Aspectos dimensionais de sementes de Dimorphandra mollis para estudo da
variabilidade entre populações de plantas
Dimensional aspects of Dimorphandra mollis seeds to study variability between
plant populations
DOI:10.34117/bjdv6n8-134
Recebimento dos originais: 12/07/2020 Aceitação para publicação: 12/08/2020
Jaqueline da Silva Santos
Mestre em Recursos Naturais pela Universidade Estadual de Mato Grosso do Sul (UEMS). Rodovia que liga Dourados-Itahúm, KM 12 Cidade Universitária 79804-970, Dourados - MS, Brasil
Montcharles da Silva Pontes
Mestre em Recursos Naturais pela Universidade Estadual de Mato Grosso do Sul (UEMS). Rodovia que liga Dourados-Itahúm, KM 12 Cidade Universitária 79804-970, Dourados - MS, Brasil
Ivanilza Moreira de Andrade
Doutorado em Botânica pela Universidade Estadual de Feira de Santana. Professora efetiva da Universidade Federal do Piauí (UFPI), Av. São Sebastião, 2819, São Benedito 64202-020, Parnaíba
- PI, Brasil
Etenaldo Felipe Santiago
Doutorado em Ciências Biológicas (Biologia Vegetal) pela Universidade Estadual Paulista Júlio de Mesquita Filho (UNESP). Professor efetivo da Universidade Estadual de Mato Grosso do Sul (UEMS), Rodovia que liga Dourados-Itahúm, KM 12 Cidade Universitária 79804-970, Dourados -
MS, Brasil
RESUMO
A morfometria de sementes possui importância taxonômica, filogenética e ecológica, sobretudo quando se trata de árvores nativas do Cerrado. Neste estudo, foram discutidas as potencias relações entre os caracteres dimensionais das sementes e o ambiente de ocorrência em populações de plantas de Dimorphandra mollis Benth., de um cerrado sensu stricto (Css) e uma floresta estacional semidecidual (FES). Foram determinadas medidas de comprimento, diâmetro e espessura das sementes, calculou-se também os diâmetros médios; geométrico (DMG), equivalente (DME), aritmético (DMA), área superficial (As), relação de aspecto (Ra), e esfericidade (Ø). As sementes do
Css apresentaram menor comprimento (média 10,68 mm) e menor diâmetro (média 5,35 mm), enquanto que, as sementes da FES exibiram maior comprimento (média 12,68 mm) e maior diâmetro (média 7,86 mm), com diferença significativa de 5% (p<0,05) entre os locais de coleta. A correlação de Pearson foi mais eficiente para distinguir Css de FES por meio da homogeneidade dimensional, ao passo que a similaridade de Morisita-Horn expressou as semelhanças entre as populações. A variabilidade observada entre sementes do Css e FES provavelmente reflete a diversidade genotípica das populações e as influências ambientais de cada fitofisionomia.
ABSTRACT
The Seed morphometry has taxonomic, phylogenetic and ecological importance, especially when it comes to native Cerrado (Brazilian savanna) trees. This study aims to discuss the potential relations between seeds dimensional characters, and the matrices environment of Dimorphandra mollis Benth from a cerrado sensu stricto (Css) and a seasonal semideciduous forest (FES). Were certain measures of length, diameter, and thickness of the seeds, and calculated the average diameters; geometric (DMG), equivalent (DME) arithmetic (DMA), surface area (As), and sphericity (Ø). Css seeds
showed smaller lengths (average 10.68 mm) and smaller diameter (average 5.35 mm), while FES seeds had a longer length (average 12.68 mm) and larger diameter (7.86 mm average), with a significant difference of 5% (p < 0.05) between sites. Pearson's correlation was more efficient to distinguish Css from FES by dimensional homogeneity, while Morisita-Horn similarity expressed similarities between populations. The variability observed between Css and FES seeds probably reflects the genotypic diversity of the populations and the environmental influences of each phytophysiognomy.
Keywords: Brazilian Savanna, phytophysiognomies, variability
1 INTRODUÇÃO
A identificação de características particulares em sementes e diásporos podem contribuir para aumento da eficiência na tecnologia de sementes e na produção de mudas de espécies nativas, voltadas tanto à restauração de áreas degradadas, quanto ao manejo e produção silvicultural (Batista et al., 2011, Ferreira & Barretto, 2015). Além disto, características estruturais de sementes apresentam valor não apenas em caráter taxonômico, como também subsídios para entendimento da plasticidade adaptativa e variabilidade genética dos indivíduos em diferentes fitofisionomias (Battilani et al., 2007). Neste sentido, estudos morfométricos são importantes ferramentas de investigação para acessar in locu as diferentes expressões fenotípicas intra e interpopulacionais (Liang et al., 2015).
O Cerrado é o segundo maior bioma brasileiro, perdendo em área, apenas para Amazônia, possui fauna e flora riquíssimas, com grande biodiversidade, compreendendo diferentes fitofisionomias que vão desde savanas a matas de galeria (Klink & Machado, 2015). Todavia, o desmatamento tem gerado preocupação, considerando os problemas ambientais provocados a este bioma, sobretudo, associados à perda da biodiversidade (Beuchle et al., 2015, Françoso et al., 2015, Panhol & Eyji, 2015). Dentre as muitas fitofisionomias ocorrentes no Cerrado destaca-se o cerrado sensu stricto (Css), caracterizado pelo acentuado escleromorfismo de seus representantes vegetais (Ribeiro et al., 2012, Gomes et al., 2026, Mendonça et al., 2017). Por outro lado, no cerrado (sensu lato) pode ser encontrada, especialmente na porção Sudeste do Mato Grosso do Sul (Brasil), a fitofisionomia de floresta estacional semidecidual (FES), fortemente relacionada ao bioma Mata Atlântica, e algumas vezes presente em regiões de ecótono (Dechoum et al., 2015, Ferreira et al., 2016, Argibay et al., 2020).
