Universidade Federal de Uberlândia
Instituto de Biologia
Pós-Graduação em Ecologia e Conservação dos Recursos Naturais
Remoção de sementes por formigas, aves e roedores em área
de Cerrado e seu efeito sobre o recrutamento de plântulas
Alana Vaz Ferreira
Alana Vaz Ferreira
Remoção de sementes por formigas, aves e roedores em área
de Cerrado e seu efeito sobre o recrutamento de plântulas
Dissertação apresentada à Universidade Federal de Uberlândia, como parte das exigências para obtenção do título de Mestre em Ecologia e Conservação de Recursos Naturais.
Orientador
Prof. Dr. Heraldo Luís de Vasconcelos
Alana Vaz Ferreira
Remoção de sementes por formigas, aves e roedores em área
de Cerrado e seu efeito sobre o recrutamento de plântulas
Dissertação apresentada à Universidade Federal de Uberlândia, como parte das exigências para obtenção do título de Mestre em Ecologia e Conservação de Recursos Naturais.
APROVADA em_____ de _______________de 2008
_______________________________________ Prof. Dr. Emilio Miguel Bruna
University of Florida
_________________________________
Prof. Dr. Inara..Roberta. LealUniversidade Federal de Pernambuco
UBERLÂNDIA Fevereiro de 2008
__________________________________________ Prof. Dr. Heraldo Luís de Vasconcelos
AGRADECIMENTOS
Aos meus pais Alan Kardec e Doraci pelo apoio, paciência e incentivo.
Aos meus irmãos Heitor e Aline pela paciência em muitos momentos de mau humor e estresse.
Ao Heraldo pela oportunidade e confiança, pelos ensinamentos e por me mostrar que críticas não devem ser entendidas como ofensas ou broncas e sim como oportunidades ricas de aprendizado e crescimento.
Ao Emilio pela oportunidade de trabalho, pelo apoio a esse projeto e por me ensinar, mesmo sem perceber, que devemos nos empolgar sempre ao menor resultado alcançado.
Aos companheiros Alan, Cauê e Renata pela preciosa ajuda em campo, pelos palpites, idéias, conselhos.
Aos demais companheiros do LEIS pelos momentos divertidos que passamos em campo, no laboratório ou nas pequenas “reuniões” em finais de semana.
Às minhas amigas Carol, Cecília, Furinho, Patrícia e Rafaela que mesmo um pouco distantes sempre estiveram perto torcendo por mim.
SUMÁRIO
Resumo...iii
Abstract...iv
1. Introdução...01
2. Material e Métodos...04
2.1. Local de estudo...04
2.2. Determinação das taxas de remoção...04
2.2. Efeitos da remoção de sementes no recrutamento de plântulas...08
2.3. Identificação das formigas removedoras de sementes...11
3. Resultados...13
3.1. Determinação das taxas de remoção de sementes...13
3.2. Efeitos da remoção de sementes no recrutamento de plântulas...18
3.3. Identificação das formigas removedoras de sementes...22
4. Discussão...24
4.1. Remoção de sementes do Cerrado por formigas, aves e roedores...24
4.2. Variações espaciais na remoção de sementes...26
4.3. Variações interespecíficas na remoção de sementes...27
4.4. Destino das sementes removidas por formigas: Predação ou dispersão secundária?.28 4.5. Impacto da remoção de sementes no recrutamento de plântulas...31
5 Conclusões...32
5. Referências Bibliográficas...34
Resumo
Ferreira, Alana V. 2008. Remoção de sementes por formigas, aves e roedores em área de Cerrado e seu efeito sobre o recrutamento de plântulas. Dissertação de Mestrado em Ecologia e Conservação de Recursos Naturais. Universidade Federal de Uberlândia. Uberlândia_MG. 43p.
Muitas populações de plantas são afetadas pela atuação dos consumidores de sementes como formigas, aves e roedores. Essa remoção pode influenciar a estruturação dessas populações através da alteração da taxa e do padrão de recrutamento. Os objetivos do estudo foram determinar qual a importância relativa de formigas, aves e roedores nas taxas de remoção de sementes em diferentes fitofisionomias de Cerrado e quais os efeitos dessa remoção no recrutamento de plântulas. O estudo foi realizado na Estação Ecológica do Panga, em Uberlândia - MG, de setembro de 2006 a janeiro de 2008 nas fitofisionomias cerrado sentido restrito, cerrado denso e cerradão. Determinei as taxas de remoção de sementes das espécies Coussarea hydrangeaefolia, Guapira graciliflora, Maprounea guianensis, Matayba guianensis, Siparuna guianensiseVirola sebiferautlizando três tratamentos, um com acesso às sementes somente por
formigas, o segundo com acesso por formigas e roedores e um terceiro em que formigas aves e roedores tiveram acesso. Em cada tratamento foram expostas 20 sementes de cada espécie por 96 horas. As taxas de remoção por formigas foi cerca de 10% maior que pelos outros agentes. As sementes de Maprounea guienensis foram as mais removidas com 92,2% da porcentagem total
de remoção. Para determinar os efeitos da remoção de sementes no recrutamento de plântulas utilizei parcelas estabelecendo dois tratamentos, parcelas de livre acesso para formigas, aves e roedores e parcelas onde esses agentes foram excluídos. Nessas parcelas foram adicionadas sementes de Guapira graciliflora. O recrutamento nas parcelas de exclusão de agentes foi cerca
de 4 vezes maior que nas parcelas abertas aos agentes.
Abstract
Ferreira, Alana V. 2008. Seed removal by ants, birds and rodents in a cerrado area and its effects on seedling recruitment
Many plant populations are affected by seed consumers since these can affect the rates and
patterns of seedling recruitment. Ants, birds and rodents are main agents of seed removal.
Therefore, the aim of this study was to determine the relative importance of ants, birds and
rodents on seed removal in different types of Cerrado vegetations and to determine the effects of
seed removal on seedling recruitment. Field work was performed at Panga Ecological Station
near Uberlândia, MG, Brazil, in the foolowing vegetation types: cerrado sensu stricto, cerrado
denso e cerradão. I determined the seed removal rates for the following tree species: Coussarea
hydrangeaefolia, Guapira graciliflora, Maprounea guianensis, Matayba guianensis, Siparuna
guianensis e Virola sebifera. Seeds were placed in three types of stations, one with access for
ants only, one with access to ants and rodents and with access to ants, rodents and birds. In each
station I placed 20 seeds of each species that were left in place for 96h. Seed removal rates
varied little among the three types of vegetation. Maprounea guianensiwas the species with the
greatest removal rate: 97,4%. Of the seeds removed for ants were 10 more than birds and
rodents. To determine the effects of seed removal on seedling recruitment I built a series of
exclosures (each 70 x 70 cm), and compared seedling recruitment (both for planted seeds of
Guapira graciliflora and for existing seeds of all dicot species) between predator exclosures and
control plots. There were significantly more seedlings of Guapira graciliflora in the exclosure
than in control plots. Also, 8 months after removal of seed consumers, I detected a significantly
greater rate of natural seedling emergence in the exclosure plots.
Key-words: savannas. Brazil, seed predation, granivory, elaiossome, ecology of plant-animal
1. INTRODUÇÃO
Muitas populações de plantas têm seu recrutamento reduzido pela atuação de
consumidores de sementes (Andersen & Ashton, 1985; Simms & Maron, 2000; Vander Wall et.
al. 2005). O consumo de sementes pode influenciar diretamente a estrutura de populações
(Crawley, 1992). O efeito da remoção de sementes é considerado importante em populações de
plantas que não são limitadas por outros fatores, como a existência de microhabitats adequados
para o estabelecimento e como a competição por espaço para esse estabelecimento, embora seja
difícil comparar a magnitude de cada um desses fatores com os efeitos da predação de sementes
(Edwards & Crawley, 1999). No Cerrado, estudos mostram que o acúmulo de matéria orgânica
no solo e a cobertura vegetal podem ser importantes facilitadores ou inibidores no recrutamento
de plântulas (Hoffmann, 1996). Mas, a predação e a remoção de sementes é também um fator a
ser considerado quando falamos em impactos sobre o recrutamento de plântulas (Andersen,
1989; Maron & Simms, 1997; Vander Wallet. al., 2005), embora para o Cerrado pouco ainda se
saiba sobre este fator.
A remoção de sementes, seja por predação ou por dispersão secundária, é realizada por
vários agentes abióticos e bióticos. Entre os agentes bióticos destacam-se as formigas, as aves
granívoras e os roedores que utilizam as sementes ou apenas partes delas como recurso alimentar
(Crawley, 1992; Hulme, 1998; Pizo & Vieira, 2004, Pèrez et. al, 2006). Fatores como a
abundância de sementes, as características do local de deposição e as características físicas das
sementes, tais como peso, presença de polpa ou elaiossoma (i.e. apêndice de composição lipídica
presente em sementes de algumas espécies), são características importantes quando se investiga
as atividades de consumo e remoção de sementes (Reader, 1993; Edwards & Crawley, 1999).
