THE CORPUS CALLOSUM AND ITS ABNORMALITIES IN CATS AND DOGS Three distinct clinical presentations
Dissertação de Mestrado Integrado em Medicina Veterinária
Francisca Soares Marques de Almeida
Orientador
Professor Doutor Artur Severo Proença Varejão Coorientador
Dr. Xavier Raurell Ribó
i
THE CORPUS CALLOSUM AND ITS ABNORMALITIES IN CATS AND DOGS Three distinct clinical presentations
Dissertação de Mestrado Integrado em Medicina Veterinária
Francisca Soares Marques de Almeida
Orientador Professor Doutor Artur Severo Proença Varejão Coorientador Dr. Xavier Raurell Ribó
Composição do Júri:
Presidente: Doutor Celso Alexandre Sá Santos Arguente: Doutora Paula Cristina Avelar Rodrigues
Arguente: Dr. João Pedro de Almeida Carneiro Silva Machado
iii Nome: Francisca Soares Marques de Almeida C.C.: 13933050
Telemóvel: (+351) 936 188 064
Correio eletrónico: [email protected]
Designação do Mestrado: Mestrado Integrado em Medicina Veterinária
Título da dissertação de Mestrado em Medicina Veterinária: The Corpus Callosum and its Abnormalities in cats and dogs: Three distinct clinical presentations
Orientador: Professor Doutor Artur Severo Proença Varejão Coorientador: Dr. Xavier Raurell Ribó
Ano de Conclusão: 2020
Declaro que esta dissertação de mestrado é resultado da minha pesquisa e trabalho pessoal e das orientações dos meus supervisores. O conteúdo é original e as fontes consultadas estão devidamente mencionadas no texto e na bibliografia final. Declaro ainda que este trabalho não foi apresentado em nenhuma outra instituição para obtenção de qualquer grau académico.
Vila Real, Maio de 2020
v
“The face predicts the brain
The image predicts its function”
vii
Em primeiro lugar gostaria de agradecer ao meu orientador Professor Doutor Artur Varejão não só pela orientação desta dissertação, mas também pelo seu exemplar trabalho como professor que me instigou o gosto pela neurologia e a vontade de aprender sempre mais. Obrigada por confiar em mim.
Em segundo lugar quero agradecer ao meu coorientador Dr. Xavier Raurell Ribó. Obrigada pela sugestão e incentivo ao desenvolvimento deste tema que me desafiou e maravilhou em vários aspetos e por todos os momentos de amizade e aprendizagem que me proporcionou no Hospital Veterinari Mollins. À Carme Centelles e Gemma Felliz um especial agradecimento por me terem acolhido como parte da equipa de neurologia e partilhado os vossos conhecimentos. Agradeço também à Angeles Zamora pelo profissionalismo na análise detalhada das imagens de ressonância magnética. Por fim, quero agradecer a todos outros membros da equipa que constituí o Hospital Veterinari Mollins que contribuíram de alguma forma para a minha aprendizagem.
Obrigada a todos os amigos, professores, colegas e médicos veterinários que durante a minha vida e percurso académico contribuíram para a minha formação profissional e crescimento pessoal. Todas as etapas foram importantes de forma singular e conduziram-me até onde me encontro hoje. Agradeço também à pequena (grande) cidade de Vila Real e à Universidade de Trás-os-Montes e Alto Douro por me darem a conhecer outros costumes, a vida académica e me formarem como futura médica veterinária.
Ao meu grupo de amigos de Aveiro, Chafarrica, agradeço as vivências ao longo destes mais de dez anos que me ensinaram muito sobre amizade e me fizeram crescer como pessoa. Em especial, um obrigado às minhas meninas Mariana Costa, Bárbara Rosário e Ana Luísa Carvalho, mesmo afastadas a amizade prevalece.
Um grande obrigado aos Manos e Manas. Vocês foram e são a minha segunda família de Vila Real, o melhor grupo de amigos que poderia ter desejado e tive a sorte de conhecer. A universidade não teria sido o mesmo sem todos vocês. Obrigada do fundo do coração, sei que vos levo comigo para a vida.