Uma importante espécie vegetal nativa ocorrente no Cerrado é Dimorphandra mollis Bent. (Leguminosae: Caesalpinioideae), popularmente conhecida como Fava-D’anta, por ser muito apreciada pela anta (Tapirus terrestris L.), sendo as sementes comumente encontradas em grande quantidade nas fezes destes animais (Feres et al., 2006). Está amplamente distribuída nas fitofisionomias do Cerrado brasileiro, com registros também na Bolívia e Paraguai (Costa et al., 2018), no Brasil sua ocorrência é descrita principalmente no Css (Costa et al., 2018, Souza & Lovato, 2010), e floresta decidual (FED) (Damasceno et al., 2018). Sendo raros os trabalhos que a descrevem para a FES (Marangon et al., 2003). As árvores adultas variam de 8 a 15 metros de altura, o fruto é um legume seco indeiscente, de coloração marrom escuro opaco, suas sementes são eurispérmicas de coloração variando do marrom ao vermelho-telha (Ferreira et al., 2001).
Informações sobre a estrutura dimensional e aspectos morfológicos de sementes, constituem parâmetro básico para compreender a ecologia das espécies, associadas à síndrome de dispersão, viabilidade e germinação das sementes (Battilani et al., 2007). Durante o processo de maturação as sementes desenvolvem-se até atingir o aspecto peculiar da espécie (Peñaloza & Durán, 2015), no entanto, uma mesma espécie pode apresentar variações relacionadas às flutuações ambientais as quais as matrizes estão sujeitas, além da variabilidade genética comum às espécies nativas (Santos et al., 2009, Oliveira et al., 2015).
Estudos morfométricos sobre os padrões estruturais de sementes de espécies nativas, em populações naturais distintas, subpopulações ou metapopulações do Cerrado são raros ou mesmo escassos. Para o gênero Dimorphandra à nível de caracterização taxonômica, a literatura disponibiliza artigos sobre morfologia de frutos, sementes, plântulas, plantas jovens e adultas (Ferreira et al., 2001). Para a diferenciação entre populações pode ser citado apenas o estudo de Souza & lovato (2010) que, avaliaram a diversidade genética em populações de D. mollis e D. wilsonii.
Portanto, partindo da hipótese de que os aspectos dimensionais das sementes podem expressar diferenças relacionadas ao ambiente de ocorrência das plantas, e que o acompanhamento dessas variações em trabalhos sistematizados possam ser úteis para se estimar diferenças restritas ao componente genético e/ou aos fatores ambientais, no presente estudo, foram discutidas as potencias relações entre os caracteres dimensionais das sementes e o ambiente de ocorrência em populações de plantas de D. mollis, oriundas de um Css, e uma FES.
2 MATERIAIS E MÉTODOS
O presente estudo foi conduzido no Laboratório de Ecologia, do Centro de Estudos em Recursos Naturais da Universidade Estadual de Mato Grosso do Sul (CERNA-UEMS), localizado no município de Dourados, Mato Grosso do Sul, Brasil. As coletas dos frutos foram realizadas no mês
de setembro, a partir de diferentes matrizes localizadas num fragmento de Css no município de Dourados/MS (S22º08’25” e W55º08’17”) e no município de Itaporã/MS (21°51’16.2”S e 54°55’45.0”W) num fragmento de FES.
Os frutos coletados estavam em perfeito estado, inteiros, sem injúrias e frescos, exalando quando abertos, o odor adocicado característico, semelhante a “biscoito”. Foram beneficiados manualmente sendo as sementes separadas e acondicionadas em refrigerador até o manuseio. Neste estudo foram avaliadas 100 sementes de D. mollis, de cada local de coleta, com o auxílio de paquímetro digital de precisão de 0,1 mm, foram tiradas medidas de comprimento (C) (medida da base até o ápice); diâmetro (D) (circunferência, ou seja, a largura da semente determinada a partir do seu ponto médio); e espessura (E) (distância determinada a partir do ponto médio (Figura 1).
Figura 1. Representação da estrutura dimensional das sementes de Dimorphandra mollis Benth. A- Vista superior do tegumento (tg), B- Corte longitudinal mediano no sentido do comprimento evidenciando o eixo embrionário (ex) e o cotilédone (ct), C- comprimento, D- diâmetro, E- espessura.
Em seguida, foram tomados, os diâmetros médios, geométrico (DMG), equivalente (DME) e aritmético (DMA), determinados de acordo com Sahay & Singh (1994. Para a medida de área superficial (As) em mm2, utilizou-se as equações descritas por McCabe et al. (2005). A esfericidade
das sementes (Ø) foi determinada em porcentagem (%) e o volume das sementes (V) em milímetro cúbico (mm3), calculados por meio dos valores observados nos eixos ortogonais, de acordo com
Mohsenin (1986). A relação de aspecto (Ra) foi calculada segundo Varnamkhasti et al. (2008).
Além destas variáveis, calculou-se a distribuição da frequência da razão As/V das sementes.
Também foi avaliado a hierarquia de aglomerados, por meio da análise de distância euclidiana. Para as observações e ilustrações das sementes, foi utilizado estereomicroscópio, provido de câmara clara. As ilustrações foram elaboradas utilizando nanquim e editadas no software ImageJ.
Os dados geométricos das sementes foram submetidos à análise de variância (ANOVA), e as médias ao teste de Tukey à 5% de probabilidade e correlação de Pearson. Em todo procedimento estatístico foi adotada a utilização do software R.
3 RESULTADOS
As sementes de D. mollis quando comparadas nas populações de Css e FES, apresentaram diferenças significativas (p < 0,05) entre os três eixos principais (C, D, E), bem como para as demais variáveis derivadas, com exceção de Ra (Tabela 1). Para as sementes Css foram observados menor C
e D com médias de 10,68 e 5,35 mm, respectivamente, ao passo que as sementes FES exibiram para os mesmos parâmetros médias maiores de 12,68 e 7,86 mm, respectivamente. Quanto aos caracteres relativos à forma D. mollis possui sementes variando de elíptica a reniforme, oblonga e algumas vezes levemente recurvada, com valores médios de Ø distintos para as duas populações estudadas, com média de 7,42 mm para as sementes Css e média de 6,75 mm para as sementes FES. Em geral, as sementes avaliadas apresentam três semieixos desiguais, associados aos baixos valores da Ra e Ø.