Muitas formigas são predadoras de sementes, porém ao transportar estas sementes por
pequenas distâncias podem também modificar a deposição realizada pelos dispersores primários,
(Robert & Heithaus, 1986). Muitos trabalhos falam sobre o efeito positivo de formigas para a
germinação de sementes e, consequentemente, para o recrutamento de plântulas, na medida em
que estas formigas consomem a polpa ou o arilo das sementes, facilitando a germinação (Leal &
Oliveira, 1998; Passos & Oliveira, 2004; Christianini, et. al., 2007). Outros destacam a
importância destas ao transportar as sementes para locais tidos como mais propícios para a
germinação como os ninhos (Passos & Oliveira, 2004).
A remoção de sementes pode ter efeitos positivos e negativos sobre a dinâmica de
populações vegetais. Os roedores são considerados predadores de sementes e plântulas, causando
redução no recrutamento de plântulas em vários ecossistemas, como em campos de herbáceas
(Ostfeld et. al., 1997), campos de gramíneas (Maron & Simms, 2001) e em vegetação xérica
(Gutièrrez, et. al., 1997). A magnitude desses efeitos depende de fatores como a estação do ano,
a densidade de roedores e o grau de perturbação do hábitat (Ostfeld et. al., 1997; Casenave et.
al., 1998; Pizo & Vieira, 2004). As aves atuam mais frequentemente dispersando sementes que
passam intactas pelo trato digestivo (Sick, 1997), mas também podem causar efeitos negativos
como a granivoria (Casenave et. al., 1998; Pizo & Vieira, 2004). A importância de aves como
predadoras dependerá também da sazonalidade e do grau de perturbação, sendo que a remoção
por aves parece ser importante em áreas onde os roedores apresentam baixas densidades
(Casenave et. al., 1998; Pizo & Vieira, 2004). As formigas aparecem como importantes
dispersoras primárias e secundárias de sementes (Horvitz & Shemske, 1986; Andersen &
Ashton, 1985; Kelt, et. al., 2004; Christianini et. al., 2007). A importância de formigas como
agentes removedoras é documentada em vários ecossistemas como Cerrado (Leal & Oliveira,
1998; Christianini et. al., 2007), Mata Atlântica (Pizo & Oliveira, 1998, 2000, 2001), Floresta
Semi-decídua (Passos & Ferreira, 1996), Caatinga (Leal, 2003), Restinga (Passos & Oliveira,
2003), Floresta de Eucaliptos na Austrália (Andersen & Ashton, 1985) e áreas semi-áridas no
Chile (Kelt et. al., 2004). O efeito da interação entre formigas e sementes depende das espécies
atrativos. Mas, de um modo geral, não há um padrão para estas atividades entre os diferentes
ecossistemas, os efeitos da remoção dependerão da densidade de agentes, da disponibilidade de
recursos em cada ecossistema e da sazonalidade em alguns casos.
No Cerrado muito estudos indicam a importância de fatores abióticos determinando
características das populações de plantas e conseqüentemente, das formações típicas desse
Bioma. Entre esses fatores temos a composição dos solos, a sazonalidade climática, que expõe as
populações a forte estresse hídrico durante parte do ano, e o fogo como um dos mais importantes
e estudados. Já os efeitos da atuação de agentes bióticos sobre as plantas dos ecossistemas de
Cerrado tem sido ainda pouco estudados, com destaque para alguns trabalhos que tratam dos
efeitos positivos da atuação de formigas sobre a germinação em espécies de Cerrado (Leal &
Oliveira, 1998; Christianini et. al., 2007). Alguns trabalhos feitos em outros ecossistemas
destacam efeitos negativos de vertebrados, como os roedores, no recrutamento e sobrevivência
de plântulas (Gutièrrez, et. al, 1997; Maron & Simms, 1997; Ostfeld et. al., 1997; Casenave &
Marone, 1998; Edwards & Crawley, 1999; Maron & Simms, 2001). Ainda temos poucas
informações sobre a influência que herbívoros, predadores e dispersores de sementes podem
exercer na dinâmica das populações de plantas do Cerrado. Assim, o presente estudo teve como
objetivo: 1) determinar as taxas de remoção de sementes por aves, formigas e roedores em áreas
de cerrado 2) determinar se existe variação nas taxas de remoção entre espécies de plantas e/ou
entre diferentes fitofisionomias, 3) determinar o (s) destino (s) das sementes removidas e 4)
2. MATERIAL E MÉTODOS
2.1. Local de estudo
O estudo foi conduzido na Estação Ecológica do Panga, uma Reserva Particular do
Patrimônio Natural (RPPN), administrada pela Universidade Federal de Uberlândia. A reserva
dista 30 km do centro da cidade de Uberlândia; MG, entre as coordenadas 19º 09’ 20”- 19º 11’
10” Sul e 48º 23’ 20”- 48º 24’ 35” Oeste, abrangendo uma área total de 403,5 ha. Localiza-se a
uma altitude média de 800 metros e o clima da região é classificado como tipo AW, segundo a
classificação de Köppen, onde ocorrem duas estações bem definidas: verões quentes e úmidos,
de outubro a março e invernos frios e secos, de abril a setembro (Schiavini e Araújo, 1989). A
temperatura média é de 22ºC e a precipitação anual é de 1550 mm (Rosaet al., 1991).
O estudo foi realizado nas seguintes fitofisionomias do Cerrado: cerrado sentido restrito,
cerrado (sentido restrito) denso e cerradão. O cerrado (sentido restrito) é caracterizado por
árvores de médio porte, 3 a 8 m, com estrato herbáceo-arbustivo denso, com destaque para as
formas arbóreas tortuosas, com folhas coriáceas e ramos e caules revestidos por espessa camada
de súber (Schiavini e Araújo, 1989). O cerrado (sentido restrito) denso apresenta a mesma
composição de espécies que o cerrado sentido restrito, mas com maior densidade de arbórea
(Vasconceloset. al., dados não publicados). O cerradão é considerado um dos tipos florestais da
vegetação de Cerrado devido ao sombreamento proporcionado pelo dossel formado por árvores
de maior porte, com 10 a 15 m (Schiavini e Araújo, 1989).
2.2. Determinação das taxas de remoção de sementes
Os experimentos para determinação das taxas de remoção foram realizados nas seguintes
fitofisionomias: cerrado sentido restrito, cerrado (sentido restrito) denso e cerradão. Neste
experimento determinei as taxas de remoção de sementes por formigas, aves e roedores,
diferentes tipos de sementes visou determinar as taxas de remoção levando em consideração as
diferenças morfológicas das sementes como peso, presença de polpa ou de elaiossoma (i.e.
apêndice composto por substâncias lipídicas).
Foram utilizadas para os testes sementes de seis espécies arbóreas: Coussarea
hydrangeaefolia (Benth.) Müll.Arg (Rubiaceae), Guapira graciliflora (Mart.) Lundell
(Nyctaginaceae), Maprounea guianensis Aublet, (Euphorbiaceae), Matayba guianensis Aublet
(Sapindaceae), Siparuna guianensis Aublet (Monimiaceae), Virola sebifera Aublet
(Miristicaceae). A espécie C. hydrangeaefolia vulgarmente chamada de falsa-quina têm alturas
entre 4 e 5 metros, o fruto é do tipo drupa globosa, com polpa suculenta, de cor branca quando
madura, contendo uma única semente, produz anualmente moderada quantidade de sementes
viáveis, disseminadas pela avifauna, floresce em mais de uma época do ano, predominando os
meses de agosto-outubro. Os frutos amadurecem em junho-julho (Lorenzi, 2002). A espécie G.
graciliflora é popularmente chamada de joão-mole, pau-mole e joão-dormido, atinge alturas
entre 4 e 5 m., possui inflorescências em cimeiras corimbiformes terminais com poucas flores de
cor esverdeada, os frutos são drupas de formato elipsóide, vermelho-vináceo, com polpa carnosa
contendo uma única sementes, produz moderada quantidade de sementes viáveis amplamente
dispersas pela avifauna, floresce durante os meses de agosto-setembro e os frutos amadurem de
outubro a novembro (Lorenzi, 2002). Maprounea guianensis é também chamada de bonifácio,
vaquinha, marmeleiro-do-campo, marmelinho–do-campo e milho-torrado, têm alturas entre 4 e
12 m., as inflorescências formam espigas axilares e terminais, com flores amarelo-creme, os
frutos são cápsulas globosas, deiscentes, abrindo-se em quatro valvas, com duas a quatro
sementes, floresce durante os meses de agosto-setembro, os frutos amadurecem em
setembro-outubro (Lorenzi, 2002). A espécie Matayba guianensis, da família Sapindaceae, possui hábito
arbóreo e é encontrada e áreas de cerrado e cerradão; o fruto é seco, deiscente, vermelho, do tipo
loculicida, as sementes têm de 8 a 16 mm e apresentam arilo de cor amarelada, a floração ocorre
ornitocoria (site: Rede de Sementes do Cerrado). A espécie Virola sebifera é popularmente
chamada de ucuúba-vermelha, gordura-de-virola ou ucuúba-do-cerrado, atingindo de 8 a 16 m.,
florescem de dezembro a fevereiro e abril a maio, a maturação dos frutos ocorre de julho a
setembro e os frutos abrem-se espontaneamente expondo as sementes de coloração vermelha,
devido à presença de arilo envolvendo-as (Lorenzi, 2002).
