983
ANALYSIS OF PATHWAYS AND NETWORKS INFLUENCING THE DIFFERENTIAL EXPRESSION OF GENES IN CORONARY ARTERY DISEASE
HUI LI, XIAOLAN ZHONG, CHAOMIN LI, LIJING PENG, WEI LIU, MINGXIA YE and BUXIONG TUO
Department of Cardiology, No.451 Hospital of PLA, Xi’an 710054, China Corresponding author: [email protected]
Abstract – Coronary artery disease (CAD) is the leading cause of death worldwide. Microarray analysis is a practical ap-proach to study gene transcription changes that may relect signatures that underlie the pathogenesis of CAD. Using gene expression proile data from the Gene Expression Omnibus database, we identiied diferentially expressed genes that can contribute to the pathology of CAD. Further pathway and network analyses were also implemented to identify pathways and hub genes related to the disease. We observed 466 downregulated and 560 upregulated genes. he ribosome pathway was the most signiicantly over-represented pathway with diferentially expressed genes. Over 35% of the genes in this pathway were downregulated. Hub genes in the network, such as IL7R, FYN, CALM1 ESR1 and PLCG1, may play crucial roles in the pathogenesis of CAD. Our results facilitate the identiication of molecular mechanisms that underlie CAD.
Key words: Coronary artery disease; gene expression; pathway; network
INTRODUCTION
As the most common type of heart disease, coronary artery disease (CAD) is the leading cause of death worldwide (homas, et al., 1988). Most patients show no symptoms or signs of CAD for decades dur-ing progression of the disease. For most patients, the irst onset of symptoms is a sudden heart attack. While considerable eforts have been devoted to the prevention and treatment of CAD, morbidity and mortality remain high. Investigation of the under-lying molecular mechanism may accelerate further investigations into new strategies of noninvasive di-agnosis and treatment.
Microarray analysis is a practical approach to study gene transcription changes that may relect sig-natures underlying CAD pathogenesis. Distinct gene expression patterns in atherosclerotic diseases have been proposed in previous gene expression studies
(Hiltunen, et al., 2002; Nanni, et al., 2006; Randi, et al., 2003; Seo, et al., 2004). Diferent pathways, such as those involved in the inlammatory process and cell cycle control, have been consistently identiied in CAD patients (Cagnin, et al., 2009; Sluimer, et al., 2007), suggesting that “molecule groups” instead of a single or few proteins should be explored while in-vestigating CAD pathogenesis. Most proteins func-tion through their interacfunc-tion with other proteins in certain biological pathways. herefore, analysis from the point of view of pathway and network would pro-vide a better understanding of the mechanisms un-derlying CAD progression. However, most previous eforts have focused on individual genes.
acquired. Protein-protein interaction (PPI) network analysis was also carried out to identify key genes among the diferentially expressed genes.
MATERIALS AND METHODS
Microarray data
he transcription proile GSE12288 was downloaded from the GEO database (http://www.ncbi.nlm.nih. gov/geo/). his series represents the gene expression proile for 110 CAD patients and 112 controls. he data set was based on the GPL96 platform: [HG-U133A] Afymetrix Human Genome U133A Array.
Identiication of diferentially expressed genes
Raw intensity values from raw data iles were nor-malized using Robust Multi-array Analysis (RMA) (Irizarry, et al., 2003) as follows: i) background noise was neutralized by using model-based background correction; ii) expression values were normalized using quantile normalization; iii) the expression value for each probe was generated using an itera-tive median polishing procedure. he log2-trans-formed RMA values for each subject were stored for further identiication of signiicantly diferen-tially expressed genes. Correlation analysis with the Benjamini and Hochberg multiple test correction (Benjamini and Hochberg, 1995) was carried out to detect diferentially expressed genes. he thresh-old was set as 0.01. Up- or downregulation of each diferentially expressed probe was expressed by fold change. All the above procedures were carried out using R sotware (v2.15.1) with BioConductor, limma packages (3.12.1) and libraries (Smyth, et al., 2005).