Um especial obrigado ao Hyan Aguiar e Silva pela amizade sincera, que evoluiu inesperada e surpreendente de forma a ser, atualmente, uma das pessoas mais importantes na minha vida. O teu apoio durante a realização deste trabalho foi fundamental e insubstituível. Obrigada por toda a motivação, compreensão, ataques de riso e por acreditares sempre em mim.
viii
as três juntas como sempre, depois de tanto riso, estudo, passeios e viagens pelo país, tantas aventuras, tanta amizade. Graças a vocês entendi o significado da expressão ‘é na Universidade que se vivem os melhores anos das nossas vidas’. Sei que, no entanto, estes foram apenas o início e que o nosso melhor estará ainda por vir. Obrigada por somarem tanta felicidade à minha vida e por serem um porto seguro de confiança e riso todos os dias. Obrigada por me terem tornado numa pessoa melhor e mais forte.
Aos meus pais e irmã não existe maneira possível de agradecer. Sem vocês tudo o resto não existiria. Aos meus pais pelo exemplo de integridade, esforço, dedicação, amizade e amor, obrigada. À minha irmã como eterna companheira e amiga obrigada por continuares a crescer comigo. Só posso esperar que se sintam tão orgulhosos de mim como eu me sinto em ser vossa filha e irmã, e continuar a contar com o vosso apoio para as próximas etapas da minha vida.
ix
The Corpus Callosum is the greatest of the three telencephalic commissures and is exclusive of placental mammals. This white matter structure mediates interhemispheric transfer of information, symmetrically connecting the neocortex of the two cerebral hemispheres. The Corpus Callosum is divided into four main segments (rostrum, genu, body and splenium) in a rostral-caudal direction, with an extra division (isthmus) also considered in human medical literature. Abnormalities of the Corpus Callosum may be classified according to their nature in acquired lesions, including the ones induced by lysosomal storage diseases, and congenital malformations.
Corpus Callosal Abnormalities is a broad designation for congenital malformations of the corpus callosum. These malformations are an important object of study in human medicine, with a wide reported spectrum of clinical manifestations and with numerous associated cerebral or extra-cerebral malformations described. The aetiology and pathophysiology are not yet completely understood nor the long-term implications in cognitive, social and behavioural functions. In veterinary medicine, Corpus Callosal Abnormalities are poorly characterized since literature case reports are scarce. According to current knowledge, Staffordshire Bull Terriers and Miniature Schnauzers are predisposed breeds to Corpus Callosal Abnormalities and to adipsic/hypodipsic hypernatremia, the most frequent clinical manifestation in this pathology. Partial agenesis or hypoplasia of the Corpus Callosum seems to affect more frequently the rostral segments of this structure in cats and dogs. The main brain malformations associated with Corpus Callosal Abnormalities in these species are believed to be mild subtypes of lobar Holoprosencephaly.
The purpose of this dissertation was to understand the Corpus Callosum’s role and importance in humans and animals, and to review literature case reports to date about congenital callosal malformations in cats and dogs, with the goal to characterize three clinical cases presented to the Hospital Veterinari Mollins. The three clinical cases correspond to two dogs and one cat: two of them took place in 2019 and the other in 2013, being included in this dissertation considering the current knowledge about this theme. The animals in question had different clinical presentations and were submitted to a complete magnetic resonance imaging exam with a 0.25 T magnetic field strength equipment. The callosal morphology was abnormal in all three animals. The abnormalities of the Corpus Callosum were classified as acquired in one clinical case, due to presence of a lysosomal storage disease, and as congenital in the other two. According to the magnetic resonance features, the congenital callosal abnormalities corresponded with the current literature in one case only. The main limitations in this
x
this subject to properly characterize each clinical case. Future studies and investigations are necessary to expand the understanding and knowledge about Corpus Callosal Abnormalities in cats and dogs.
Key words: corpus callosum; corpus callosal abnormalities; congenital malformations; cat; dog.
xi
O Corpo Caloso é a maior das três comissuras telencefálicas e é exclusivo dos mamíferos placentários. Esta estrutura de substância branca regula a transmissão de informação inter-hemisférica, comunicando simetricamente o neocórtex dos dois hemisférios cerebrais. O Corpo Caloso é dividido em quatro segmentos principais (rostro, joelho, corpo e esplénio) em direção rostro-caudal, sendo também considerada uma divisão extra (istmo) na literatura médica humana. As alterações do Corpo Caloso podem ser classificadas segundo a natureza das mesmas em lesões adquiridas, incluindo as resultantes de doenças de armazenamento lisossomal, e em malformações congénitas.