Todas as sementes foram obtidas nas áreas de estudo. Foram coletados frutos maduros,
frutos em maturação e sementes sob a planta-mãe. Os frutos em maturação foram expostos em
bandejas ao sol para que completassem a maturação permitindo a retirada manual das sementes e
para que os frutos deiscentes se abrissem e liberassem as sementes. Os frutos que continham
polpa foram expostos ao sol para desidratação, simulando o efeito do sol sobre estes quando
depositados no solo. As sementes que continham elaiossoma foram utilizadas com o elaiossoma.
A coleta das sementes e os experimentos foram realizados em 2006 e 2007, iniciando no
final da estação seca, em setembro, e estendendo-se até o final da estação chuvosa, em março,
em ambos os anos. Os testes com cada tipo de sementes foram realizados à medida que as
espécies arbóreas estavam frutificando nas áreas de estudo. Foram pesadas trinta sementes de
cada espécie arbórea em balança analítica (com precisão de quatro casas decimais) para
determinar o peso médio de cada tipo de semente.
Para a determinação das taxas de remoção por cada agente foram estabelecidos três
tratamentos distintos baseados na limitação do acesso por agentes removedores (formigas, aves e
roedores), metodologia adaptada de Horvitz & Schemske (1986). Um tratamento proporcionou o
acesso às sementes somente por formigas (TRAT.1) e foi composto por potes plásticos, de 250
ml, com três aberturas laterais eqüidistantes, de 1 cm2 cada. O segundo tratamento permitiu que
roedores e formigas (TRAT.2) tivessem acesso às sementes e foi estruturalmente idêntico ao
utilizado para formigas, mas foi utilizado um pote plástico maior, de 500 ml, com três abertura
laterais equidistantes de 4,5 x 4,5 cm, para que aves não conseguissem acessar o interior. Os
introduzidas nas laterais. As sementes foram depositadas no interior dos potes, diretamente sobre
o solo. E o terceiro tratamento em que todos os agentes tinham acesso (TRAT.3) foi constituído
por tampas plásticas de 15 cm de diâmetro onde foi colocado um pouco de terra para que as
sementes fossem depositadas, as tampas foram fixadas no solo por um prego. Em todos os
tratamentos foram expostas aos agentes removedores 20 sementes de cada espécie a ser testada
(Figuras 1).
Em cada fitofisionomia as estações foram distribuídas aleatoriamente distantes ao menos
30 metros umas das outras. Os tratamentos dentro de cada estação contiveram o mesmo tipo de
semente e foram distantes por aproximadamente 1 m um do outro. Cada um dos três tratamentos
continha 20 sementes da espécie testada, totalizando 1.200 sementes de cada espécie expostas
aos agentes em cada fitofisionomia. As sementes ficaram expostas aos agentes removedores por
96 horas e as estações em cada ponto foram supervisionadas ao final desse período. Para a
inferência das porcentagens de remoção por cada agente foi tomada a diferença nas taxas de
remoção entre os diferentes tratamentos, da seguinte forma:
remoção no TRAT. 1 = % de remoção de sementes por formigas.
remoção no TRAT. 3 – remoção no TRAT.2 = % de remoção por aves.
remoção no TRAT. 2 – remoção no TRAT. 1 = % de remoção por roedores.
Para determinar as diferenças nas taxas de remoção entre os tratamentos e entre os
diferentes ambientes utilizou-se análise de variância do tipo split-plot (SPSS 2000). Foi aplicada
a transformação por arcoseno das proporções de remoção para a normalização dos dados. A
2.3. Efeitos da remoção de sementes no recrutamento de plântulas
Este experimento foi realizado no cerrado denso para determinar a influência da remoção
de sementes por formigas, aves e roedores no recrutamento de plântulas. Para tanto foram
utilizados dois tratamentos: parcelas livres para formigas, aves e roedores e parcelas onde esses
agentes foram excluídos (Figuras 2 e 3). As parcelas foram arranjadas em pares, isto é formando
um bloco com uma parcela de cada tipo de tratamento lado a lado (Figura 4). No total foram
estabelecidos 30 blocos distantes entre si por 30 metros no mínimo.
As parcelas de exclusão constituíram-se de cercados confeccionados em chapas de zinco,
tendo 0,70 x 0,70 m e com 15 cm de altura, sendo que 5 cm ficaram enterrados no solo para
fixação do cercado nos pontos. Esses cercados tiveram toda a borda impregnada por uma resina
que impede o acesso de insetos via solo (Tanglefoot®) para limitar a entrada de formigas e foram
fechadas por tela metálica fina (orifícios de 1,5 x 1,5 cm) para impedir a entrada de roedores e
1A
1Aaves. As parcelas livres para os agentes removedores eram semelhantes às parcelas de exclusão,
a fim de se reproduzir as mesmas condições de sombreamento, mas possuíam três aberturas
retangulares de 7 x 10 cm em cada um dos lados. Essas aberturas permitiram a entrada de
formigas, aves e roedores, tanto pelas aberturas quanto por cima, já que não haverá tela fechando
a entrada. A tela no tratamento livre foi posicionada a aproximadamente dois metros do solo,
preza as árvores do entorno por arames metálicos, para que houvesse condições semelhantes de
oferta de serapilheira dentro de ambos as parcelas. Toda a vegetação rasteira que encostava nas
Para a determinação do recrutamento foram testadas sementes de Guapira graciliflora
(Mart.) Lundell (Nyctaginaceae) uma vez que esta é uma espécie comum na área de estudos e
também porque em estudos preliminares observei que possui boa taxa de germinação. Também
contabilizei o número de plântulas de outras espécies naturalmente emergentes em todas as
parcelas. Antes da deposição das sementes parte da serapilheira formada por folhas inteiras e
galhos foi retirada para que a germinação não fosse inibida (Hoffman, 1996). As sementes foram
depositadas, a distâncias iguais, dentro das parcelas, dos dois tratamentos, numa área de 0,50 x
0,50 m. Foram postas para germinar 10 sementes de G. graciliflora em cada uma das 60
parcelas. A localização de cada uma das sementes na parcela foi marcada com palito de madeira
graciliflora e das plântulas provenientes do recrutamento natural nas parcelas foi realizada
mensalmente de março a dezembro de 2007. Para analisar se houve diferenças no recrutamento
de plântulas entre as parcelas livres aos agentes removedores e parcelas de exclusão de agentes
utilizei um testet pareado.
2.4. Identificação das formigas removedoras de sementes
Para o levantamento das espécies de formigas que removem sementes, e para a
determinação do destino das sementes removidas por estas formigas, realizei observações focais
da atividade de remoção por formigas em pontos ao longo de um transecto de 100 m. Cada tipo
de semente foi testado separadamente e nos períodos diurno e noturno. Os pontos de observação
estavam distantes por 10 m um do outro, totalizando 10 pontos de observação na fitofisionomia
de cerrado (sentido restrito) denso. Em cada ponto foi colocado um pedaço de papel branco de
10 x 10 cm onde foram colocadas as sementes para exposição. A quantidade de sementes variou
de espécie para espécie, mantendo-se constante o volume ocupado sobre o papel. Assim foram
expostas 30 sementes de Maprounea guianensis, sete de Virola sebifera, 10 de Coussarea
hydrangeaefoliae 10 de Guapira graciliflora.Foram realizadas observações diurnas das 9:00 às
12:00h e noturnas das 19:00 às 21:00h. As observações diurnas foram realizadas em duas
etapas, com censos em apenas cinco pontos ao mesmo tempo para que se pudesse seguir as
formigas carregando sementes, marcando os sítios de deposição das sementes. Nas observações
noturnas foram observadas as 10 estações ao mesmo tempo, já que os sítios de deposição de
sementes não foram identificados devido à dificuldade de visualização das formigas
transportando sementes na serapilheira.