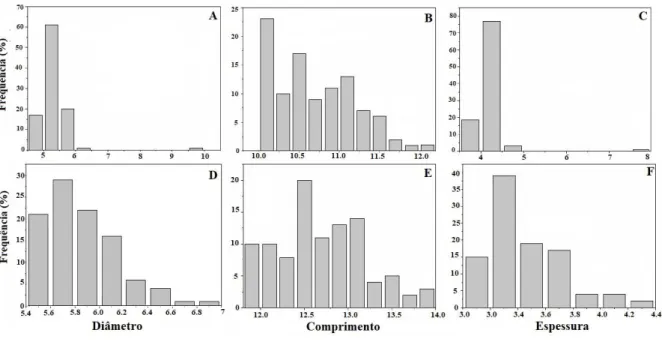
Cerca de 60% das sementes de D. mollis analisadas nesse estudo apresentaram diâmetro compreendido entre 5 a 5,5 mm para a população Css. Já para as sementes de FES apenas 28% apresentaram diâmetro entre 5,6 a 5,8 mm. Em relação ao comprimento, os maiores valores representam cerca de 22,5% e 20%, com comprimento de 10 a 10,2 mm e 12,5 mm para sementes de Css e FES, respectivamente. A espessura das sementes Css concentra cerca de 75% entre 4 a 4,5 mm, as de FES 38% concentram-se próximas de 3,3 mm.
Foram observadas diferenças significativas na As e no V (p<0,05). Os valores médios foram de
129,42 mm2 e 127,01 mm2 para As (Css e FES) e, 124,86 mm3 e 135,23 mm3 para V (Css e FES),
respectivamente.
Tabela 1. Valores médios dos parâmetros físicos de sementes de Dimorphandra mollis Benth., coletadas em duas diferentes fitofisionomias, cerrado stricto sensu (Css) e floresta estacional semidecidual (FES). Comprimento (C). Diâmetro (D). Espessura (E). Diâmetro médio geométrico (DMG). Diâmetro médio equivalente (DME). Diâmetro médio aritmético (DMA). Esfericidade (Ø). área superficial (As). Volume (V). Relação de aspecto (Ra).
Parâmetros Min x̄ Max E2 Min x̄ Max E2
Cerrado stricto sensu (Css) Floresta estacional semidecidual (FES)
C (mm) 10.02 10.68 b 12.01 0.05 11.83 12.68 a 13.94 0.05 D (mm) 4.67 5.35 b 9.95 0.05 5.46 7.86 a 6.94 0.03 E (mm) 3.87 4.16 a 7.87 0.04 3.02 3.44 b 4.32 0.02 DMG 5.78 6.18 b 8.25 0.03 5.82 6.34 a 7.41 0.03 DME 5.79 6.22 b 8.38 0.03 5.99 6.49 a 7.56 0.03 DMA 6.30 6.73 b 8.60 0.03 6.81 7.33 a 8.36 0.03 Ø 6.24 7.42 a 15.58 0.12 5.52 6.75 b 9.74 0.09 AS 104.94 120.42 b 213.80 1.43 106.52 127.01 a 172.62 1.44 V 101.09 124.86 b 293.97 2.47 103.38 135.23 a 213.26 2.35 Ra 0.41 0.50 a 0.87 0.005 0.46 0.47 a 0.49 0.0006
x̄ = média; E2 = erro padrão; Min = mínimo; Max = Máximo. Diferentes letras minúsculas na mesma linha representam
diferença significativa a 1% de probabilidade pelo teste de Tukey.
A distribuição da frequência para os três eixos principais das sementes, não apresentam distribuição aproximada da distribuição Gaussiana. Observa-se tanto para as sementes Css quanto para as FES que, a tendência de distribuição de frequências descreve a assimetria positiva (Figura 2).
Figura 2. Distribuição de frequência do diâmetro, comprimento, e espessura de sementes de Dimorphandra mollis Benth., de um cerrado stricto sensu (A, B, C) e de uma floresta estacional semidecidual (D, E, F).
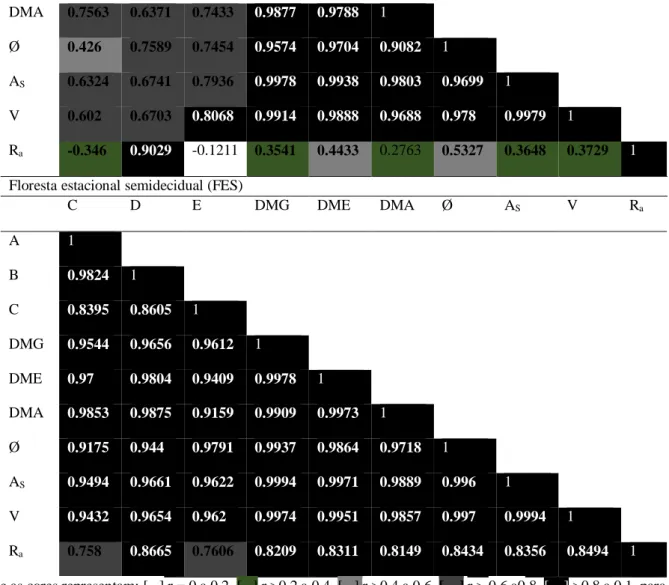
Os coeficientes de correlação entre as variáveis dimensionais para as sementes de Css e FES são apresentados na Tabela 2, sendo observado forte correlação positiva (>0,7) entre as variáveis descritas para FES.
Entre as variáveis C e E; D e E; E e Ra; Ra e DMA a diferença observada não foi significativa
para as sementes Css. Destacando-se ainda, que em relação à variável Ra essas sementes apresentaram
uma baixa correlação negativa com C e E (-0.3 e -0,1 respectivamente). Com base nos dados de correlação obtidos, foi possível verificar que algumas características dimensionais das sementes avaliadas nas duas populações de D. mollis apresentaram alta correlação, evidenciando a possibilidade de seleção direta e indireta destas características morfológicas (Tabela 2).
