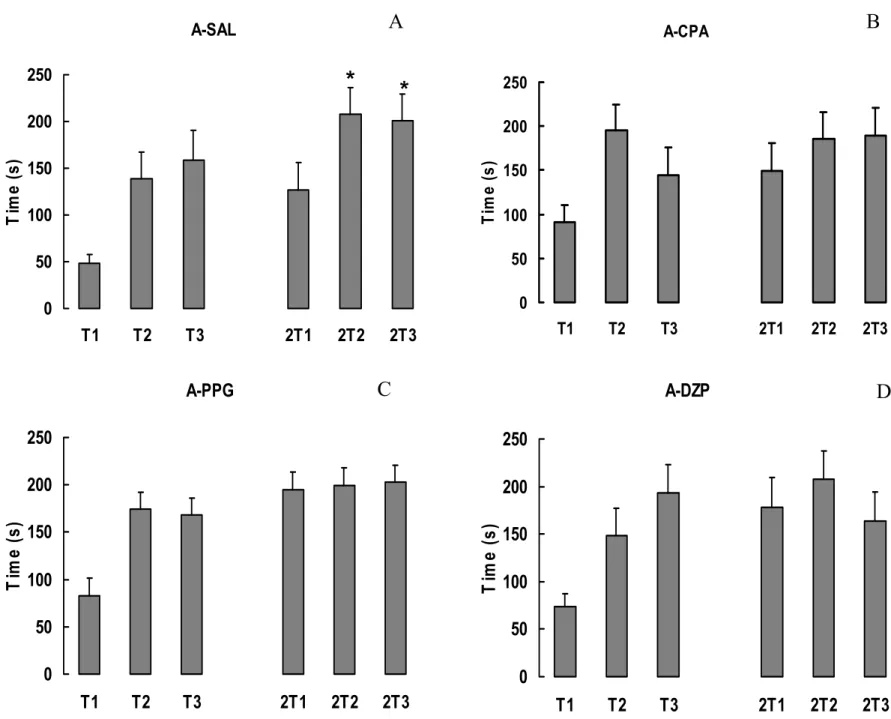
A figura 8 mostra as médias e erro-padrão das médias (+SEM) das latências dos dias
de treino dos grupos com ablação telencefálica. O grupo A-SAL apresentou um aumento
significativo nos valores de latências entre cada tentativa nos dias de treino (ANOVA d.f=5;
p=0,0018 e SNK p<0,05). Os grupos A-CPA, A-PPG e A-DZP não mostraram diferença
significativa nos valores de latência em nenhum dia de treino (ANOVA d.f.=5: p=0,29,
p=0,09 e p=0,28 respectivamente).
Esses resultados sugerem que o tratamento com CPA, PPG ou DZP prejudicou a
aquisição do condicionamento de esquiva inibitória nos animais submetidos ou não a ablação
telencefálica, já que tanto os animais dos grupos Sham como dos grupos submetidos à ablação
telencefálica apresentaram prejuízos semelhantes após tratamento com essas drogas.