As sementes foram colocadas em cada ponto, e após 15 minutos de espera iniciavam-se
as observações. Em cada ponto foram feitas quatro observações, de 5 minutos durante o dia e de
espécie visitante para identificação e também foram medidas as distâncias a que as formigas
transportaram as sementes. A identificação dos locais de deposição das sementes foi realizada
com o intuito de se obter conhecimentos mais exatos sobre o papel das formigas como agentes
removedores das sementes testadas. Com isso verificou-se em visitas posteriores se havia
sementes das espécies expostas fora dos ninhos ou se havia plântulas das mesmas emergindo nos
3. RESULTADOS
3.1. Determinação das taxas de remoção de sementes
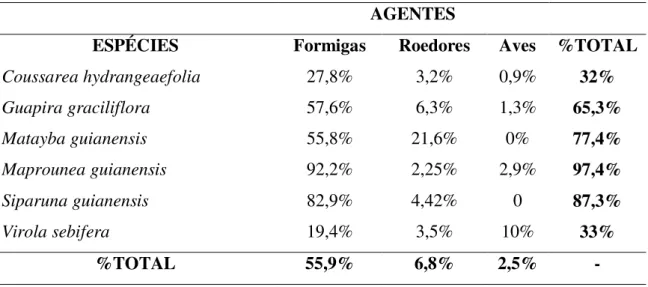
Do total de sementes expostas 57,8% foram removidas. As porcentagens de remoção
encontradas diferiram muito entre os tipos de sementes testadas e entre formigas, aves e
roedores, mas pouco entre os ambientes. Utilizando a subtração entre os tratamentos para inferir
sobre as porcentagens de remoção por cada agente, encontrei porcentagem total de remoção por
formigas de 56,6%; por roedores de 5,8% e por aves de 2,6%. As porcentagens totais de remoção
nas fitofisionomias foram de 66,1%; 55,6% e 55,5% para cerradão, cerrado sentido restrito e
cerrado denso, respectivamente. Entre as sementes utilizadas, a espécie Maprounea guianensis
foi a que teve a maior porcentagem de remoção, seguida por Siparuna guianensis, Matayba
guianensis, Guapira graciliflora, Coussarea hydrangeaefolia e Virola sebifera (Tabela 1). As
taxas de remoção de cada uma dessas espécies variou em cada fitofisionomia (Tabela 2) e de
acordo com cada um dos agentes analisados (Tabelas 1 e 2)
Tabela 1:Taxa de remoção (% do total removido em 96h nas fitofisionomias de cerrado sentido
restrito, cerrado denso e cerradão) por aves, formigas e roedores de sementes de seis espécies de
árvores do Cerrado, durantes as estações chuvosas dos anos de 2006 e 2007.
AGENTES
ESPÉCIES Formigas Roedores Aves %TOTAL
Coussarea hydrangeaefolia 27,8% 3,2% 0,9% 32%
Guapira graciliflora 57,6% 6,3% 1,3% 65,3%
Matayba guianensis 55,8% 21,6% 0% 77,4%
Maprounea guianensis 92,2% 2,25% 2,9% 97,4%
Siparuna guianensis 82,9% 4,42% 0 87,3%
Virola sebifera 19,4% 3,5% 10% 33%
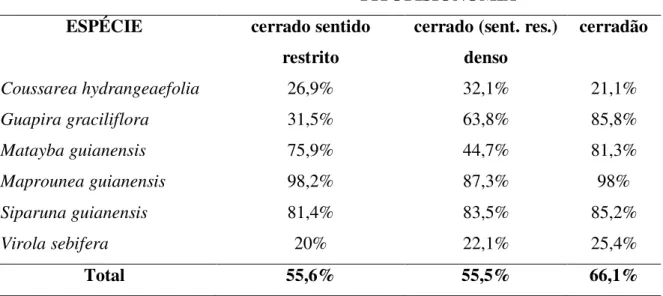
-Tabela 2: Taxa de remoção (% do total removido em 96h) de sementes de seis espécies arbóreas
em três diferentes fitofisionomias do Cerrado, durantes as estações chuvosas de 2006 e 2007.
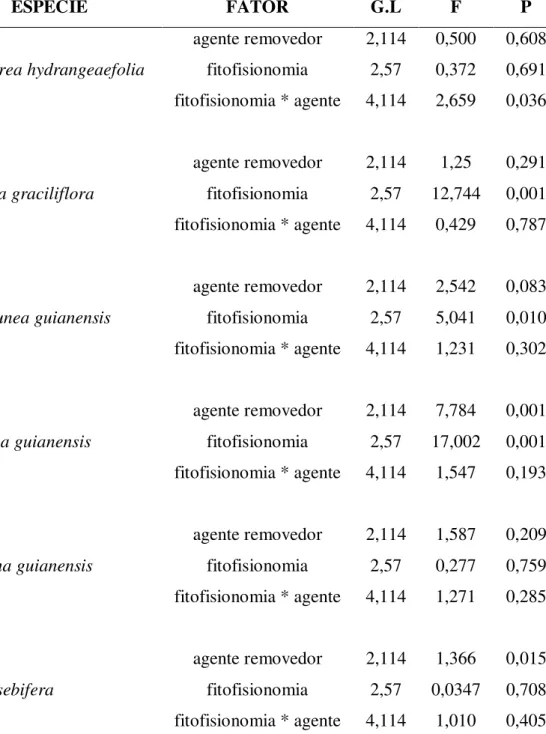
Para a maioria das espécies testadas, não houve diferenças significativas no número de
sementes removidas entre os três tratamentos, indicando que formigas foram o principal agente
removedor para a maioria das espécies testadas. As exceções a esse padrão foramVirola sebifera
e Matayba guianensis que diferiram significativamente quanto ao número de sementes
removidas em cada tratamento, tendo, além da remoção por formigas, sementes removidas por
aves e roedores, respectivamente (Figura 5). De um modo geral, não houve diferenças nas taxas
de remoção das espécies testadas entre as três fitofisionomias. Apenas Guapira graciliflora,
Maprounea guianensis e Matayba guianensis tiveram taxas de remoção significativamente
diferentes entre as fitofisionomias, sendo queMatayba guianensis eGuapira graciliflora tiveram
elevadas taxas de remoção no cerradão em comparação com as outras fitofisionomias, enquanto
Maprounea guianesis teve taxas de remoção no cerradão e no cerrado sentido restrito maiores
que no cerrado sentido restrito denso. Com exceção de Coussarea hydrangeaefolia, não houve
interação entre os efeitos da fitofisionomia e do agente removedor nas taxas de remoção de
FITOFISIONOMIA
ESPÉCIE cerrado sentido
restrito
cerrado (sent. res.) denso
cerradão
Coussarea hydrangeaefolia 26,9% 32,1% 21,1%
Guapira graciliflora 31,5% 63,8% 85,8%
Matayba guianensis 75,9% 44,7% 81,3%
Maprounea guianensis 98,2% 87,3% 98%
Siparuna guianensis 81,4% 83,5% 85,2%
Virola sebifera 20% 22,1% 25,4%
sementes (Tabela 3). ParaCoussarea hydrangeaefolia, houve maior remoção por vertebrados no
cerrado sentido restrito e por formigas no cerrado denso e no cerradão.
Tabela 3: Tabela de análise de variância mostrando os efeitos do tipo de agente removedor e da
fitofisionomia sobre as taxas de remoção de sementes de seis espécies arbóreas do Cerrado,
durante as estações chuvosas de 2006 e 2007.
ESPÉCIE FATOR G.L F P
Coussarea hydrangeaefolia
agente removedor 2,114 0,500 0,608 fitofisionomia 2,57 0,372 0,691 fitofisionomia * agente 4,114 2,659 0,036
Guapira graciliflora
agente removedor 2,114 1,25 0,291 fitofisionomia 2,57 12,744 0,001 fitofisionomia * agente 4,114 0,429 0,787
Maprounea guianensis
agente removedor 2,114 2,542 0,083 fitofisionomia 2,57 5,041 0,010 fitofisionomia * agente 4,114 1,231 0,302
Matayba guianensis
agente removedor 2,114 7,784 0,001 fitofisionomia 2,57 17,002 0,001 fitofisionomia * agente 4,114 1,547 0,193
Siparuna guianensis
agente removedor 2,114 1,587 0,209 fitofisionomia 2,57 0,277 0,759 fitofisionomia * agente 4,114 1,271 0,285
Virola sebifera
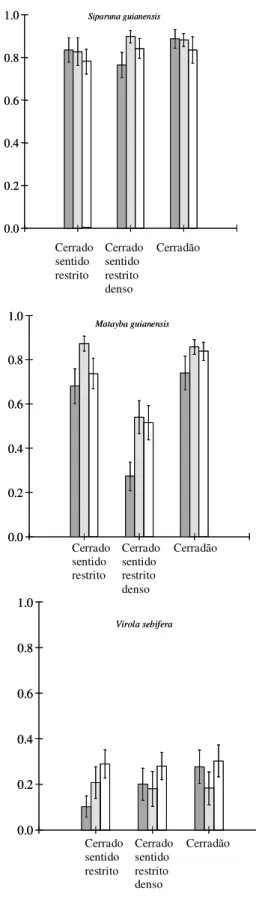
Figura 5:Taxa média de remoção (proporção do total removido em 96 h ± 1. E.P.M.) de sementes de seis espécies de plantas em três fitofisionomias do Cerrado por: formigas (barras cinza), por formigas e roedores (barras achuradas) e por formigas, aves e roedores (barras brancas).
