Screening for antioxidant activity in extracts obtained from extremophile microalgae
Texto
Imagem

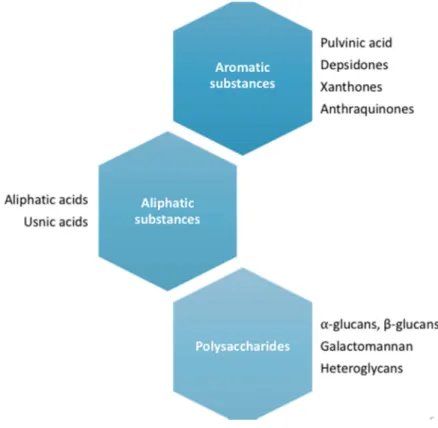
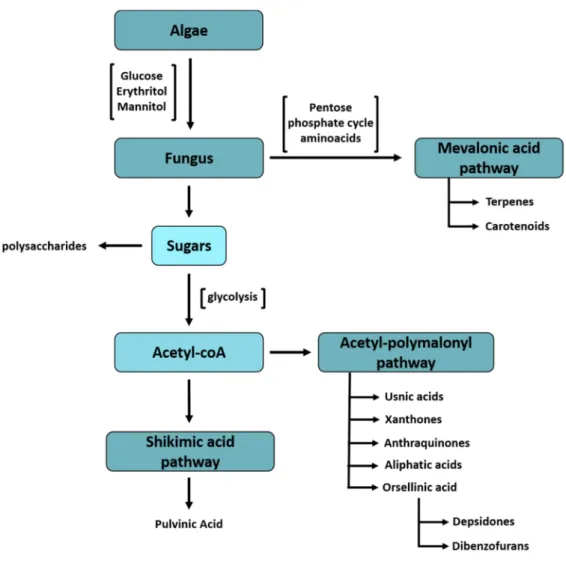


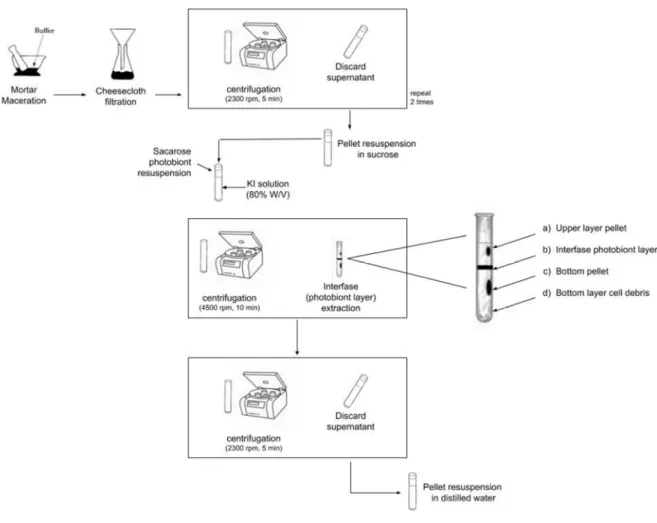
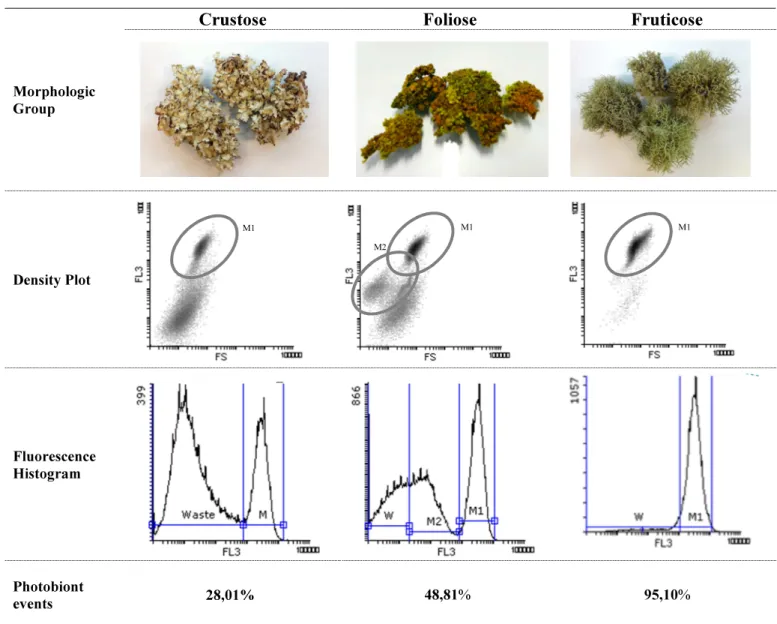



Documentos relacionados
O processo de orientac;ao adoptado nestas disciplinas, bem como o levantamento das lacunas registadas na elaborac;ao dos projectos de intervenc;ao dos formandos, permitiram detectar
Os resultados preliminares indicam que o processo formativo do professor de Língua Gestual Portuguesa reflete uma conjuntura de necessidades de valorização desta língua, do
Foram elaborados três bolos com concentrações (A=16%, B=14% e C=12%) diferentes de pó alimentício da casca de manga, para verificação da melhor concentração que agrada ao
Thus, the sovereign inequality affirmed in the Al Bashir Case, based on the rules established by the Rome Statute, is also present in the fundamental institutions, since
no canto superior esquerdo, escrito a tinta preta, no sentido horizontal, frente e verso], não datado e sem carimbo do correio, sem destinatário explícito.. Meu
DevOps Practices in Incident Management Process xv Abbreviations AM – Application Management CI – Continuous Integration CS – Case Study IM – Incident Management IT
MNEs' international experience offsets barriers imposed by institutional distance between home and host countries on MNEs' entry strategy such that for both low and high levels
EC 50 values (mg/mL) obtained for the antioxidant activity by scavenging of 2,2-diphenyl-1-picryl-hydrazyl radicals (DPPH) and β -carotene bleaching assay (BCB) in the