Tabela 2. Coeficientes de correlação não-paramétrica linear de Pearson entre as variáveis de sementes de Dimorphandra mollis Benth. de um Cerrado stricto sensu (Css) e uma floresta estacional semidecidual (FES). Comprimento (C). Diâmetro (D). Espessura (E). Diâmetro médio geométrico (DMG). Diâmetro médio equivalente (DME). Diâmetro médio aritmético (DMA). Esfericidade (Ø). área superficial (As). Volume (V). Relação de aspecto (Ra).
Cerrado stricto sensu (Css)
C D E DMG DME DMA Ø AS V Ra C 1 D 0.0885 1 E 0.5824 0.1328 1 DMG 0.6609 0.6743 0.7773 1 DME 0.6066 0.7455 0.7226 0.9945 1
DMA 0.7563 0.6371 0.7433 0.9877 0.9788 1
Ø 0.426 0.7589 0.7454 0.9574 0.9704 0.9082 1
AS 0.6324 0.6741 0.7936 0.9978 0.9938 0.9803 0.9699 1
V 0.602 0.6703 0.8068 0.9914 0.9888 0.9688 0.978 0.9979 1
Ra -0.346 0.9029 -0.1211 0.3541 0.4433 0.2763 0.5327 0.3648 0.3729 1
Floresta estacional semidecidual (FES)
C D E DMG DME DMA Ø AS V Ra A 1 B 0.9824 1 C 0.8395 0.8605 1 DMG 0.9544 0.9656 0.9612 1 DME 0.97 0.9804 0.9409 0.9978 1 DMA 0.9853 0.9875 0.9159 0.9909 0.9973 1 Ø 0.9175 0.944 0.9791 0.9937 0.9864 0.9718 1 AS 0.9494 0.9661 0.9622 0.9994 0.9971 0.9889 0.996 1 V 0.9432 0.9654 0.962 0.9974 0.9951 0.9857 0.997 0.9994 1 Ra 0.758 0.8665 0.7606 0.8209 0.8311 0.8149 0.8434 0.8356 0.8494 1
Onde as cores representam: [ ] r = 0 a 0.2, [ ] r >0.2 a 0.4, [ ] r >0.4 a 0.6, [ ] r > 0.6 a0.8, [ ] >0.8 a 0.1, para valores positivos ou negativos. Valores em negrito representam diferenças significativas a 1% (p<0,0001).
Observa-se na Figura 3, alta relação entre V e As, o modelo de regressão linear que melhor se
ajustou aos dados de V e As das sementes apresentou R2 > 0,99, para ambas as populações.
Figura 3. Regressão linear da área superficial e volume de sementes de Dimorphandra mollis Benth., de um cerrado stricto sensu (A), e de uma floresta estacional semidecidual (B).
Os agrupamentos por similaridade baseados nos caracteres morfológicos das sementes de D. mollis ocorrentes nas populações Css e FES são apresentados na Figura 4. O dendrograma utilizando a similaridade de Morisita-Horn apontou semelhanças entre as populações, com destaque para o isolamento da As e V, estes caracteres apresentaram maior distanciamento dos demais e maior
proximidade entre si. A ‘E e Ra’ das sementes de ambas as populações também se agrupam em um
conjunto. O comprimento se agrupa com as demais variáveis dimensionais. No entanto, o DMA e a Ø das sementes se alterna de uma população para a outra.
Figura 4. Dendograma utilizando a similaridade de Morisita-Horn para valores médios das análises dimensionais das sementes de Dimorphandra mollis Benth., de um cerrado sensu stricto (A) e de uma floresta estacional semidecidual (B). Em que: 1: Comprimento, 2: Diâmetro, 3: Espessura, 4: Diâmetro médio geométrico, 5: Diâmetro médio equivalente, 6: Diâmetro médio aritmético, 7: Esfericidade, 8: Área superficial, 9: Volume, 10: Relação de Aspecto.
4 DISCUSSÃO
As variáveis dimensionais das sementes de D. mollis, avaliadas no presente estudo sugerem diferenças fenotípicas ajustadas aos ambientes de ocorrência das populações de plantas. Com exceção de Ra a diferença significativa entre as variáveis representadas pelos três eixos ortogonais (Figura 1
e Tabela 1) e suas derivadas, sugerem variações de tamanho com seleção de sementes pequenas para a fitofisionomia de Css e sementes grandes para a FES.
As diferenças fenotípicas entre populações de plantas baseadas em caracteres morfológicos são normalmente assumidas como tendo origem genética (Rawat et al., 2017), por outro lado, também devem ser consideradas as flutuações (plasticidade) associadas à incerteza gerada pela heterogeneidade ambiental espacial e temporal (Valladares et al., 2007).
A variação no tamanho das sementes é comum em espécies nativas, tendo em vista, a variabilidade dentro e entre populações naturais (Gratani, 2014), por outro lado, dados relacionados à biometria de sementes podem contribuir na diferenciação de espécies pertencentes a um mesmo
gênero (Battilani et al., 2007, Roveri & Paula, 2017), ou mesmo possibilitar inferências acerca da variabilidade fenotípica em cultivares (Christo et al., 2014).
Os caracteres relativos à forma das sementes de D. mollis para Css e FES são equivalentes aos descritos por Ferreira et al. (2001) como variando de oblongas a reniformes. Em nosso estudo, referente à forma da semente foi observada também a variável Ø, sendo a média desta, maior para as sementes Css. Esta diferença observada entre Ø média de sementes de D. mollis das duas diferentes fitofisionomias deve ser avaliada com cuidado. Ainda que, a forma esférica (ou que mais se aproxime de esférica) possua valor adaptativo em propágulos, uma vez que sementes pequenas e arredondadas tendem a persistir e acumular com maior facilidade em banco de sementes, sendo em alguns casos, considerado caractere até mais importante que a própria massa (Thompson et al., 2001). Todavia, considerações comparativas entre populações (Css e FES) não devem ser realizadas com base nesta única variável, e sim acrescentá-la a um conjunto de caracteres, tornando a análise mais robusta.