0.0 0.2 0.4 0.6 0.8 1.0 Maprounea guainensis 0.0 0.2 0.4 0.6 0.8 1.0 Maprounea guainensis 0.0 0.2 0.4 0.6 0.8 1.0 Guapira graciliflora 0.0 0.2 0.4 0.6 0.8 1.0 Guapira graciliflora 0.0 0.2 0.4 0.6 0.8 1.0 Matayba guianensis 0.0 0.2 0.4 0.6 0.8 1.0 Matayba guianensis 0.0 0.2 0.4 0.6 0.8 1.0 Coussarea hydrangeaefolia 0.0 0.2 0.4 0.6 0.8 1.0 Coussarea hydrangeaefolia 0.0 0.2 0.4 0.6 0.8 1.0 Virola sebifera 0.0 0.2 0.4 0.6 0.8 1.0 Virola sebifera 0.0 0.2 0.4 0.6 0.8
1.0 Siparuna guianensis
0.0 0.2 0.4 0.6 0.8
1.0 Siparuna guianensis
As sementes que tiveram as maiores taxas de remoção foram as de Maprounea
guianensis e as que tiveram as menores taxas, foram as de Virola sebifera. Em todos os casos
formigas foram o principal agente removedor, com exceção de Virola sebifera e Matayba
guianensis, que tiveram remoção também por aves e roedores, respectivamente. O peso das
sementes foi importante na determinação das taxas de remoção das diferentes espécies arbóreas
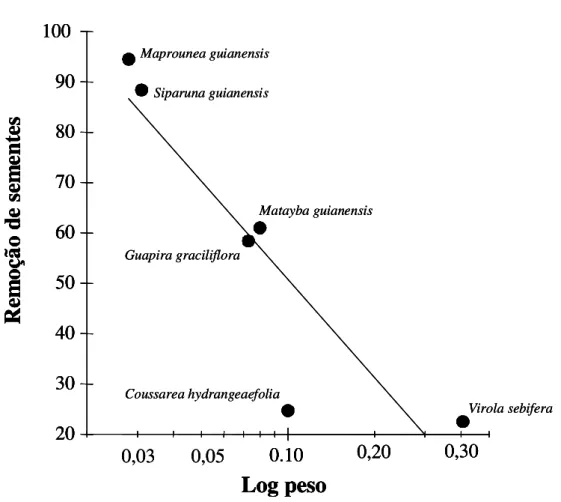
testadas (Figura 6), havendo uma relação negativa significativa entre o peso das sementes e a
porcentagem de remoção (F1,4 = 16,469; P < 0,05; r2 = 0,805). A relação é descrita pela equação:
y = 14,121 + 16,002 . log peso .
0,05 0,03 0.10
Log peso
20 30 40 50 60 70 80 90 100Rem
oçã
o d
e sem
ent
es
0,20 0,30 Virola sebifera Coussarea hydrangeaefolia Maprounea guianensis Siparuna guianensis Guapira graciliflora Matayba guianensis 0,05 0,03 0,05 0,03 0.10Log peso
20 30 40 50 60 70 80 90 100Rem
oçã
o d
e sem
ent
es
0,20 0,30 Virola sebifera Coussarea hydrangeaefolia Maprounea guianensis Siparuna guianensis Guapira graciliflora Matayba guianensis 0.10Log peso
20 30 40 50 60 70 80 90 100Rem
oçã
o d
e sem
ent
es
0,20 0,30 0.10Log peso
20 30 40 50 60 70 80 90 100Rem
oçã
o d
e sem
ent
es
0.10Log peso
20 30 40 50 60 70 80 90 100Rem
oçã
o d
e sem
ent
es
0,20 0,30 Virola sebifera Coussarea hydrangeaefolia Maprounea guianensis Siparuna guianensis Guapira graciliflora Matayba guianensisFigura 6: Relação entre o logaritmo do peso das sementes e a taxa de remoção de sementes
3.2. Efeitos da remoção de sementes no recrutamento de plântulas
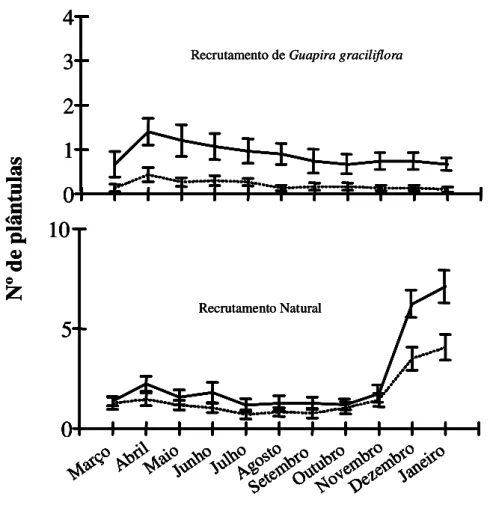
Houve diferença significativa no recrutamento de plântulas de Guapira graciliflora entre
as parcelas de exclusão de agentes e as parcelas livres a estes em todos os meses, exceto em
março, um mês após o plantio de G. graciliflora. Nas parcelas onde formigas, aves e roedores
foram excluídos, o número de plântulas de G. graciliflora foi significativamente maior que nas
parcelas onde estes agentes tiveram livre acesso (Tabela 4 e Figura 7). Não houve diferença
significativa no recrutamento de plântulas naturalmente emergentes entre as parcelas de exclusão
Meses
Nº
de
plâ
nt
ula
s
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
0
1
2
3
4
Recrutamento deGuapira graciliflora
Recrutamento Natural
Meses
Nº
de
plâ
nt
ula
s
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
0
1
2
3
4
Recrutamento deGuapira graciliflora
Recrutamento Natural
Meses
Nº
de
plâ
nt
ula
s
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
0
1
2
3
4
Recrutamento deGuapira graciliflora
Recrutamento Natural
Meses
Nº
de
plâ
nt
ula
s
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
0
1
2
3
4
Meses
Nº
de
plâ
nt
ula
s
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
0
1
2
3
4
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
0
1
2
3
4
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
0
1
2
3
4
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
Março AbrilMaioJunhoJulhoAgosto
Setem bro Outub ro Novem bro Deze mbro Janeiro
0
5
10
0
1
2
3
4
0
1
2
3
4
Recrutamento deGuapira graciliflora
Recrutamento Natural
Figura 7: Número total de plântulas de Guapira glaciliflora e de plântulas provenientes de
recrutamento natural nas parcelas sem removedores de sementes (linha cheia) e nas parcelas
com removedores de sementes (linha tracejada) entre março e dezembro de 2006 para G.
graciliflora e entre março e janeiro para recrutamento natural. As sementes (10 sementes por
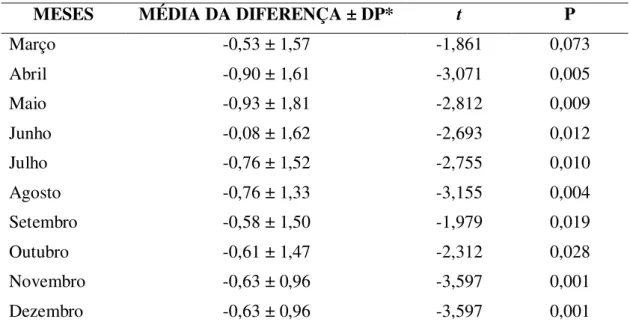
Tabela 4: Resultados dos testes t pareado comparando o número de plântulas de Guapira graciliflora entre as parcelas sem removedores de sementes e as parcelas com removedores de
sementes. Para cada mês foi comparado a diferença média no número de plântulas existentes em cada parcela.
MESES MÉDIA DA DIFERENÇA ± DP* t P
Março -0,53 ± 1,57 -1,861 0,073
Abril -0,90 ± 1,61 -3,071 0,005
Maio -0,93 ± 1,81 -2,812 0,009
Junho -0,08 ± 1,62 -2,693 0,012
Julho -0,76 ± 1,52 -2,755 0,010
Agosto -0,76 ± 1,33 -3,155 0,004 Setembro -0,58 ± 1,50 -1,979 0,019 Outubro -0,61 ± 1,47 -2,312 0,028 Novembro -0,63 ± 0,96 -3,597 0,001 Dezembro -0,63 ± 0,96 -3,597 0,001
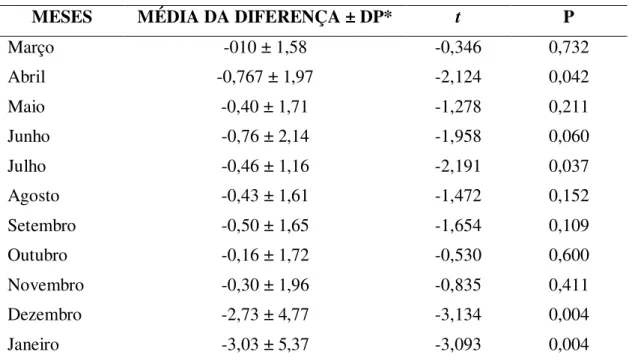
Tabela 5: Resultados dos testes t pareado comparando o número de plântulas naturalmente
emergentes entre as parcelas sem removedores de sementes e as parcelas com removedores de
sementes. Para cada mês foi comparado a diferença média no número de plântulas existentes em
cada parcela.
