Diversity and aspects of the ecology of social wasps (Vespidae, Polistinae) in Central Amazonian “terra firme” forest
Texto
Imagem

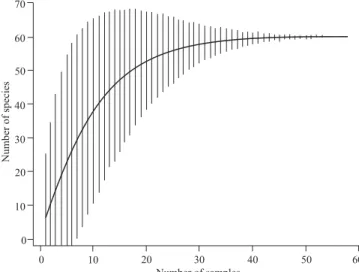

Documentos relacionados
Ousasse apontar algumas hipóteses para a solução desse problema público a partir do exposto dos autores usados como base para fundamentação teórica, da análise dos dados
competindo já perdeu todos os mapas das festas. A participação em uma festa conseguimos com um corpo leve e desarmado.. Com a referida estrutura, trata- se, pois,
Neste trabalho será utilizada uma temperatura de deposição de 300 °C, para obter durezas máximas das camadas individuais, bem como será realizada uma caracterização mais
Conjuntamente, a dupla de artistas concebeu o Grupo Teatro de Dança de São Paulo (anos mais tarde, seria chamado de Célia Gouvêa Grupo de Dança, do qual fizemos
O presente capítulo sustenta-se em quatro pesquisas realizadas no período de 2012 a 2019: Experiência de Integração Ensino e Serviço: um estudo do PET Saúde a partir
Parede anterior (bordo posterior do masséter e do ramo ascendente do maxilar inferior, pterigoideu interno, porção posterior da aponevrose interpterigoideia e
Para validar o manual, os juízes especialistas tiveram como critério de inclusão ser profissionais da saúde com experiência de cinco anos na gestão de serviços de terapia
O negócio proposto é a criação de uma empresa de Assessoria em Comércio Internacional, com foco, inicialmente, em Regime Tributário de Drawback. Uma empresa de