Os aspectos morfológicos das sementes fornecem caracteres de importância ecológica, taxonômica e filogenética (Sadeghian et al., 2014). De modo geral, as características mais superficiais apresentam maior valor taxonômico e os caracteres internos são determinantes para diferenciação dos diferentes grupos vegetais (Rodrigues et al., 2015).
A distribuição assimétrica das frequências dimensionais das sementes de D. mollis, (Figura 2) independentemente do ambiente de origem, podem refletir a variabilidade genética dentro das populações. Sementes de espécies nativas normalmente geram dados de frequência não-Gaussiana, com distribuição multimodal, sendo está a distribuição esperada para amostras de uma mesma espécie ou quando comparadas diferentes espécies de um mesmo gênero (Rovner & Gyulai, 2007, Pontes et al., 2018).
As variáveis As e V apresentaram diferenças quando comparados para sementes de Css e FES.
Estas variáveis (derivadas do C, D e E) (Figura 3) são afetadas diretamente por fatores ambientais como tempo de secagem, embebição e requisitos energéticos das sementes durante o processo germinativo (Zareiforoush et al., 2011, Mir et al., 2013, Pontes et al., 2018), entre outros processos.
O V é definido pela ocupação de um objeto no espaço tridimensional, de maneira dependente às suas dimensões lineares, área superficial e massa (Mohsenin, 1986). Apesar de ainda incipientes em estudos, a razão As/V das sementes, do ponto de vista ecológico, pode estar associada à
longevidade das sementes no banco de sementes do solo em áreas de Cerrado, devido à resistência ao fogo ou aquecimento derivado de queimadas (Ruprecht et al., 2015), não se descartando os efeitos destas relações dimensionais na absorção e dissipação da energia térmica nestas condições.
Aspectos dimensionais das sementes podem também estar associados a aspectos reprodutivos. Em algumas espécies, sementes grandes normalmente são esperadas em frutos grandes (Jiménez et
al., 2016). A semente grande está diretamente ligada à maior reserva para o embrião e consequentemente para a plântula nos primeiros estádios do desenvolvimento, cuja dependência das reservas presentes nas folhas cotiledonares é maior (Han et al., 2017).
De acordo com os resultados apresentados na Tabela 2, existe uma forte correlação positiva (>0,7) entre as variáveis analisados para as sementes FES, sugerindo uma maior homogeneidade dimensional nos caracteres das sementes desta fitofisionomia, quando comparada às de Css. Tal fato pode ser relevante na distinção das populações, quando avaliados basicamente os lotes de sementes. A homogeneidade dimensional expressa nas sementes FES pode ser reflexo das condições próprias deste ambiente, cujas condições físicas e químicas tendem à maior homogeneidade (Ferreira et al., 2016). As sementes do Css podem ter sido expostas a interferências ambientais maiores, pois são sujeitas à fatores casuísticos como estiagem, fogo, variação da umidade relativa, temperatura, ventilação, entre outros (Gomes et al., 2016). Estes fatores são capazes de afetar o estado fisiológico das plantas matrizes (Mendes et al., 2016), com potenciais reflexos tanto nos caracteres vegetativos, naturalmente mais plásticos, como por exemplo, número de folhas, número de folíolos por folhas, tamanho da copa, entre outros, quanto nos reprodutivos, incluindo aqui os caracteres morfológicos das sementes.
A plasticidade fenotípica, quando implica na expressão de caracteres com valor adaptativo, pode contribuir para a sobrevivência de indivíduos e consequentemente das populações (Maldonado et al., 2017, Ruiz et al., 2019). A maior ou menor homogeneidade dimensional dos diásporos pode ser um indicativo de plasticidade adaptativa em espécies nativas sendo, portanto, mais um elemento que contribui para o entendimento da distribuição e ocorrência das espécies em ambientes distintos.
Em relação as análises de agrupamento (Figura 4). Pode-se dizer que, embora o DMA e a Ø das sementes tenham diferido entre as populações Css e FES, o dendrograma utilizando a similaridade de Morisita-Horn foi mais eficiente para apontar as semelhanças entre populações, com destaque para o isolamento entre a As e V das sementes como caracteres mais próximos entre si, possivelmente pela
relação direta entre estas variáveis. Todavia, devido à escassez de dados na literatura reportando tais variáveis em espécies do mesmo gênero ou família inviabilizou comparações sobre a análise de grupamentos com dados de outros autores.
Em síntese, ainda que em diversas espécies vegetais, a morfologia de sementes possa divergir conforme sua procedência e data de coleta (Nascimento et al., 2014). As variáveis dimensionais estudadas nas sementes de D. mollis apresentaram diferenças significativas entre os ambientes. O fato dos diásporos ocorrentes em FES serem normalmente maiores que os encontrados no Css, pode ser reflexo da diferença na informação ambiental. Fatores como a maior disponibilidade de nutrientes para a planta matriz, bem como fatores ecológicos (por exemplo competição por dispersores), podem
ser relevantes na expressão das diferenças fenotípicas nos dois ambientes. Também, deve ser considerado que as condições de radiação nos ambientes de FES e Css são distintas, o que implica em diferentes demandas de consumo/produção para plântulas no início do desenvolvimento (Ribeiro et al., 2016).
O desenvolvimento das sementes pode sofrer alterações pelas características genéticas, fisiológicas ou pelas flutuações nas condições ambientais (Nagel et al., 2015). A variação genética e do ambiente da planta-mãe, durante o tempo de maturação dos diásporos, são os principais fatores controladores dessa resposta (Baskin & Baskin, 1998).
Informações sobre características dimensionais de sementes, associadas com a sua
amplitude de variação dentro e entre populações, são importantes como critério de seleção de matrizes, podendo incrementar características de interesse produtivo (Gonçalves et al., 2013). Apesar da relevância e ampla distribuição de D. mollis, não existe na literatura dados em número suficiente que descrevam os aspectos dimensionais dos diásporos e as fitofisionomias de ocorrência das matrizes, de modo a permitir a identificação de padrões entre caracteres expressos e as fitofisionomias.