Pathway enrichment analysis
Selected probes were annotated according to the sim-ple omnibus format in text (SOFT) iles. All genes were then annotated based on the KEGG pathways database (http://www.genome.jp/kegg/) (Kanehisa and Goto, 2000). Enrichment analysis was carried
out using the hypergeometric distribution test to identify pathways enriched with diferentially ex-pressed genes.
Network analysis
Protein-protein interaction (PPI) is critical for bio-logical processes (Stelzl, et al., 2005) since most proteins function through interactions with other proteins. To illustrate the relationship between the diferentially expressed genes, a network was con-structed by using Cytoscape (V 2.8.3, http://www. cytoscape.org/) (Shannon, et al., 2003) and the Na-tional Center for Biotechnology Information (NCBI) database (http://tp.ncbi.nlm.nih.gov/gene/GeneR-IF/, 2013-2-25).
RESULTS
We detected 1 026 diferentially expressed genes of which 466 genes correlated negatively and 560 genes correlated positively with CAD.
Of all of the well-characterized human genes in the array, 5 298 genes were mapped to various path-ways, including 447 diferentially expressed genes. he top 10 pathways enriched with dysregulated genes (Table 1), include two signaling pathways: the calcium signaling pathway (hsa04020) and the adi-pocytokine signaling pathway (hsa04920). he ribos-ome pathway (hsa03010) was the most signiicantly enriched with diferentially expressed genes. Among the 119 genes in the ribosome pathway, 45 genes were negatively correlated with CAD. he vascu-lar smooth muscle contraction pathway (hsa04270) and the dilated cardiomyopathy pathway (hsa05414) were also signiicantly dysregulated.
Table 1. Pathways with dif erentially expressed genes.
KEGG_ID Pathway Description P-value
03010 Ribosome 4.37E-21
04114 Oocyte meiosis 8.18E-04
04740 Olfactory transduction 4.44E-03
00982 Drug metabolism-cytochrome P450 6.42E-03
04020 Calcium signaling pathway 1.51E-02
04920 Adipocytokine signaling pathway 2.09E-02
03040 Spliceosome 2.67E-02
04270 Vascular smooth muscle contraction 3.39E-02
03018 RNA degradation 3.50E-02
05414 Dilated cardiomyopathy (DCM) 4.43E-02
Table 2. h e top i ve molecules in the network constructed by dif erentially expressed genes.
Gene Implications in CAD or other heart disorders (PubMed ID) Degree
IL7R Heart disease (20379172) 19
FYN Coronary artery vasculopathy (18336251) 17