MESES MÉDIA DA DIFERENÇA ± DP* t P
Março -010 ± 1,58 -0,346 0,732
Abril -0,767 ± 1,97 -2,124 0,042
Maio -0,40 ± 1,71 -1,278 0,211
Junho -0,76 ± 2,14 -1,958 0,060
Julho -0,46 ± 1,16 -2,191 0,037
Agosto -0,43 ± 1,61 -1,472 0,152 Setembro -0,50 ± 1,65 -1,654 0,109 Outubro -0,16 ± 1,72 -0,530 0,600 Novembro -0,30 ± 1,96 -0,835 0,411 Dezembro -2,73 ± 4,77 -3,134 0,004 Janeiro -3,03 ± 5,37 -3,093 0,004
3.3. Identificação das formigas removedoras de sementes
As espécies diferiram na forma de interação com as sementes, variando de acordo com as
características morfológicas das sementes (Tabela 6). O transporte das sementes foi feito quase
sempre para ninhos próximos aos pontos de observação. As formigas de pequeno porte como
Solenopsis e Linepithema normalmente foram encontradas consumindo o arilo das sementes ou
retirando partes dele e transportando. As espécies que recrutam grande número de operárias
como as do gêneroPheidole e as formigas de maior tamanho corporal como as dos gênerosAtta,
Ectatomma e Pachycondyla removeram ativamente as sementes para o interior de seus ninhos,
sendo Atta o gênero que removeu mais Guapira graciliflora e Ectatomma o que removeu mais
Maprounea guianensis (Tabela 6). O tempo gasto para identificar a presença das sementes de
Maprounea guianensis eVirola sebifera, que contém elaiossoma e arilo, respectivamente, foi de
menos de dez minutos em média. Já para encontrar as sementes de Guapira graciliflora e
Coussarea hydrangeaefolia, em que a parte atrativa é a polpa dos frutos, as formigas levaram 20
min e 15 min, respectivamente. Durante as observações marquei nove ninhos de formigas do
gênero Pheidole, dois ninhos do gênero Ectatomma e dois do gênero Atta.A distância entre os
pontos estabelecidos e os ninhos onde foram depositadas as sementes variou de 0,45 m até 14 m.
Os ninhos das formigas do gênero Pheidole estavam sempre próximos aos pontos de observação
(0,45m ± 1,30m), os ninhos de Ectatomma encontravam-se a 0,75 e 0,77 m dos pontos e os
ninhos deAtta tiveram distâncias variando entre 1,2 a 14 m. As sementes que foram depositadas
não foram vistas no material de descarte nos entornos dos ninhos, nem foram observadas
Tabela 6: Gêneros de formigas observados no levantamento de formigas que interagem com
sementes, número de espécies de cada gênero e forma com que esses gêneros interagiram com as
sementes. Os gêneros marcados com (*) foram os mais comuns interagindo com o respectivo
tipo de semente. Os números entre parênteses correspondem ao número de espécies de cada
gênero registrado por tipo de semente.
ESPÉCIE DE
PLANTA GÊNEROS QUEREMOVERAM SEMENTES GÊNEROS QUE REMOVERAM ELAIOSSOMA OU POLPA APENAS APROXIMAÇÃO
Maprounea guianensis Pheidole (5) *
Ectatomma(2)* Solenopsis (1)* Atta(1) Pachycondyla (1) Solenopsis (1)* Linepithema (1) Trachymyrmex(1) Camponotus(2) Atta(1)
Cephalotes(1 )
Virola sebifera Pheidole (4)*
Ectatomma(1)*
Attini não identificada
Solenopsis (2)*
Linepithema(1)
Camponotus (3)
Pseudomyrmex(1)
Guapira graciliflora Pheidole(3)*
Atta(1)*
Attini não identificada*
Ectatomma(2) Solenopsis (3) Crematogaster(1)* Camponotus(2) Linepithema(1) Coussarea hydrangeaefolia Pheidole(4)* Atta(1)*
Attini não identificada*
Ectatomma(1)
Solenopsis (1)*
Crematogaster(1)
4. DISCUSSÃO
4.1. Remoção de sementes do Cerrado por formigas, aves e roedores
As formigas foram o principal grupo removedor de sementes neste estudo, com altas
taxas de remoção em todas as fitofisionomias. Diferente do encontrado aqui, em estudo sobre
remoção e predação de sementes realizado também em área savânica na Venezuela, Pérezet. al
(2006) verificaram que os roedores foram o principal grupo predador de sementes com 58,7% de
remoção, enquanto as formigas removeram apenas 22,6% das sementes expostas e aves 18,6%.
Em floresta secundária no México formigas removeram cerca de 89,2% das sementes expostas
em 90 minutos, com vertebrados removendo taxa não significativa (Horvitz & Schemske, 1986).
Em áreas de campo na Argentina formigas foram, no geral, o principal consumidor de sementes,
mas houve variação nas taxas de remoção entre as estações do ano com formigas sendo mais
importantes no verão e na primavera e vertebrados no outono e inverno (Casenaveet. al., 1998).
Em área semi-árida do Chile formigas e aves foram os principais consumidores de sementes, mas
com atividade de remoção por formigas maior no verão (Kelt et. al., 2004). Na Austrália, em
floresta de eucaliptos as formigas também tiveram taxas de remoção de sementes maiores que de
vertebrados (Andersen & Ashton, 1985). Como o verificado aqui, em área de cerrado sentido
restrito formigas removeram mais sementes que vertebrados (Christianini et. al., 2007). As
interações entre formigas e sementes vêm sendo foco de estudos em diversos biomas brasileiros
nos últimos anos. Na Mata Atlântica várias espécies de formigas interagem com sementes
consumindo polpas e elaiossomas, transportando as sementes para o interior de seus ninhos (Pizo
& Oliveira, 2001, 2000, 1998). Estas interações entre formigas e sementes também foram
documentadas por Leal & Oliveira (1998) em áreas de Cerrado e por Leal (2003) na Caatinga.
Diferentemente das altas taxas de remoção encontradas para formigas, houve baixa taxa
forrmigas levam menos tempo para encontrar as sementes. Isto porque, normalmente, as
primeiras formigas a visitar as sementes possuem ninhos próximos aos pontos de deposição
destas. As primeiras formigas a encontrarem as sementes transportam grande parte destas antes
mesmo que roedores e aves encontrem. Estudo sobre remoção de sementes por formigas em
áreas de Cerrado mostrou médias das distâncias de dispersão de sementes variando entre 1,46 e
17,7m (Christianini et. al. 2007), distâncias relativamente pequenas, o que evidencia que os
primeiros agentes a encontrar as sementes são mesmo as formigas com ninhos próximos às
sementes.
Mas estas taxas podem ter sido causadas por outros dois fatores que podem estar ligados:
a baixa densidade de roedores e aves na área de estudo e a perturbação. A Estação Ecológica do
Panga é hoje um remanescente conservado de Cerrado cercado por extensas áreas de exploração
agropecuária, sendo que a reserva era explorada para esses fins há mais de 20 anos atrás, antes de
se tornar uma RPPN, por isso a área pode sofrer impactos pelo uso intensivo dos entorno para
estas práticas.
Essa perturbação pode ser evidenciada após levantamento de pequenos mamíferos
realizado na área de 2005 a 2006 (Apêndice 1), onde constatou-se baixa densidade de roedores.
A taxa de captura de 1,58% encontrada aqui é considerada baixíssima, assim como a taxa de
4,8% encontrada por Pizo & Vieira (2004) em fragmento de Mata Atlântica perturbado. Em
contrapartida, Vieira (1999) encontrou taxas de captura de 6 a 15% em áreas de Mata atlântica
não perturbada, mostrando que a perturbação é um fator que interfere negativamente na
densidade de roedores. A dependência da densidade de roedores para a determinação das taxas
de remoção foi mostrada no estudo de Ostfeld et. al.(1997), onde a baixa densidade de roedores
foi um dos fatores causadores de baixas taxas de remoção de sementes por roedores.
Para aves, a perturbação também é um fator importante, já que a redução de áreas
naturais conservadas fora da reserva limita a movimentação das epécies de aves, podendo
podem ser consideradas como causadoras das baixas taxa de remoção, já que das seis espécies
utilizadas quatro são ornitocóricas (Matayba guianensis, Guapira graciliflora, Coussarea
hydrangeaefolie Virola sebifera).
4.1. Variações espaciais na remoção de sementes
Houve diferença nas taxas de remoção de sementes entre fitofisionomias para três das
seis espécies testadas. De modo similar, em floresta secundária no México, a remoção de
sementes variou entre diferentes áreas, e a diferença encontrada deveu-se à presença ou ausência
de espécies de Pachycondyla nas áreas (Horvitz & Schemske, 1986). Em áreas de campo na
Argentina, a diferença na remoção de sementes entre diferentes áreas foi significativa apenas no
verão, havendo uma interação entre localidades, sazonalidade e agente removedor (Casenave et.
al., 1998).
As diferenças nas taxas de remoção entre ambientes encontradas para Guapira
graciliflora, Matayba guianensiseMaprounea guianensispodem ser resultantes de dois fatores.