Anomalias do Corpo Caloso é uma designação abrangente para as malformações congénitas do corpo caloso. Estas malformações são um objeto importante de estudo em medicina humana, existindo um vasto espetro de manifestações clínicas relatadas e numerosas malformações cerebrais e extra-cerebrais associadas descritas. A etiologia e fisiopatologia ainda não estão totalmente esclarecidas nem as implicações a longo prazo nas funções cognitiva, social e comportamental. Em medicina veterinária, as Anomalias do Corpo Caloso estão pobremente caraterizadas, dada a escassez de relatos de caso na literatura. De acordo com o conhecimento atual, as raças Staffordshire Bull Terrier e Miniature Schnauzer são predispostas a Anomalias do Corpo Caloso e a hipernatremia adípsica/hipodípsica, a manifestação clínica mais frequente nesta patologia. A agenesia parcial ou hipoplasia do Corpo Caloso parece afetar mais frequentemente os segmentos rostrais desta estrutura em cães e gatos. Pensa-se que subtipos ligeiros de Holoprosencefalia lobar sejam as principais malformações associadas a Anomalias do Corpo Caloso nestas espécies.
O objetivo desta dissertação consistiu na compreensão do papel e importância do Corpo Caloso em humanos e animais, e no estudo de relatos de casos da literatura até à data sobre malformações calosas congénitas em cães e gatos, com a finalidade de caraterizar três casos clínicos apresentados ao Hospital Veterinari Mollins. Os três casos clínicos são relativos a dois cães e um gato, tendo dois deles ocorrido em 2019 e o restante em 2013, sendo introduzido nesta dissertação considerando o atual conhecimento sobre este tema. Os animais em questão apresentavam diferentes quadros clínicos e foram submetidos a um exame imagiológico de ressonância magnética completo, através de um equipamento com um campo magnético com uma intensidade de 0.25 T. A morfologia do Corpo Caloso verificou-se anormal nos três animais. As alterações do corpo caloso foram classificadas como adquiridas num caso clínico, devido à presença de uma doença de armazenamento lisossomal, e como congénitas nos outros dois. De acordo com as caraterísticas imagiológicas
xii
as restrições do equipamento de baixo campo magnético na caracterização detalhada dos achados imagiológicos, e a escassa informação disponível sobre este tema para caracterizar adequadamente cada caso clínico. São necessários estudos e investigações futuras de forma a expandir a compreensão e conhecimentos sobre Anomalias do Corpo Caloso em cães e gatos.
Palavras chave: corpo caloso; anomalias de corpo caloso; malformações congénitas; gato; cão.
xiii
Acknowledgements ... vii
Abstract ... ix
Resumo ... xi
Index of Figures... xv
Index of Tables ... xvii
List of Abbreviations ... xix
1. Introduction...1
2. State of the Art ...3
2.1. The cerebral hemispheres ...3
2.1.1. The cerebral cortex ...3
2.1.2. The cerebral commissures ...6
2.2. The Corpus Callosum ...7
2.2.1. Embryology ...7
2.2.2. Structure ...11
2.2.3. Function ...13
2.2.3.1. The Corpus Callosum and brain lateralization ...14
2.2.4. Neuroimaging ...15
2.3. Corpus Callosal Abnormalities ...18
2.3.1. Pathophysiology ...20
2.3.2. Holoprosencephaly and Lysosomal storage diseases ...21
2.3.2.1. Holoprosencephaly ...21
2.3.2.2. Lysosomal storage diseases ...22
2.3.2.2.1. Mucopolysaccharidosis type I ...23
2.3.2.2.2. Neuronal Ceroid Lipofuscinosis ...24
2.3.2.2.3. Sphingolipidoses ...24
2.3.3. Classification ...26
xiv
2.3.6. Management and Prognosis ...37
2.3.6.1. Prognosis of a Lysosomal storage disease ...38
3. Study Objectives ...41
4.Case Presentation ...43
4.1. Materials and Methods ...43
4.2. Clinical Cases ...44 4.2.1. Clinical case nº1 ...44 4.2.2. Clinical case nº2 ...48 4.2.3. Clinical case nº3 ...52 5. Discussion ...55 6. Conclusion ...69 7. References ...71
xv