CALM1 Coronary artery disease (17569884) 16
ESR1 Coronary artery disease (23471591, 16612467, 16551651) 15
PLCG1 No known association with heart disorders 13
UBA
UBAP22L RPSPS3333
RR
R
RPL233
RPL23
RPL23A TN TNPO11
R
RPL122
XNA XNA PLX PLXNALXNA22
EIF
EIF5AA
TTT TRHRR
RP
RPL55
CC CC CCNB CCN1 HBE HBE HBEG HBEF HGGFF
CDC2 CDC27 ANP32 ANP32B WB WBP55 LT
LTBP33
EPB41L EPB EPB41L EPB42 TGFB TGFB2 FKBP FKBP2 DH DH DHX DHX DHX99 FN FNBPNBP FNBP4444 TAN TAN SPT SPTANTAN11
PD PD PDE PD PDEDDDD EPB41L
EPB41L3 RRRRRRRPPPL299
AR AR ARRB AR1 ILF3 RP RP RPS2
RPSPS20000 ME METT
ELA
EL
ELAVLLAVL11 RNPA RNPA HNR HNRNPARNPA33 LE LE LEP LERR BA
BA
BAG
BAG BA BAG333333333333
RRR
R
RRPPL188
RR
R
RPPL22222
RANBP RANBP2 NR NR NRT NRNN DV DVL DV22 PD PD PDE4 PD PDE PDE4
PDEDDDD IL7 IL7 IL7 IL7 IL7 IL7 IL7 IL7
IL7RRRR IL6IL6STT
ANKRD ANK ANKRD ANK6 HTR HTR1
HTR1FF PLCPLPLPLCPLCGPLCPLPLCGPLCGPLCGPLPLPLPLCGLCGLCG1111111111CNN31111111111111111CNN3CNN3CNN3CNNCNNCNNCNN3CNN3CNN3CNN3CNN3CNN3CNN SO
SO SO
SO
SO SOCSCSCSCS3333
P P P P P P P PTTTTTTPNPNPN1PN1PN1PN1PN1PN1PN111111 AC
ACP
ACP11 CA CAV22 RE RETT SH2D2 SH2 SH2 SH2 SH2 SH2D2 SH2D SH2 SH2 SH2D2
SH2DAA PL PLC PLC PLC PLC PLC FYYYNNNNNN KP
KPNAPNA11 N
BMX BMX BMX BMX BMX BMX BMX BMX S S
SH3H3BPBPBPBPBPBP22 PTP PT
PTPRRCCCCCCCC
ITKK X39 DDX DDX DDX3 DDX DDX39BFF
RP
RPL77
RSL1D
RSL1D1 TBN
TBN
SPT SPTBNTBN1 HNRHNHNRNPAHNRNPARNPARNPA0
P PRRRPF40PFF40A PT
PTMAAAA BM
BMI11
NOTOTCHCHCHCH22 SR
SRRTTTT
SRR SRR ENP ENP CEN CENPENPBB HIST1H3 HIST1H3C ITG ITGAMM CENP
CENPENPOO CECECECENPCECEAA R
R
RGGS7777
PIN PIN PIN PIN44444 NPH NPH NPH HNR HNR HNR HNR HNR HNR HNR HNRNPHRNPH HNR HNR 33 RPS
RPS4XXXX
HS HSPA888888
YW YW YW YW YW YW YW YW YW YW YWHA YWH YW YW YW YW YW YWH YWHQQQQQQ
BU BU BUB BU1111
RP RP RP RPS1 RPS1 RPS44 BU
BUB3333 TTTTT FA99
MYO10 MYO10 AR
AR
ARIH
AR2222 DNAJB1 DNAJB1 DNAJB1 DNAJB1 DNAJB1 DNAJB1 DNAJB1 DNAJB1 RPS2 RPS28 OG OG OGG
OG1111 CHGB CHGB CHGB CHGB CHGB HM
HMOXOX2
E E E E E EEF1GGGG
DCD DCD PDC PDCDDCD55 UP8 UP8 UP8 UP8 NU NU NUP8
NUP8UP85555
ITG ITGB11
F1A
EEF1A
EEF1AF1AF1A111
MYOC MYOC MYOC MYOC TM4M4SFSF1
CDC25 CDC25C SPE SPE HSP HSPE HSPE11 T
TPT11
ITG ITGA8888 S27 S27 RPS RPS27S27AA FN1 NTS SORT SOORT SORTORT SO11
ME MEF2CC ICAM1 ICAM1 ICAM1 ICAM1 ICAM1 GG GG GGA
GG1111 MMMMEP1AAAA
C3A C3A SLC SLC SLC3AC3A SLC2 L3A L3A COL COL COL3A COL3A11 CA
CALRR
TFPI MMP1 MMP12 LHC LHCGGR ITGA2 ITGA2B RSS RSS PRS PRSSRSS3
TN TN TNCC
TH THBS11
MFAP MFAP5 FB FBLN11