Pelo fato dessas espécies terem altas taxas de remoção por formigas, devido à presença de
substâncias e apêndices atrativos (polpa e elaiossoma), as diferenças entre as fitofisionomias
podem refletir diferenças na atividade de formigas entre essas áreas ou, ainda, diferenças na
composição de espécies de formigas que utilizam diferentemente cada semente. As taxas de
remoção foram maiores onde estas espécies são menos abundantes, como no cerradão,
fitofisionomia que difere em composição de espécies arbóreas dos cerrados sentido restrito e
cerrado sentido restrito denso. Outra possível explicação, então, é a maior atratividade dessas
4.2. Variações interespecíficas na remoção de sementes
Houve grande variação nas taxas de remoção entre os seis tipos de sementes testadas. O
peso e a presença de substâncias atrativas foram importantes na definição das taxas de remoção.
Sementes menores como as de Maprounea guianesis, Siparuna guainensis e Guapira
graciliflora foram rapidamente e quase que exclusivamente removidas por formigas. Já sementes
de maior porte como Virola sebifera e Matayba guianensis além da remoção por formigas,
também foram removidas por aves e roedores. As taxas de remoção de sementes de cinco
espécies de Cerrado foram positivamente correlacionadas com a massa de substâncias lipidícas
(i. e. elaiossoma) presentes nas sementes testadas (Christianini et. al., 2007). O trabalho de
Andersen & Ashton. (1985) mostrou taxas de remoção variando de acordo com a presença de
elaiossoma, com 95% das sementes que continham elaiossoma sendo removidas. O peso das
sementes e a presença de elaiossoma foram fatores importantes na determinação de taxas de
remoção de sementes no estudo realizado por Pèrez et. al. (2006), onde formigas preferiram
sementes pequenas e que continham elaiossoma, enquanto roedores removeram sementes
maiores e aves sementes menores, mas sem elaiossoma. Leal & Oliveira (2007) viram que o
tamanho das sementes influenciou não apenas a taxa de remoção como também a distância a que
sementes são transportadas pelas formigas.
Os resultados obtidos no presente estudo, assim como os resultados obtidos em outros
estudos mostram que o peso das sementes e a presença de elaiossoma são importantes na
determinação das taxas de remoção de sementes. A determinação da importância de cada um
desses fatores depende de uma análise mais cuidadosa caso a caso. Quando verificamos as taxas
de remoção por formigas de duas sementes de baixo peso como Maprounea guianensis (0,0307
g) e Siparuna guianensis (0,031 g) verifica-se que M. guianensis, que contém elaiossoma, teve
taxa de remoção de 92,2%, enquanto S. guianensis, que tem praticamente o mesmo peso, mas
não contém elaiossoma, teve taxa de remoção menor (86,9%). Nesse caso o elaiossoma foi o
contrapartida, quando analisamos o caso de sementes maiores que contém arilo como Virola
sebifera (0,406 g) eMatayba guianensis(0,067 g), nota-se que as taxas de remoção por formigas
foram bem menores se comparadas com as obtidas para sementes pequenas e que além da
remoção por formigas houve alguma remoção por vertebrados, mostrando que aqui o
determinante das taxas foi o peso das sementes que causou menor remoção por formigas apesar
da presença do elaiossoma como atrativo em ambas as sementes. Muitas vezes, durante a
contagem de sementes nas estações, as formigas foram vistas consumindo o elaiossoma dessas
sementes de maior porte, mas o transporte das sementes foi baixo devido ao peso elevado das
mesmas.
4.3. Destino das sementes removidas por formigas: Predação ou dispersão
secundária?
Através das observações focais das interações das formigas com as sementes e da
marcação de alguns ninhos onde as sementes foram depositadas, pude notar que nos arredores
dos ninhos marcados não havia plântulas. Em observações posteriores, também não observei
presença de plântulas emergindo nos ninhos, bem como a presença de sementes descartadas
intactas, o que caracteriza atividade de predação de sementes e não de dispersão secundária.
Foram marcados apenas ninhos dos gêneros Pheidole, Ectatomma e Atta. As formigas dos
gêneros Solenopsis, Camponotus e Linepithema, que removeram elaiossoma mas não
transportaram sementes, ou, na tentativa de transportar, deixaram sementes no caminho até seus
ninhos, podem ter papel de dispersoras secundárias, sendo mais comum não transportarem as
sementes e sim partes do elaiossoma. Em estudo realizado por Levey & Byrne (1993), também
não foram registradas sementes descartadas no entorno dos ninhos de formigas do gênero
Pheidole, sendo estas consideradas predadoras de sementes. Em trabalho realizado aqui na
laevigata e fora destes, verificando que não há diferenças significativas na densidade de
plântulas sobre os ninhos e no entorno dos ninhos. Isto mostra que os solos dos ninhos nem
sempre são sítios favoráveis para a germinação de sementes (Perin & Vasconcelos, dados não
publicados). Silva et. al. (2007) viram que formigas do Atta sexdens carregam sementes para
seus ninhos e depositam fora destes após limpa-las, mas as plântulas emergentes são cortadas por
estas formigas, causando baixo recrutamento de plântulas sobre os ninhos.
As espécies de formigas registradas interagindo com as sementes foram as dos gêneros
Atta, Cephalottes, Pseudomyrmex, Pachycondyla, Ectatomma, Camponotus, Crematogaster,
Trachymyrmex, Solenopsis, Pheidole e Linepithema. As formigas do gênero Pheidole,
Camponotus e Atta foram registradas interagindo com todas as sementes testadas. Mas as
formigas que efetivamente transportaram sementes para seus ninhos foram as de maior porte
corporal, como Atta e Ectatomma, além dos gêneros que recrutam muitas operárias como
Pheidole. Os outros gêneros de menor porte interagiram removendo elaiossoma das sementes,
mas não transportando estas para o interior de seus ninhos.
As formigas interagiram de diferentes formas com as sementes. As sementes de menor
peso como Maprounea guianensis, Guapira graciliflora e Coussares hydrangeaefolia foram
rapidamente transportadas enquanto Virola sebifera, a de maior peso, demorou mais tempo para
ter suas sementes transportadas. Em casos como o de Coussarea hydrangeaefolia, essas formigas
normalmente transportaram frutos para seus ninhos, retiraram a polpa para o cultivo do fungo de
que se alimentam e depois descartaram as sementes limpas nas lixeiras dos ninhos intactas (A. V.
Ferreira, observações pessoais). Nesses casos, as sementes poderão germinar normalmente,
caracterizando a atividade dispersora do gênero Atta. Nas observações focais de remoção de
Coussarea hydrangeaefolia, pôde-se notar que, apesar do pequeno peso das sementes, não houve
elevada taxa de remoção, possivelmente porque as sementes desta espécie não possuem nenhum
Considerando a ecologia das espécies arbóreas utilizadas nos testes, notamos que
Guapira graciliflora, Coussarea hydrangeaefolia, Matayba guianensis e Virola sebifera são
espécies primariamente dispersas por aves, passando intactas pelo trato digestivo destas e sendo
descartadas intactas nas fezes (A. V. Ferrerira, observações pessoais), sugerindo que, muitas
vezes, as formigas podem estar transportando essas sementes após a dispersão primária nas fezes
de aves, não tendo, portanto, papel na otimização das condições de germinação através da
eliminação dos arilos e polpas. JáMaprounea guianensis, espécie da família Euphorbiaceae, que
possui sementes mirmecocóricas, pode ter seu recrutamento beneficiado no caso da remoção por
formigas de pequeno porte como as do gênero Pheidole e Solenopsis, que utilizariam o
elaiossoma e possivelmente descartariam essas sementes limpas nos arredores dos ninhos,
embora isto não tenha sido observado em campo no estudo. Mas, por outro lado, podem ter seu
potencial de germinação destruído pela ação predatória de formigas de grande porte como as dos
gênerosEctatomma, PachycondylaeAtta, registradas removendo sementes dessa espécie na área
de estudo. Enquanto os principais gêneros de formigas registradas interagindo com sementes
aqui foram as de menor porte, como Pheidole e Solenopsis, trabalhos nas áreas de restinga e
Mata Atlântica, realizados por Passos & Oliveira (2003) e por Pizo & Oliveira (1998),
respectivamente, os principais gêneros registrados interagindo com as sementes foram os de
maior tamanho corporal Pachycondyla e Ectatomma. Formigas do gênero Ectatomma foram
registradas por Leal & Oliveira (1998), em áreas de Cerrado, removendo cerca de 50% das
sementes deVirola sebifera, juntamente com formigas do gêneroOdontomachus, não registradas
aqui. Analisando as interações entre formigas e sementes de espécies arbóreas de Cerrado,
Christianiniet al (2007) registraram 27 espécies de formigas interagindo com as sementes. Nesse
estudo, cada espécie de formigas mostra preferência e comportamento distintos para cada tipo de
semente exposta, com destaque para Pheidole que apresentou 10 espécies interagindo com os
nove tipos de sementes expostas. Os outros gêneros registrados no estudo foram Atta, Azteca,
Pachycondyla, Solenopsis e Wasmania. Corroborando com o que verifiquei aqui, em estudo
realizado em área de floresta secundária por Horvitz & Schemske (1986), as primeiras formigas
a interagirem com sementes foram as Pheidole, seguidas por outros gêneros que interagem após
a chegada destas. Nesse mesmo estudo, 77% das sementes deCalathea ovadensis (Marantaceae)
foram transportadas por formigas, sendo que a espécie mais frequentemente encontrada nas
sementes foi Pachycondyla harpax, contribuindo com 54% das interações e transportando as
sementes a distâcia máxima de 76 cm. Outras formigas registradas no estudo de Horvitz &
Schemske (1986) foram Pheidole sp, Solenopsis geminata e Wasmania auropunctata que
recrutaram grande número de operárias.