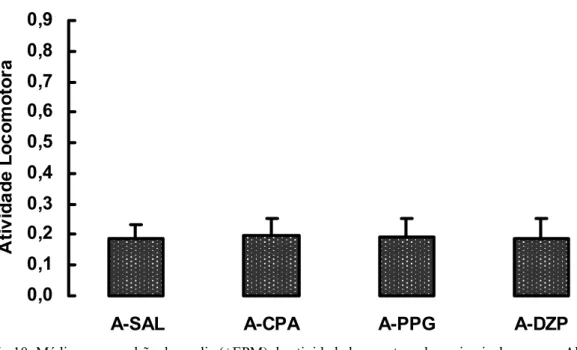
A figura 9 mostra as médias e erro-padrão das médias (+SEM) da atividade
locomotora dos grupos Sham (ANOVA: d.f.=3, p=0,29) e a figura 10 mostra as médias e
erro-padrão das médias (+SEM) da atividade locomotora dos grupos submetidos à ablação
telencefálica (ANOVA: d.f.=3, p=0,99). Com base nesses dados sugerimos que os diferentes
tratamentos farmacológicos não interferiram na locomoção dos animais. Entretanto, quando
realizada uma comparação da atividade locomotora dos animais dos grupos sham e ablação,
verificamos que a retirada do telencéfalo causa uma diminuição na locomoção dos animais
S-CPA 0 50 100 150 200 250
T1 T2 T3 2T1 2T2 2T3
Ti m e ( s ) S-DZP 0 50 100 150 200 250
T1 T2 T3 2T1 2T2 2T3
Ti m e ( s ) S-SAL 0 50 100 150 200 250
T1 T2 T3 2T1 2T2 2T3
Ti m e ( s ) S-PPG 0 50 100 150 200 250
T1 T2 T3 2T1 2T2 2T3
Ti m e ( s )
A B
C D
* *
*
*
A-CPA 0 50 100 150 200 250
T1 T2 T3 2T1 2T2 2T3
Ti m e ( s ) A-DZP 0 50 100 150 200 250
T1 T2 T3 2T1 2T2 2T3
Ti m e ( s ) A-SAL 0 50 100 150 200 250
T1 T2 T3 2T1 2T2 2T3
Ti m e ( s ) A-PPG 0 50 100 150 200 250
T1 T2 T3 2T1 2T2 2T3
Ti m e ( s )
A
B
C
D
*
*
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9
S-SAL S-CPA S-PPG S-DZP
A tiv ida d e loc o m o to ra 0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9
A-SAL A-CPA A-PPG A-DZP
A tiv id a d e L o c o m o to ra
Fig 9: Média e erro padrão da media (+EPM) da atividade locomotora dos animais dos grupos Sham (ANOVA: p=0,29).
Discussão
Tanto os animais submetidos à ablação telencefálica como os animais submetidos à
lesão fictícia (grupos SHAM) foram capazes de aprender a tarefa de esquiva inibitória quando
tratados com salina, sugerindo que a ablação telencefálica não impede o animal aprender essa
tarefa.
Em um estudo realizado por Werka e Zielinski (1998) ratos submetidos à lesão na
amídala (que corresponde à parte dorso-medial do telencéfalo de teleósteo) foram capazes de
aprender a reposta de esquiva, com leve atraso. Da mesma forma, lesão hipocampal (que
corresponde à parte dorso-lateral do telencéfalo de teleósteo) em ratos não afetou
significativamente a aquisição e retenção da esquiva inibitória (GIONET et al.,1991). Por
outro lado, a completa ablação do telencéfalo prejudicou a aprendizagem de esquiva em
peixes dourados em outros estudos (OVERMIER e STARKMAN, 1974; FLOOD et al., 1976;
FLOOD e OVERMIER, 1981; OVERMIER e HOLLIS, 1990).
Segundo Brandão e colaboradores (2003) estruturas mesencefálicas apresentam
circuitos locais que são capazes de gerar comportamentos defensivos, como no caso a
resposta de esquiva. Sendo assim, provavelmente, graças à esses circuitos locais no
mesencéfalo, os peixes submetidos ablação telencéfalica foram capazes de aprender a tarefa
proposta em nosso estudo.
Foi verificado, neste estudo, que a ablação telencefálica causou uma diminuição na
atividade locomotora dos animais. Resultados semelhantes já foram citados por outros
autores, que sugeriram uma diminuição da atividade espontânea após ablação telencefálica em
peixes (JANZEN, 1933; HALE, 1956). A aquisição da tarefa, dos grupos sham e ablação,
foram analisadas separadamente para evitar uma possível interpretação equivocada dos dados,
como nenhuma diferença na atividade locomotora foi encontrada entre os tratamentos
farmacológicos, tantos nos animais submetidos à ablação telencefálica ou à lesão fictícia, a
aprendizagem de esquiva inibitória não foi devido a uma mudança na atividade locomotora,
mas realmente à aquisição da tarefa.
O comportamento dos animais tratados com PPG e CPA foi similar aos animais
tratados com DZP, uma droga ansiolítica clássica, sugerindo um provável efeito ansiolítico do
PPG e da CPA. Em nosso estudo, DZP bem como o PPG e a CPA prejudicaram a aquisição
da esquiva inibitória nos animais submetidos ou não a ablação telencefálica.