T TSHRR GA GALNTT6 L4A L4A COL COL4A COL4A6 SPATA SPA SPATA SPA2 TP TP TP6 TP TP633 RX RXRBB
RCA RCA SMA SM SMARCARCA4 DUT RU RUNX1 RB RBL
RBL2222222
SS
SS188
SN SNW11 WWTR WWTR1 RA RARGG PP
PPARDD
CE
CEP35350
KAT
KAT6BB
RA RARBB
LATAA SITSIT11CSNCSNCSNCSNKK1EE S S
S
SNRNRPPFF S
S
S
SAFAFBB
P PRRKKCHHHHHHHH
PDLIM PDLIM7 MRPL2 MR MRPL20 H H H H H UBE2 UB H UBE2 H H UBE2 H I PRMT PRMT2 WT1 A A A A A A A ARNRNTTTT
PML SA SATB SATB1 LR1 LR1 LR1 LR1 LR1 POL POL POL POLR1OLR1 POLR1LR1LR1 POLBBBB MEB MEB GME GME GMEB GMEB2222 FFF F
FHITT
GS GS GS GS GSK3 GSK GSAAAA
CB CB CBX CB44 KCNK1 KCNK KCNK1 KCNK US US USO
US1111111 NPY NPY1RR H H H HEY11 GOSR GO GOSR GOS2 SAR SAR SAR
SARTT3
GATAATA GATAATA11 EXOSC EXOSC7
GO
GO
GO GOLLGAAAA22 EXOSC
EXOSC8
LS LSM77
LS
LSM22
CA CALM33
AK AK AK AK AK AK AK AK AK AKAP AK AKA AK AK99 M MEED2D2D2D244
MMS1 MMMS1
MMS1
MMS9 AKA AKAP133
3 3 3 3 3 3 AK AK ESR ESR
ESR111
EEEE
E
E EIF3LL
M
MAFF
LS
LSM55
SE SE
SENP
SE66
UBE
UBE3
UBE3AA
TL
TLK22
AURK AURKAURKURKRKRKA
DEAF DEEAF DEAF DEA1 M M M
MST44
ABLIM
ABLIM1
HH
HSPBB1 RAB RAB2AA
ACTG
ACTG1 TF
TFPTT
ARHRHGEFGEF2 PBRM1 PBRM1 VDR TA TAL11 HIST1H3 HIST1H3A HMT HMT EHM EHMTHMT2 PE PE PEX1 PE44 PE PEX13333 ID4 EXOC EXOC2 AIR
AIREE
PE PEX77
CC
CCT6AA
D DAPKAPK1 MM3 MM3 TOM TO TOMM3MM34
K2A K2A K2A CSN CSN CSN CSN CSN CSN CSNK2AK2AK2A CSN CSN CSN CSN CSN 22
AD ADD1111
R
RA R R LAA
EIF2AK EIF2AK1
RP
RPS3PS3AAAAA ST
STIP11
TRPV TRP TRPV TRPV TRPV TRPV1 DNR DNRNR EDN EDN EDN EDN EDNR EDNR
EDNDNRDNRDNRBBBB
CAV CAV CA CAV1 CNN1 CNN1 CNN1 CNN1 ST ST STRN STR44
H
HSPSPSPSPSP909090AAAAAAAAAA1
CDK11B CDK11B FGFR FG FGFR22 TR TRPC333 PR PR PRKCRKC PRKCRKC
PRKQQQQQQQQQQ TCF TCF33
GR GRK44RKRK
IKB
IKB
IKBK
IKBK IKBK
IKBKBB CA CALM CALM111111
YW YW YW YWHAEEE
NEUROD NEU NEUROD NEUR1 ID3 SK SKP22 TNNFAIPFAIP3 FKBP FKBP8 PAPOL PAPOLA HS HS HSPH
HSPH111 HSP90B HSP90B1 RCCCC XR XRRCC XRCC
XRRCC11
CS CSN CSN333333 F1B F1B EEF
EEF1BF1B22 ULK2 ULK2 UNC5C UNC5C HSPA HSPA9 P1R P1R PPP PPP1R PPP1R2
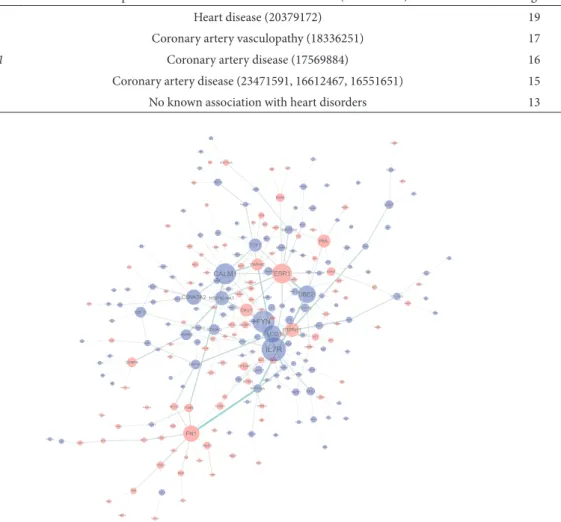
Fig. 1. Interaction network of dif erentially expressed genes. Only genes with more than two direct or indirect links are shown. Genes
DISCUSSION
he pathophysiology of CAD is highly complex. Here we carried out expression proile analysis to identify diferentially expressed genes in CAD patients and healthy controls. Pathway and interaction network analyses were also carried out to explore the under-lying mechanism.