Ainda que a maior ou menor homogeneidade dimensional dos diásporos possa ser indicativo de plasticidade adaptativa em D. mollis sendo, portanto, mais um elemento que contribuiria no entendimento de sua ocorrência em ambientes distintos, é importante destacar que a ocorrência de D. mollis é predominantemente descrita para as fitofisionomias de Css, FED, e cerradão (CD), sendo pouco frequente na FES (Marangon et al., 2003), especialmente por sua baixa tolerância ao sombreamento (Pinheiro & Durigan, 2012). No presente estudo, as matrizes estão estabelecidas em ambiente de transição Cerrado e Mata atlântica o que provavelmente justifica a presença de D. mollis no fragmento de FES. Siqueira e Durigan (2007) ao proporem um modelo preditivo para a determinação da ocorrência de espécies lenhosas de Cerrado no estado de São Paulo, sugeriram a exclusão das áreas de transição como forma de aumentar a eficiência do modelo. Dessa forma, é possível supor que as diferenças nos caracteres dimensionais dos diásporos também sejam mais efetivas quando estudados material proveniente de áreas não ecotonais.
A distinção das populações de plantas com base em traços fenotípicos reconhecíveis em órgãos ou estruturas, mesmo que reprodutivas, não é tarefa elementar, dado a multiplicidade de fatores de variação ambiental. O acúmulo de informações descritivas dos caracteres morfológicos expressos em cada ambiente pode ser útil na identificação de padrões ajustados aos condicionantes presentes em diferentes fitofisionomias.
5 CONCLUSÕES
As sementes de D. mollis apresentam variabilidade para a maioria dos caracteres dimensionais analisados, bem como correlação significativa e positiva predominante entre determinados traços. As similaridades e diferenças observadas entre as duas populações estudadas refletem tanto a diversidade genotípica da espécie quanto as influências ambientais de cada fitofisionomia. Pesquisas sobre as diferenças fenotípicas entre populações de D. mollis são justificáveis tendo em vista a importância socioeconômica do táxon, e necessidade de uma compreensão detalhada de sua biodiversidade, bem como para a ampliação dos esforços na conservação do patrimônio gênico das populações de plantas nativas.
AGRADECIMENTOS
Os autores agradecem à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), código financeiro 001, pelas bolsas de pós-graduação concedidas a Jaqueline da Silva Santos e Montcharles da Silva Pontes.
CONFLITOS DE INTERESSE
Os autores declaram não ter conflito de interesse
REFERÊNCIAS
Smiderle, O. J.; Schwengber, D. R.; de Oliveira, J. M. F.; Jordão, S. M. S.; Gomes, H. H. 2020. Disponibilidade natural de sementes de espécies madeireiras em áreas de cerrado e floresta de Roraima. Brazilian Journal of Development, 6(3), 9625-9631. https://doi.org/10.34117/bjdv6n3-003
Ferreira, R. A.; Barretto, S. S. B. 2015. Caracterização morfológica de frutos, sementes, plântulas e Mudas de pau-brasil (Caesalpinia echinata Lamarck). Revista Árvore 39, 505-512. https://doi.org/10.1590/0100-67622015000300011.
Battilani, J. L.; Santiago, E. F.; Souza, A. L. T. D. 2007. Aspectos morfológicos de frutos, sementes e desenvolvimento de plântulas e plantas jovens de Unonopsis lindmanii Fries (Annonaceae). Acta
Liang, S.; Rong, X.; Sai, L., Chen, J.; Changqing, X.; Caixiang, X.; Tongning, L. 2015. Phenotypic variation of seed traits of Haloxylon ammodendron and its affecting factors. Biochemical
Systematics and Ecology 60, 81-87. https://doi.org/10.1016/j.bse.2015.04.012.
Klink, C. A.; Machado, R. B. 2005. A conservação do Cerrado brasileiro. Megadiversidade 1, 147-155.
Beuchle, R.; Grecchi, R. C.; Shimabukuro, Y. E.; Seliger, R.; Eva, H. D.; Sano, E.; Achard, F. 2015. Land cover changes in the Brazilian Cerrado and Caatinga biomes from 1990 to 2010 based on a systematic remote sensing sampling approach. Applied Geography 58, 116-127. https://doi.org/10.1016/j.apgeog.2015.01.017.
Françoso, R. D.; Brandão, R.; Nogueira, C. C.; Salmona, Y. B.; Machado, R. B.; Colli, G. R. 2015. Habitat loss and the effectiveness of protected areas in the Cerrado Biodiversity. Hotspot. Natureza
& Conservação 13, 35-40. https://doi.org/10.1016/j.ncon.2015.04.001.
Panhol, B. A.; Eyji, S. E. 2015. Séries temporais de índices de vegetação (NDVI e EVI) do sensor modis para detecção de desmatamentos no bioma cerrado. Bulletin of Geodetic Sciences 21, 797-813. https://doi.org/10.1590/S1982-21702015000400047.
Ribeiro, M. N.; Pedroni, F.; Peixoto, K. D. S. 2012. Fogo e dinâmica da comunidade lenhosa em cerrado sentido restrito, Barra do Garças, Mato Grosso. Acta Botanica Brasilica 26, 203-217. https://doi.org/10.1590/S0102-33062012000100020.
Gomes, L.; Maracahipes, L.; Reis, S. M.; Marimon, B. S.; Marimon-Junior, B. H.; Lenza, E. 2016. Dynamics of the woody vegetation of two areas of Cerrado sensu stricto located on different substrates. Rodriguésia 67, 859-870. https://doi.org/10.1590/2175-7860201667401.
Torres, D. M.; Fontes, M. A. L.; Samsonas, H. D. P. 2017. Relações solo-vegetação na estruturação de comunidades de cerrado sensu stricto no sul de Minas Gerais, Brasil. Rodriguésia 68, 115-128. https://doi.org/10.1590/2175-7860201768121.