Estudos anteriores sugeriram um efeito-ansiolítico do PPG. Em 1998, Lin e
colaboradores reportaram que o PPG reduziu a ansiedade em um teste de labirinto elevado em
ratos. Lourenço da Silva e Elisabetsky (2001) também demonstraram uma interferência
ansiolítica do PPG, na concentração de 30%, em um teste “hole board” em ratos. Sendo
assim, uma vez que os animais tratados com PPG apresentaram comportamento semelhante
aos animais que receberam DZP, e que outros autores já verificaram efeitos ansiolítcos do
PPG, principalmente quando administrado em altas concentrações, como em nosso estudo,
podemos sugerir que o prejuízo na aquisição do condicionamento de esquiva inibitória foi
causado pela redução da ansiedade nos animais também pela administração prévia do PPG.
O envolvimento do sistema histaminérgico no estado de ansiedade do animal não está
ainda bem esclarecido. Oishi e colaboradores (1986) reportaram que o DZP diminui a taxa de
renovação da histamina em ratos, sugerindo uma participação do sistema histaminérgico
cerebral na ansiedade. Malberg-Aiello e colaboradores (2002) mostraram que substâncias
capazes de aumentar a transmissão da histamina reduzem o tempo de permanência no
compartimento claro, indicando um provável efeito ansiogênico.
Resultados de um estudo fisiológico mostraram que a destruição da subregião E-2 na
histaminérgicos, causou efeito ansiolítico em um teste de labirinto elevado, esse efeito foi
provavelmente devido à redução da atividade histaminérgica induzida pela lesão (FRISH et
al., 1998). Em uma situação de estresse agudo, foi verificado um aumento no nível de
histamina no hipotálamo (MAZURKIEWICZ-KWILECK e TAU, 1978) bem como no
diencéfalo, núcleo acumbente e estriado de rato (ITO et al., 1999, 2000). Ito e colaboradores
(1999) sugeriram que o aumento no nível de histamina no diencéfalo após estresse agudo
pode representar um aumento na síntese de histamina, uma vez que o diencéfalo é o principal
local de síntese desse neurotransmissor. Depois de sintetizada a histamina é transportada por
axônios até o núcleo acumbente e estriado onde é metabolizada.
Em peixes, a relação entre o sistema histaminérgico e a ansiedade já foi sugerida em
estudos comportamentais (MATTIOLI et al., 1998; MEDALHA et al., 2000, COELHO et al.,
2001, PEITSARO et al., 2003). Recentemente, Medalha e colaboradores (2003), em um
estudo realizado em peixes dourados usando o método de cromatografia líquida (HPLC),
sugeriram um efeito ansiolítico da CPA. Nesse estudo a CPA não alterou os níveis de
monoaminas e seus metabólicos no telencéfalo, mas os resultados referentes ao diencéfalo
indicaram que o grupo tratado com CPA apresentou baixos níveis de HIAA (ácido
5-hidroxihindoleacético) comparados ao grupo tratado com salina, sugerindo que a CPA pode
ter um efeito ansiolítico uma vez que a serotonina está envolvida nos processo de ansiedade
(GRAEFF, 2002). É importante salientar que esses dados neuroquímicos foram encontrados
nas áreas diencefalicas, que de acordo com Ekströn et al. (1995), é a origem do sistema
histaminérgico em peixes.
Em conclusão, a ablação telencefálica não interrompeu a capacidade do animal em
aprender a tarefa de esquiva inibitória, e, baseado no fato da CPA ter mostrado efeitos
sugerimos que a CPA apresenta um efeito ansiolítico mediado pelo diencéfalo e/ou
Desenvolvimento a partir dos estudos anteriores
Nos dois estudos anteriores foi verificado que os animais submetidos à ablação
telencefálica são capazes de aprender o condicionamento de esquiva inibitória proposto
devido ao fato de estruturas cerebrais primitivas serem capazes de gerar reações defensivas. E
a CPA parece atuar através dessas estruturas causando um efeito ansiolítico. Entretanto,
estruturas cerebrais superiores são necessárias para o controle de comportamentos mais
complexos, como por exemplo para o controle de diferentes formas de aprendizagem e
memória.
Dessa maneira é de grande importância a realização de estudos que verifiquem as
funções do telencéfalo de teleósteos, uma vez que essa estrutura contém processos
anatômicos que são considerados precursores do sistema límbico de vertebrados superiores
(Droogleever Fortuyn, 1961).