Pathway analysis revealed an over-representa-tion of diferentially expressed genes in the calcium-signaling pathway (hsa04020). his is consistent with prior knowledge, since Ca2+ plays important roles in
the regulation of contraction and intracellular sig-naling, which is crucial for the normal function of healthy heart (Cartwright, et al., 2011). Signiicant dysregulation of another two pathways, the vascular smooth muscle contraction pathway and the dilated cardiomyopathy pathway, also implicated the inter-ruption of normal heart function in CAD patients. he ribosome pathway was most signiicantly over-represented with diferentially expressed genes. Over 35% of the genes in this pathway were downregu-lated. No previous report has proposed the relation-ship of the ribosome pathway and CAD. However, ribosomal RNA expression was revealed to be highly variable in CAD patients (Fong et al, 2013). Moreo-ver, in Drosophila melanogaster, cardiomyopathy is associated with ribosomal protein gene haploinsuf-iciency (Casad, et al., 2011). Further investigation of this pathway on CAD is warranted.
In the constructed network, IL7R showed the highest degree (Fig. 1, Table 2). he protein encoded by the IL7R is a receptor of IL-7, which has been as-sociated with inlammatory processes (Mobini, et al., 2009). Inlammation has been shown to play cru-cial pathologic roles in atherosclerosis CAD (Libby,, 2002). Previous gene expression analysis has also re-ported the downregulation of IL7R in heart disease patients (Lukk, et al., 2010). Our investigation fur-ther highlights the importance of IL7R in the patho-genesis of CAD.
FYN was a hub gene with the second highest de-gree (Fig. 1, Table 2). he protein encoded by this
gene is a membrane-associated tyrosine kinase that has been involved in the control of cell growth. Dif-ferential expression of cell cycle genes in CAD pa-tients has been consistently reported (Cagnin, et al., 2009; Ijas, et al., 2007; Mohler, et al., 2008). he in-volvement of FYN in the pathogenesis of coronary artery vasculopathy through its activation of the T-cell receptor was observed. Our investigation con-irms the relationship between FYN and CAD.
CALM1 encodes a member of the
calcium-binding protein family. Our pathway analysis has highlighted the involvement of the calcium-signal-ing pathway in the pathogenesis of CAD. Previous studies have indicated that a genetic variability of
CALM1 was associated with CAD (Luke, et al., 2009;
Luke, et al., 2007). Herein we present evidence for the involvement of this gene in CAD. Another hub gene is ESR1, which encodes the estrogen receptor. Polymorphism of this gene has been associated with CAD in various populations (Almeida and Hutz, 2006; Wei, et al., 2013; Lawlor, et al., 2006). Our in-vestigation further conirms the involvement of ESR1
in CAD. PLCG1 was also identiied as a hub gene. No previous report of the relationship between CAD
and PLCG1 has been proposed. Further investigation
should be carried out to conirm its implication in the pathogenesis of CAD.