Dechoum, M. S.; Castellani, T. T.; Zalba, S. M.; Rejmánek, M.; Peroni, N.; Tamashiro, J. Y. 2015. Community structure, succession and invasibility in a seasonal deciduous forest in southern Brazil.
Biological Invasions 17, 1697-1712. https://doi.org/10.1007/s10530-014-0827-6.
Ferreira, M. P.; Zortea, M.; Zanotta, D. C.; Shimabukuro, Y. E.; Souza Filho, C. R. 2016. Mapping tree species in tropical seasonal semi-deciduous forests with hyperspectral and multispectral data.
Remote Sensing of Environment 179, 66-78. https://doi.org/10.1016/j.rse.2016.03.021.
Argibay, D. S.; Sparacino, J.; Espindola, G. M. 2020. A long-term assessment of fire regimes in a Brazilian ecotone between seasonally dry tropical forests and savannah. Ecological Indicators 113, 106151. https://doi.org/10.1016/j.ecolind.2020.106151.
Féres, C. A. O.; Madalosso, R. C.; Rocha, O. A.; Leite, J. P. V.; Guimarães, T. M. D. P.; Toledo, V. P. P.; Tagliati, C. A. 2006. Acute and chronic toxicological studies of Dimorphandra mollis in experimental animals. Journal of Ethnopharmacology 108, 450-456. https://doi.org/10.1016/j.jep.2006.06.002.
Costa, G. R.; Carvalho, S. T. T.; Silva, T. G. N.; Andrade, V. C. L.; Souza, P. B. 2018. Fitossociologia e estrutura diamétrica de um fragmento de Cerrado sensu stricto, Formoso do Araguaia, Tocantins.
Revista Verde De Agroecologia E Desenvolvimento Sustentável 13, 501-507. http://dx.doi.org/10.18378/rvads.v13i4.5552.
Souza, H. A. V; Lovato, M. B. 2010. Genetic diversity and structure of the critically endangered tree Dimorphandra wilsonii and of the widespread in the Brazilian Cerrado Dimorphandra mollis: Implications for conservation. Biochemical Systematics and Ecology 38, 49-56. https://doi.org/10.1016/j.bse.2009.12.038.
Damasceno-Junior, G. A.; Pott, A.; Neves, D. R. M.; Sciamarelli, A.; Fina, B. G. 2018. Flora lenhosa de Florestas estacionais do estado de Mato Grosso do Sul: estado da arte. Iheringia. Série Botânica 73, 65-79. https://doi.org/10.21826/2446-8231201873s65.
Marangon, L. C.; Soares, J. J.; Feliciano, A. L. P. 2003. Florística arbórea da Mata da Pedreira no município de Viçosa, MG. Revista Árvore 27, 207-215. https://doi.org/10.1590/S0100-67622003000200010.
Ferreira, R. A.; Botelho, S. A.; Davide, A. C.; Malavasi, M. D. M. 2001. Morfologia de frutos, sementes, plântulas e plantas jovens de Dimorphandra mollis Benth. –faveira (Leguminosae- Caesalpinioideae). Brazilian Journal of Botany 24, 303-309. https://doi.org/10.1590/S0100-84042001000300009.
Peñaloza, P.; Durán, J. M. 2015. Association between biometric characteristics of tomato seeds and seedling growth and development. Electronic Journal of Biotechnology 18, 267-272. https://doi.org/10.1016/j.ejbt.2015.04.003.
Santos, F. S.; Paula, R. C.; Sabonaro, D. Z.; Valadares, J. 2009. Biometria e qualidade fisiológica de sementes de diferentes matrizes de Tabebuia chrysotricha (Mart. Ex A. DC.) Stand I. Scientia
Forestalis 37, 163-173. http://hdl.handle.net/11449/71027.
Oliveira Melo, A. T.; Coelho, A. S. G.; Pereira, M. F.; Blanco, A. J. V.; Franceschinelli, E. V. 2015. Genética da conservação de Cabralea canjerana (Vell.) Mart. (Meliaceae) em fragmentos florestais de Mata Atlântica na Apa Fernão Dias. Revista Árvore 39, 365-374. https://doi.org/10.1590/0100-67622015000200016.
Sahay, K. M; Singh, K. K. 1994. Unit operations of agricultural processing. New Delhi: Vikas Publishing House Pvt Ltd.
McCabe, W. L.; Smith, J. C.; Harriott, P. 2005. Unit operation of chemical engineering. New York: McGraw-hill.
Mohsenin, N. N. Physical properties of plant and animal materials. 2th Revised. New York, 1986.
Varnamkhasti, M. G.; Mobli, H.; Jafari, A.; Keyhani, A. R.; Soltanabadi, M. H.; Rafiee, S.; Kheiralipour, K. 2008. Some physical properties of rough rice (Oryza sativa L.) grain. Journal of
Cereal Science 47, 496-501. https://doi.org/10.1016/j.jcs.2007.05.014.
Rawat, S.; Jugran, A. K.; Bhatt, I. D.; Rawal, R. S.; Nandi, S. K. 2017. Effects of genetic diversity and population structure on phenolic compounds accumulation in Hedychium spicatum. Ecological
Valladares, F.; Gianoli, E.; Gómez, J. M. 2007. Ecological limits to plant phenotypic plasticity. New
Phytologist 176, 749-763. https://doi.org/10.1111/j.1469-8137.2007.02275.x.
Gratani, L. 2014. Plant phenotypic plasticity in response to environmental factors. Advances in
botany 2014, 1-17. https://doi.org/10.1155/2014/208747.
Roveri N. A.; Paula, R. C. 2017.Variability among mother trees of Ceiba speciosa St. Hil for characteristics of the fruits and seeds. Revista Ciência Agronômica 48, 318-327. https://doi.org/10.5935/1806-6690.20170037.
Christo, L. F.; Colodetti, T. V.; Rodrigues, W. N.; Martins, L. D.; Amaral, J. F. T.; Laviola, B. G.; Tomaz, M. A. 2014. Genetic variability among genotypes of physic nut regarding seed biometry.