O telencéfalo dos teleósteos desempenha um importante papel em diferentes tipos de
aprendizagem. López e colaboradores (2000) relacionaram o telencéfalo com a capacidade de
memória espacial em teleósteos. Ohnishi (1989) sugeriu que o telencéfalo é fundamental na
integração de eventos neurais que ocorrem em estruturas extratelencefálicas para a
aprendizagem instrumental e Portavella e colaboradores (2002) mostraram a relação do
telencéfalo à aprendizagem emocional e temporal.
Sabendo-se que os processos neurais que ocorrem durante a aprendizagem e formação
da memória, denominados de plasticidade neural, são semelhantes em aspectos
neuroquímicos e morfológicos aos que ocorrem durante a recuperação funcional após lesão do
SNC (COTMAN et al., 1981; COTMAN e LYNCH, 1989), o terceiro estudo teve como
a fim de observar a aprendizagem após ablação telencefálica em Carassius auratus e,
Estudo III
“Papel da CPA na aprendizagem de teleósteos submetidos à ablação
telencefálica em novo modelo de condicionamento operante para peixes”
Resumo
O objetivo desse estudo foi desenvolver um modelo de condicionamento operante para estudar a aprendizagem após ablação telencefálica em peixes dourados e verificar o papel da CPA nessa função. Os animais eram submetidos á ablação telencefálica ou a lesão fictícia cinco dias antes do início dos procedimentos experimentais. Seis aquários retangulares, com uma barreira opaca delimitando a área de alimentação, foram usados. O condicionamento operante foi realizado em dez dias consecutivos. A barreira opaca era retirada e, após 30 segundos, o alimento era oferecido por um tubo de acrílico colocado dentro da área de alimentação. O tempo que o animal levava para entrar na área de alimentação era registrado. Foram realizados quatro grupos experimentais sendo, grupo Ablação (n=14): animais submetidos à ablação telencefálica, grupo Sham (n=10): animais submetidos à lesão ficitícia, grupos Abla-Sal (n=12): animais submetidos à ablação telencefálica e tratados com salina e grupo Abla-CPA (n=11): animais submetidos à ablação telencefálica e tratados com 16 µg/g de CPA. O tratamento farmacológico era administrado em dias alternados, dez minutos após o término dos procedimentos experimentais. Foi observada uma diminuição nos valores de latência no grupo Sham a partir do segundo dia até o final do experimento (ANOVA p< 0,001, Student Newman Keuls P<0,01). O grupo ablação apresentou uma leve diminuição nas latências, mas nenhuma diferença significativa foi observada (ANOVA p=0,237), indicando um prejuízo após ablação telencefálica. O grupo Abla-sal mostrou diminuição nas latências a partir do sexto dia até o final (ANOVA p<0,001, Student Newman Keuls P<0,01). O grupo A-CPA não apresentou diminuição nos valores de latência (ANOVA p=0,607). Os resultados sugerem que esse modelo é adequado para da aprendizagem após ablação telencefálica em teleósteos, e a CPA apresenta um provável efeito ansiolítico que prejudicou o desempenho do animal.
“
The Role of CPA in the learning of teleosts submitted to telencephalic
ablation in a new model of fish operant conditioning
”
Abstract
The aim of the present study was to develop an instrumental conditioning model to study the learning after telencephalic ablation in goldfish, and also verify the role of CPA in this function. The animals were submitted to telencephalic ablation or sham surgery five days prior to the beginning of the experimental procedure. Six rectangular aquariums with an opaque barrier delimitating the feeding area, were employed. The operant conditioning was realized during 10 consecutive days. The conditioning was consisted of the retreating of the opaque barrier followed by the food offering (30 seconds after) through an acrylic tube inserted at the feeding area. The time spent by the animal to enter at the feeding area was recorded. Four experimental groups were carried out. Ablation group (n=14): the animals were submitted to the telencephalic ablation; Sham group (n=10): the animals were submitted to the sham operation. Abla-Sal (n=12): animals submitted to the telencephalic ablation and treated whit saline and Abla-CPA (n=11) group: animals submitted to the telencephalic ablation and received 16 µg/g of CPA. The treatment was administered in alternate days, ten minutes after the end of the experimental procedure. A decrease in latencies for the sham animals was observed from the second day until the end of experiment (ANOVA p< 0,001, Student Newman Keuls P<0,01). The ablation group presented a slight decrease in latencies, but not statistically significant differences were observed (ANOVA p=0,237), indicating an impairment after telencephalic ablation. The Abla-Sal group showed a statistically significant decrease in latencies from sixth day until the end (ANOVA p<0,001, Student Newman Keuls P<0,01). The Abla-CPA group did not present any decrease in latency values (ANOVA p=0,607). Results suggest that this model is suitable for the study of learning recovery after telencephalic ablation in teleost and that CPA presents an anxiolytic-like effect that impaired animal performance.