In summary, using gene expression proile data from the GEO database we identiied diferentially expressed genes that may contribute to the pathology of CAD. Signiicantly increased representations of dysregulated genes in the ribosome pathway and cal-cium signaling pathway were identiied. Hub genes in the network, such as IL7R, FYN, CALM1 ESR1 and
PLCG1,could play crucial roles in the pathogenesis
of CAD. Our results facilitate the identiication of the molecular mechanism underlying CAD.
REFERENCES
Benjamini, Y. and Hochberg, Y. (1995). Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. Journal of the Royal Statistical Society Se-ries B (Methodological). 57, 289-300.
Cagnin, S., Biscuola, M., Patuzzo, C., Trabetti, E., Pasquali, A., Laveder, P., Faggian, G., Iafrancesco, M., Mazzucco, A., Pig-natti, P.F. and Lanfranchi, G. (2009). Reconstruction and functional analysis of altered molecular pathways in hu-man atherosclerotic arteries. BMC genomics. 10, 13.
Cartwright, E.J., Mohamed, T., Oceandy, D. and Neyses, L. (2011). Calcium signaling dysfunction in heart disease. BioFac-tors. 37, 175-181.
Casad, M.E., Abraham, D., Kim, I.M., Frangakis, S., Dong, B., Lin, N., Wolf, M.J. and Rockman, H.A. (2011). Cardiomyo-pathy is associated with ribosomal protein gene haplo-insuiciency in Drosophila melanogaster. Genetics. 189, 861-870.
Fong, S.W., Mohamed, M.S., Few, L.L., Too, W.C., Khoo, B.Y., Yu-sof, Z., Rahman, A.R. and Yvonne-Tee, G.B. (2013). Identi-ication of endogenous control genes for gene expression studies in peripheral blood of patients with coronary ar-tery disease. Molekuliarnaia biologiia. 47, 218-223.
Hiltunen, M.O., Tuomisto, T.T., Niemi, M., Brasen, J.H., Rissanen, T.T., Toronen, P., Vajanto, I. and Yla-Herttuala, S. (2002). Changes in gene expression in atherosclerotic plaques an-alyzed using DNA array. Atherosclerosis. 165, 23-32.
Ijas, P., Nuotio, K., Saksi, J., Soinne, L., Saimanen, E., Karjalain-en-Lindsberg, M.L., Salonen, O., Sarna, S., Tuimala, J., Ko-vanen, P.T., Kaste, M. and Lindsberg, P.J. (2007). Microar-ray analysis reveals overexpression of CD163 and HO-1 in symptomatic carotid plaques. Arteriosclerosis, thrombosis, and vascular biology. 27, 154-160.
Irizarry, R.A., Hobbs, B., Collin, F., Beazer-Barclay, Y.D., Anto-nellis, K.J., Scherf, U. and Speed, T.P. (2003). Exploration, normalization, and summaries of high density oligonucle-otide array probe level data. Biostatistics. 4, 249-264.
Kanehisa, M. and Goto, S. (2000). KEGG: kyoto encyclopedia of genes and genomes. Nucleic acids research. 28, 27-30.
Lawlor, D.A., Timpson, N., Ebrahim, S., Day, I.N. and Smith, G.D. (2006). he association of oestrogen receptor alpha-haplotypes with cardiovascular risk factors in the British Women’s Heart and Health Study. European heart journal.
27, 1597-1604.
Libby, P. (2002). Inlammation in atherosclerosis. Nature. 420, 868-874.
Luke, M.M., Kane, J.P., Liu, D.M., Rowland, C.M., Shifman, D., Cassano, J., Catanese, J.J., Pullinger, C.R., Leong, D.U., Arellano, A.R., Tong, C.H., Movsesyan, I., Naya-Vigne, J., Noordhof, C., Feric, N.T., Malloy, M.J., Topol, E.J.,
Koschin-sky, M.L., Devlin, J.J. and Ellis, S.G. (2007). A polymor-phism in the protease-like domain of apolipoprotein(a) is associated with severe coronary artery disease. Arterior, hromb. Vasc. Biol. 27, 2030-2036.
