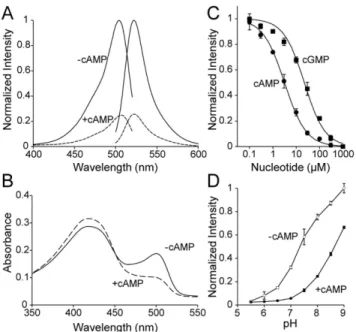
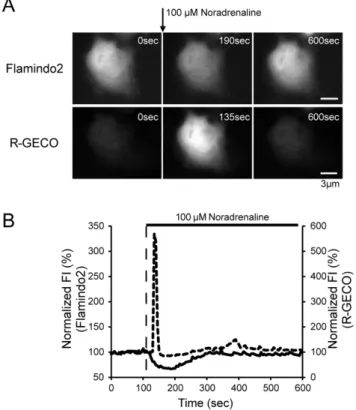
Genetically-encoded yellow fluorescent cAMP indicator with an expanded dynamic range for dual-color imaging.
Texto
Imagem
Documentos relacionados
We showed that intrathecal delivery of IL-6 immediately after spinal cord injury elevated cAMP level in the sensorimotor cortex and increased the expression of growth-associated
During travel between hosts, the genome of a virus is well protected by the viral capsid and/or envelope. After binding specifically to target cells, the virus particles enter cells
Conclusions/Significance: Bioluminescent and fluorescent optical imaging was combined with X-ray and m CT imaging to provide noninvasive and longitudinal measurements of the
For this purpose, we treated ARO (colon cancer), NPA (melanoma) and WRO (follicular thyroid carcinoma) cells with different concentrations of 8-Cl-cAMP or the PKA I-selective
phenomenon occurs in numerous cell types, as consequence of GsPCR activation and increment in intracellular cAMP levels, in this review, we will discuss the impact of cAMP efflux
Evaluation of dynamic sentinel lymph node biopsy in patients with squamous cell carcinoma of the pênis and palpable inguinal nodes.. Intraoperative probes and
In the pre~cnt paper we have made use of a series of mutants affected either in cAMP synthesis or in the activity of cAMP-dependent protein kinase and we provide evidence
Given that cAMP pathway controls GSH levels we tested whether insulin sensitivity decreases after intraportal (ipv) administration of a cAMP analog (DBcAMP), and investigated