Materiais e Métodos
Animais
Foram utilizados 47 peixes da espécie Carassius auratus de sexo indeterminado e peso
entre 4 e 10 gramas, provenientes do comércio local. Foram mantidos em aquários de 30 litros
(15 peixes por aquário) com filtro e oxigenação constante até o dia da cirúrgia de ablação
telencefálica. Nesses aquários eram alimentados cinco vezes por semana com ração flutuante
(Wardly Corporation, New Jersey, U.S.A.) entre 8 e 9 horas da manhã.
Após a ablação telencefálica os peixes eram colocados individualmente nos aquários
experimentais, onde eram alimentados, com clara de ovo cozida, até um dia antes do início do
experimento.
Ablação Telencefálica e Análise histológica
Todos os procedimentos cirúrgicos, bem como a análise histológica, foram
semelhantes aos realizados nos estudos anteriores.
Drogas e tratamentos
Clorfeniramina (CPA) sal de maleato (Sigma, MO, USA) foi dissolvida em salina
(0.9%) até a concentração de 16 µg/µl. CPA ou salina (SAL) foram administrado
intraperitonealmente no volume de 1µl/g. As substâncias foram codificadas e os códigos não
eram conhecidos pelo experimentador.
Os peixes foram divididos em 4 grupos: considerando o procedimento cirúrgico, Sham
Tabela 2: grupos experimentais
Grupo Cirurgia N Tratamento
Sham Sham 10 -
Ablação Ablação 14 -
Abla-Sal Ablação 12 Salina Abla-CPA Ablação 11 CPA 16 µg/g
Aquários Experimentais
Foram utilizados 6 aquários de vidro transparente de formato retangular, (16cm de
largura, 35cm de comprimento e 20cm de altura) que possuíam em uma das arestas um tubo
de acrílico transparente de 16 cm de comprimento que direcionava a queda do alimento (clara
de ovo) até o fundo do aquário. A 8 cm da parede lateral, que continha o tubo de fornecimento
de alimento, foi colocada uma divisória de plástico rígido e opaco com 24 cm de altura, e 15
cm de comprimento para delimitar a área de alimentação (Figura 11).
Procedimentos
O condicionamento operante foi realizado em 10 dias consecutivos e teve inicio 5 dias
após a cirurgia de ablação telencefálica.
A divisória, que delimita a área de alimentação, era retirada e após 30 segundos o
alimento era oferecido pelo tubo. A partir do momento da retirada da divisória o tempo de
entrada na área de alimentação era cronometrado. Ao entrar na área de alimentação o animal
podia se alimentar por 2 minutos, ao final desse período a divisória era recolocada e o
excedente de alimento retirado. Caso o animal não entrasse na área de alimentação em 10
minutos a divisória era recolocada e o alimento retirado. Para os animais dos grupos tratados,
10 minutos após a recolocação da divisória os animais recebiam SAL ou CPA no primeiro,
terceiro, quinto, sétimo e nono dias de experimento.
Uma câmera de vídeo (JVC VHS Super Digital Signal Processing) era colocada em
um tripé a uma distância de 83 cm da parede frontal dos aquários que foram mantidos na
mesma posição durante todos os dias do experimento.
Análise estatística
Todos os resultados foram inicialmente submetidos ao teste Levene para verificar a
homogeneidade dos dados. Os dados foram analisados pela análise de variância (ANOVA)
seguida pelo teste de comparações múltiplas de Student-Newman-Keuls (SNK).