4.4. Impacto da remoção de sementes no recrutamento de plântulas
O recrutamento de plântulas de Guapira graciliflora nas parcelas abertas a formigas,
roedores e aves teve redução de 83%, enquanto nas parcelas com exclusão desses agentes apenas
30,3% das sementes introduzidas não germinaram. O impacto dos removedores sobre o
recrutamento de plântulas naturalmente emergidas só foi significativo a partir do mês de
novembro, quando houve dispersão massiva de frutos e sementes e intensificação das chuvas. A
maioria dos trabalhos envolvendo efeitos de granivoria sobre o recrutamento de plântulas trata
dos efeitos de vertebrados sobre a vegetação, principalmente do efeito de roedores. A exclusão
de roedores causou aumento no recrutamento de plântulas em áreas de dunas e de pastagens
(Maron & Simms, 1997)
Tendo em vista as observações realizadas acerca das interações entre formigas e
sementes, podemos concluir que estas como mais importantes removedoras e predadoras de
sementes causam impacto sobre o recrutamento de plântulas. Esse impacto poderá ser positivo
quando formigas de menor porte, ao tentar transportar essas sementes, deixam cair parte delas
maior competição como as áreas sob a planta-mãe (Janzen, 1970). Por outro lado, esse efeito
pode ser negativo, quando as formigas atuam como predadoras de sementes, removendo estas
sementes e indisponibilizando estas para o recrutamento através da deposição no interior de
ninhos ou quando utilizam partes dessas sementes como alimento. Os predadores podem ser
vistos, então, como agentes que afetam diretamente as populações de plantas em áreas de
Cerrado e os dispersores teriam efeito positivo indireto sobre essas populações. Tendo em vista
que os efeitos de consumidores de sementes devem ser mais importantes para populações de
plantas que não têm seu recrutamento diminuído pela ausência de microhabitats e competição, há
necessidade de estudos mais detalhados sobre os efeitos de consumidores de sementes em longo
prazo para essas populações. Embora para Guapira graciliflora tenha sido observado um
aumento significativo no recrutamento de plântulas na ausência de predadores, não está claro se
este aumento resultará num maior número de indivíduos adultos na população. Essa é uma
questão interessante para o entendimento da dinâmica de comunidades vegetais, principalmente
para os ecossistemas de Cerrado, onde parece haver uma dinâmica sucessional entre as diferentes
fitofisionomias pela atuação de fatores abióticos como o fogo e bióticos como a predação,
5. CONCLUSÕES
Os agentes removedores e os tipos de sementes foram os fatores mais importantes na
determinação das taxas de remoção. As formigas foram o principal grupo removedor de
sementes na área de estudo, sendo as formigas o grupo que interagiu com as sementes na maioria
das vezes. As formigas atuam como dispersoras e como predadoras de sementes, muitas vezes
levam as sementes para o interior dos ninhos após já serem utilizadas por outros agentes, como
as aves, por exemplo. As diferentes fitofisionomias de Cerrado foram fator determinante de
diferenças nas taxas de remoção apenas para três das seis espécies testadas. O peso e a presença
de elaiossoma explicaram os padrões de remoção encontrados. Depois do peso das sementes, um
fator importante na remoção foi a presença de elaiossoma e arilo. Portanto, os fatores que
influenciaram os padrões de remoção de sementes foram, primeiramente, o peso das sementes, e
depois a presença de elaiossoma.
As formigas que mais frequentemente foram registradas interagindo com sementes na
área de estudo foram as dos gêneros Pheidole, Solenopsis e Atta. A remoção por formigas pode
ter efeitos positivos indiretos para a comunidade quando há dispersão para locais favoráveis à
germinação, rearranjo espacial da deposição, consumo de elaiossoma e polpa das sementes. Em
contrapartida, as formigas podem causar efeitos negativos diretos sobre comunidades quando
atuam como predadoras de sementes, reduzindo o recrutamento pela redução da densidade de
sementes de algumas populações preferencialmente consumidas devido à presença de atrativos.
No presente estudo, os efeitos negativos da remoção por formigas parecem ser mais
proeminentes, tendo em vista que o impacto da ausência desses agentes removedores foi
positivamente significativo para o recrutamento de plântulas. Aqui, estes agentes podem estar
atuando como reguladores da densidade de plântulas de espécies preferencialmente removidas,
Este estudo trouxe uma visão quantitativa dos impactos da remoção de sementes no
recrutamento em áreas de Cerrado. São necessários estudos mais aprofundados sobre o impacto
da remoção/predação de sementes em nível de populações individuais de plantas para que se
possa prever qual a magnitude desses efeitos em nível estrutural das comunidades vegetais.
Quando se analisa a função dos agentes granívoros na dinâmica de populações vegetais, deve-se
tomar cuidado com as generalizações, tendo em vista que cada estudo é pontual e diz respeito à
realidade ecológica de um ecossistema com suas particularidades. Esses efeitos vistos aqui
certamente diferirão de população para população vegetal, sendo positivo para umas e negativo
para outras. Só com o entendimento parte a parte poderemos chegar a evidências dos reais
6. REFERÊNCIAS BIBLIOGRÁFICAS
ANDERSEN, A. N. 1989. How important is seed predation to recruitment in stable populations of long-lived perennials. Oecologia. 81: 310-315.
ANDERSEN, A. N.; ASHTON, D. H. 1985. Rates of seed removal by ants at heath and woodland sites in southeastern Australia. Australian Journal of Ecology. 10: 381-390.
CASENAVE, J. L.; CUETO, V. R.; MARONE, L. 1998. Granivory in the Monte Desert, Argentina: Is it less intense in other arid zones of the word?. Global Ecology and Biogeography Letters. 7 (3): 197-204.
CHRISTIANINI, A. V.; MAYHÉ-NUNES, A. J.; OLIVEIRA, P. S. 2007. The role of ants in the removal of non-myrmecochorous diaspores and seed germination in a neotropical savanna. Journal of Tropical Ecology. V. 23. p. 343-351.
CRAWLEY, M. J. 1992. Seed predator and plant population dynamics seeds, the ecology of regeneration in plant communities. CAB International, Wallingford, p. 157-192.
EDWARDS, G. R.; CRAWLEY, M.J. Rodent seed predation and seedling recruitment in mesic grassland. Oecologia, v. 118, p. 288-296. 1999.
GUTIÈRREZ, J.R.; MESERVE, P. L.; HERRERA, S.; CONTRERAS, L.C.; JAKSIC, F. M. 1997. Effects of small mammals and vertebrate predators on vegetation in the Chilean semiarid zone. Oecologia. 109: 398-406.
HOFFMANN, W. A. 1996. The effects of fire and cover on seedling establishment in a neotropical savanna. Journal of Ecology. .84: 383-393.
HULME, P. E. 1998. Post-dispersal seed predation: consequences for plant demography and evolution. Perspectives in Plant Ecology, Evolution and Systematics. 1: 32-46.
JANZEN, D. H. 1971. Seed predation by animals. Annual Review of Ecology and Systematics. 2: 465-492.
JANZEN, D. H. 1970. Herbivores and the number of tree species in tropical forests. American Naturalist. 104: 501-529.
KELT, D. A.; MESERVE, P. L.; GUTIÈRREZ, J. R. 2004. Seed removal by small mammals, birds and ants in semi-arid Chile, and comparison with other systems. Journal of Biogeography. 31: 931-942.
LEAL, I. R. Dispersão de sementes por formigas na Caatinga. In: LEAL, I. R.; TABARELLI, M.; SILVA, J. M. C. Ecologia e Conservação da Caatinga. Pernambuco: Universitária. 2003.
LEAL, I. L.; OLIVEIRA, P. S. 1998. Interactions between fungus-grrowing ants (Attini), fruits and seeds in Cerrado vegetation in southeast Brazil. Biotropica. 30 (2): 170. 1998.
LEVEY, D. J.; BYRNE, M. M. 1993. Complex ant-plant interactions: rain Forest ants as secondary disperses and posdispersal seed predators. Ecology. 74: 1802-1812.
LORENZI, H. Árvores Brasileiras: manual de identificação e cultivo de plantas arbóreas nativas do Brasil. 4 ed. V. 1 e 2. Nova Odessa-SP: Instituto Plantarum. 2002.
MARON, J. L.; SIMMS, E. L. 1997. Effect of seed predation on seed bank size and sedling recruitment of bush lupine (Lupinus arboreus). Oecologia. 111: 76-83.
MARON, J. L.; SIMMS, E. 2001. Rodent-limited establishment of bush lupine:fiel experiments on the cumulative effects of granivory. Journal of Ecology. 89: 578-588.