Luke, M.M., O’Meara, E.S., Rowland, C.M., Shifman, D., Bare, L.A., Arellano, A.R., Longstreth, W.T. Jr., Lumley, T., Rice, K., Tracy, R.P., Devlin, J.J. and Psaty, B.M. (2009). Gene variants associated with ischemic stroke: the cardiovascu-lar health study. Stroke; 40, 363-368.
Lukk, M., Kapushesky, M., Nikkila, J., Parkinson, H., Goncalves, A., Huber, W., Ukkonen, E. and Brazma, A. (2010). A glob-al map of human gene expression. Nature biotechnology.
28, 322-324.
Mobini, R., Andersson, B.A., Erjefalt, J., Hahn-Zoric, M., Langs-ton, M.A., Perkins, A.D., Cardell, L.O. and Benson, M.
(2009). A module-based analytical strategy to identify novel disease-associated genes shows an inhibitory role for interleukin 7 Receptor in allergic inlammation. BMC systems biology. 3, 19.
Mohler, E.R. 3rd, Sarov-Blat, L., Shi, Y., Hamamdzic, D., Zalewski, A., Macphee, C., Llano, R., Pelchovitz, D., Mainigi, S.K., Os-man, H., HallOs-man, T., Steplewski, K., Gertz, Z., Lu, M.M. and Wilensky, R.L. (2008). Site-speciic atherogenic gene expression correlates with subsequent variable lesion de-velopment in coronary and peripheral vasculature. Arte-riosclerosis, thrombosis, and vascular biology. 28, 850-855.
Nanni, L., Romualdi, C., Maseri, A. and Lanfranchi, G. (2006). Diferential gene expression proiling in genetic and mul-tifactorial cardiovascular diseases. Molec. Cell. Cardiol. 41, 934-948.
Randi, A.M., Biguzzi, E., Falciani, F., Merlini, P., Blakemore, S., Bramucci, E., Lucreziotti, S., Lennon, M., Faioni, E.M., Ar-dissino, D. and Mannucci, P.M. (2003). Identiication of diferentially expressed genes in coronary atherosclerotic plaques from patients with stable or unstable angina by cDNA array analysis. Arterior, hromb. Vasc. Biol. : JTH.
1, 829-835.
Seo, D., Wang, T., Dressman, H., Herderick, E.E., Iversen, E.S., Dong, C., Vata, K., Milano, C.A., Rigat, F., Pittman, J., Nev-ins, J.R., West, M. and Goldschmidt-Clermont, P.J. (2004). Gene expression phenotypes of atherosclerosis. Arterio-sclerosis, thrombosis, and vascular biology. 24, 1922-1927.
Shannon, P., Markiel, A., Ozier, O., Baliga, N.S., Wang, J.T., Ramage, D., Amin, N., Schwikowski, B. and Ideker, T.
(2003). Cytoscape: a sotware environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498-2504.
Sluimer, J.C., Kisters, N., Cleutjens, K.B., Volger, O.L., Horrevoets, A.J., van den Akker, L.H., Bijnens, A.P. and Daemen, M.J.
atherosclerotic plaques from autopsy and surgery. Physiol.l Genom. 30, 335-341.
Smyth, G.K., Michaud, J. and Scott, H.S. (2005). Use of within-array replicate spots for assessing diferential expression in microarray experiments. Bioinformatics. 21, 2067-2075.
Stelzl, U., Worm, U., Lalowski, M., Haenig, C., Brembeck, F.H., Goehler, H., Stroedicke, M., Zenkner, M., Schoenherr, A., Koeppen, S., Timm, J., Mintzlaf, S., Abraham, C., Bock, N., Kietzmann, S., Goedde, A., Toksoz, E,. Droege, A., Krobitsch, S., Korn, B., Birchmeier, W., Lehrach, H. and Wanker, E.E.
(2005). A human protein-protein interaction network: a resource for annotating the proteome. Cell. 122, 957-968.
homas, A.C., Knapman, P.A., Krikler, D.M. and Davies, M.J. (1988). Community study of the causes of “natural” sud-den death. BMJ. 297, 1453-1456.