Resultados
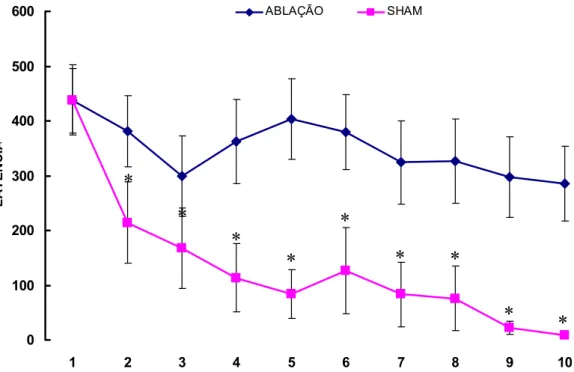
A figura 12 apresenta as médias (+ EPM) das latências de entrada na área de
alimentação dos animais dos grupos Ablação e Sham
No grupo Sham foi verificado uma diminuição da latência dia a dia de treino, sendo
significativa a partir do segundo dia (ANOVA p<0,001, Student Newman Keuls p<0,01). Já
os animais do grupo Ablação não apresentaram queda nos valores de latência nos dias de
treino (ANOVA p=0,237).
0 100 200 300 400 500 600
1 2 3 4 5 6 7 8 9 10
LA
TÊ
N
C
IA
ABLAÇÃO SHAM
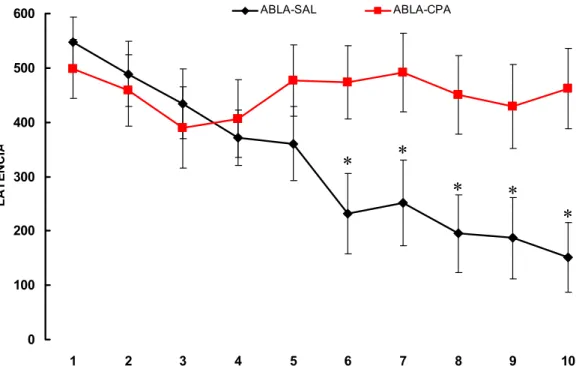
A figura 13 apresenta as médias (+ EPM) das latências de entrada na área de
alimentação dos animais dos grupos Abla-Sal e Abla-CPA.
Os animais do grupo Abla-Sal apresentaram uma diminuição na latência significativa
(ANOVA p<0,001, Student Newman Keuls P<0,01) a partir do sexto dia de treino. No do
grupo Abla-CPA não houve diminuição nos valores de latência (ANOVA p=0,607).
*
*
*
*
*
*
*
*
*
0 100 200 300 400 500 600
1 2 3 4 5 6 7 8 9 10
L
AT
Ê
NCI
A
ABLA-SAL ABLA-CPA
*
*
*
*
*
Discussão
Os animais do grupo Sham apresentaram uma diminuição nos valores de latência
significativa a partir do segundo dia de treino, mostrando que foram capazes de associar a
retirada da divisória com o fornecimento do alimento. Já os animais do grupo Ablação
mostraram uma leve queda nos valores de latência durante os dias de treino, porém está queda
não foi significativa, sugerindo que a ablação telencefálica acarretou prejuízo na
aprendizagem do condicionamento operante proposto.
Vários estudos demonstraram prejuízos causados pela ablação telencefálica em
teleósteos em diferentes tipos de aprendizagem. No estudo realizado por López e
colaboradores (2000) animais submetidos à ablação telencefálica tiveram sucesso em tarefas
espaciais constantes, mas severas restrições nas tarefas que envolviam reversões espaciais. No
estudo de Peeke e Gordon (1981) teleósteos submetidos à ablação telencefálica, apresentaram
um grande atraso na aquisição e não foram capazes de reter a tarefa de comportamento
exploratório proposto.
Considerando que os animais submetidos à ablação telencefálica apresentaram grande
prejuízo na tarefa proposta, a utilização desse modelo experimental se torna viável para o
estudo de drogas que possam atuar nos processos de aprendizagem e memória bem como nos
processos de recuperação funcional de teleósteos submetidos à ablação telencefálica.
Nos animais do grupo Abla-Sal foi observada uma diminuição nos valores de latência
dia após dia de treino, sendo essa diminuição estatisticamente significativa a partir do sexto
dia do experimento. Indicando que quando injetados com salina, mesmo animais submetidos à
ablação telencefálica, conseguiram desempenhar a tarefa.
Kaneto (1997) sugere que os processos envolvidos na formação da memória, como a