American Journal of Plant Sciences 5, 1416-1422. https://doi.org/10.4236/ajps.2014.510156.
Thompson, K.; Jalili, A.; Hodgson, J. G.; Hamzeh'ee, B.; Asri, Y.; Shaw, S.; Ghahramani, M. A. 2001. Seed size, shape and persistence in the soil in an Iranian flora. Seed Science Research 11, 345-355. https://doi.org/10.1079/SSR200191.
Sadeghian, S.; Zarre, S.; Heubl, G. 2014. Systematic implication of seed micromorphology in Arenaria (Caryophyllaceae) and allied genera. Flora 209, 513-529. https://doi.org/10.1016/j.flora.2014.07.004.
Rodrigues, J. K.; Mendonça, M. S. D.; Gentil, D. F. D. O. 2015. Morphoanatomical, histochemical and biometric aspects of pyrene of Bactris maraja (Arecaceae). Rodriguésia 66, 75-85. https://doi.org/10.1590/2175-7860201566105.
Rovner I.; Gyulai F. 2007. Computer-assisted morphometry: a new method for assessing and distinguishing morphological variation in wild and domestic seed populations. Economic Botany 61, 154-172. https://doi.org/10.1663/0013-0001(2007)61[154:CMANMF]2.0.CO;2.
Pontes, M. S.; Montefusco-Pereira, C. V.; Misra, B. B.; Ribeiro-Junior, H. L.; Graciano, D. E.; Santos, J. S.; Santiago, E. F. 2018. High-throughput phenotyping by applying digital morphometrics and fluorescence induction curves in seeds to identifying variations: A case study of Annona (Annonaceae) species. Information Processing in Agriculture 5, 443-455.
Zareiforoush, H.; Hosseinzadeh, B.; Adabi, M. E.; Motavali, A. 2011. Moisture dependent physical properties of paddy grains. The Journal of American Science 7, 175-182, 2011. https://doi.org/10.1016/S0021-8634(86)80060-9.
Mir, S. A.; Bosco, S. J. D.; Sunooj, K. V. 2013. Evaluation of physical properties of Rice cultivars grown in the temperate regions of India. International Food Research Journal 20, 1521-1527. https://doi.org/10.1002/fsn3.746.
Ruprecht, E.; Fenesi, A.; Fodor, E. I.; Kuhn, T.; Tökölyi, J. 2015. Shape determines fire tolerance of seeds in temperate grasslands that are not prone to fire. Perspectives in Plant Ecology 17, 397-404. https://doi.org/10.1016/j.ppees.2015.07.001.
Jiménez‐Alfaro, B.; Silveira, F. A.; Fidelis, A.; Poschlod, P.; Commander, L. E. 2016. Seed germination traits can contribute better to plant community ecology. Journal of Vegetation Science 27, 637-645. https://doi.org/10.1111/jvs.12375.
Han, C.; Zhen, S.; Zhu, G.; Bian, Y.; Yan, Y. 2017. Comparative metabolome analysis of wheat embryo and endosperm reveals the dynamic changes of metabolites during seed germination. Plant
Physiology and Biochemistry 115, 320-327. https://10.1016/j.plaphy.2017.04.013.
Mendes, K. R.; Machado, S. R.; Amaro, A. C. E.; Silva, S. C. M.; Júnior, V. F.; Rodrigues, T. M. 2016. Distribution of homobaric and heterobaric leafed species in the Brazilian Cerrado and seasonal semideciduous forests. Flora 225, 52-59. https://doi.org/10.1016/j.flora.2016.10.005.
Maldonado-Chaparro, A. A.; Read, D. W.; Blumstein, D. T. 2017. Can individual variation in phenotypic plasticity enhance population viability?. Ecological Modelling 352, 19-30. https://doi.org/10.1016/j.ecolmodel.2017.02.023.
Ruiz, M. B.; D’Andrea, K. E.; Otegui, M. E. 2019. Phenotypic plasticity of maize grain yield and related secondary traits: Differences between inbreds and hybrids in response to contrasting water and nitrogen regimes. Field Crops Research 239, 19-29. https://doi.org/10.1016/j.fcr.2019.04.004.
Nascimento, M. E.; Bertolucci, S. K. V.; Santos, F. M.; Santos Jr, J. M.; Castro, E. M.; Pinto, J. E. B. P. 2014. Morphlogical evaluation of Copaifera langsdorffii Desf. saplings grown in different temperatures. Revista Brasileira de Plantas Medicinais 16, 931-937.
Ribeiro, J. W. F.; Kolb, R. M. 2016. Distinct germination responses may contribute to the distribution pattern of two Moquiniastrum species in different phytophysiognomies from the Brazilian savanna.
Flora 223, 159-166. https://doi.org/10.1016/j.flora.2016.06.001.
Nagel, M.; Kranner, I.; Neumann, K.; Rolletschek, H.; Seal, C.E.; Colville, L.; Börner, A. 2015. Genome‐wide association mapping and biochemical markers reveal that seed ageing and longevity are intricately affected by genetic background and developmental and environmental conditions in barley. Plant, Cell & Environment 38, 1011-1022. https://doi.org/10.1111/pce.12474.
Baskin, C. C.; Baskin, J. M. Seeds: ecology, biogeography, and evolution of dormancy and
germination. Academic Press, London, 1998.
Gonçalves, L. G. V.; Andrade, F. R.; Marimon, J. B. H.; Schossler, T. R.; Lenza, E. E.; Marimon, B. S. 2013. Biometria de frutos e sementes de mangaba (Hancornia speciosa Gomes) em vegetação natural na região leste de Mato Grosso, Brasil. Revista Brasileira de Ciências Agrárias 36, 31-40.
Pinheiro, E. D. S.; Durigan G. 2012. Diferenças florísticas e estruturais entre fitofisionomias do Cerrado em Assis, SP, Brasil. Revista Árvore 36, 181-193, 2012. https://doi.org/10.1590/S0100-67622012000100019.