Figure 1 – Schematic representation of the four main cerebral lobes according to Evans & De Lahunta (2010) (right hemisphere) and Uemura (2015) (left hemisphere): frontal lobe (pink), temporal lobe (blue), parietal lobe (gray) and occipital lobe (green) [Adapted from Uemura (2015)]. ...4 Figure 2 – (A) Schematic representations of the development of the three main telencephalic commissures; (B) The CC relationship with other midline structures [Adapted from de DeLahunta et al. (2015)]. ...8 Figure 3 – Diagram of some domestic animal species’ CC and the human CC. The posterior third (splenium) is represented in all species and the posterior fifth in the human CC, delimiting the isthmus. The image is not to scale [(Adapted from Olivares et al. (2000)]. ...12 Figure 4 – Midsagittal T2-weighted image of an adult dog’s brain. The main segments of the CC are highlighted (red arrows): R – rostrum; G – genu; B – body; S – splenium. The RC (RC) and fornix (F) are also indicated (black arrows) (Adapted from http://vanat.cvm.umn.edu/mriHeadAtlas/sagHeadAtlas.html). ...16 Figure 5 – Difference between two midsagittal T2-weighted images of two healthy Boxer dogs with different magnetic field strengths: 1.5T (A) and 0.25T (B) (Adapted from Mai, 2018)...16 Figure 6 - Lateral aspects of the six segmented tracts of the canine CC in the left hemisphere represented through DTT superimposed on a parasagittal T2-weighted image. The callosal projections are Orbital Frontal (red), Anterior Frontal (orange), Superior Frontal (yellow), Parietal (green), Occipital (purple) and Temporal (blue) [Adapted from Pierce et al. (2014)]. ...17 Figure 7 – Midsagittal T2-weighted images of three different case reports of CCA (A) in a cat and in (B and C) two dogs, with different magnetic field strengths: 0.4T (A and B) and 3T (C). The CC’s rostrum and genu are affected in all three cases. (C) the splenium (white arrow) and the fornix (black arrow) are highlighted [Adapted from Shimbo et al. (2018) (A), Shimokawa Miyama et al. (2009) (B) and P. J. Johnson et al. (2019) (C)]. ...34 Figure 8 - Tractography three-dimensional image of the CC (blue) in a dorsoventral view in a normal control dog (A) and a CCA dog (B). The CC fibers only cross the midline at the level of the splenium and the Probst bundles (red arrows) are longitudinal callosal fascicles [Adapted from P. J. Johnson et al. (2019)]. ...35 Figure 9 – Midsagittal and parasagittal T2-weighted images of the brain. (A) The CC is partially absent with a visible hypoplastic genu, while the rostrum, body and splenium cannot be observed. (B) The fornix is barely visible. ...46
xvi
asymmetric with ventriculomegaly of the right lateral ventricle. The cingulate gyri have an abnormal morphology and project to the ventricular walls. The subcortical WM is abnormally thin and cannot be clearly distinguished form GM. ...47 Figure 11 – Midsagittal T2-weighted image of the brain. The CC is partially absent with a visible hypoplastic body and splenium, while the rostrum and genu cannot be observed. ....49 Figure 12 – Midsagittal T2-weighted image of the brain. The CC is partially absent with a visible hypoplastic rostrum and genu, while the body and splenium cannot be observed. ....53 Figure 13 – Transverse T2-weighted images at the level of the interthalamic adhesion (A) and mesencephalic aqueduct (B). Diffuse subcortical WM hyperintensity is observed in both images...53
xvii
Table 1 – Signalment of seven case reports of cats and dogs with CCA and their respective literature citation. ...19 Table 2 – Clinical presentations of seven case reports of cats and dogs with CCA (signalment and citations in Table 1). ...28 Table 3 – Brain abnormalities described in seven case reports of cats and dogs with CCA (signalment and citations in Table 1). ...33 Table 4 – Serum Na, Cl and K levels (mmol/l) during initial hospitalization (Day 1, 4 and 5) and re-admission (Day 7) [reference values: Na (142–152 mmol/l), Cl (110–124 mmol/l) and K (3.9–5.1 mmol/l)]. ...49 Table 5 – Serum Na, Cl and K levels (mmol/l) after spironolactone therapy initiation [reference values: Na (142–152 mmol/l), Cl (110–124 mmol/l) and K (3.9–5.1 mmol/l)]. ...51
xix ADC Apparent Diffusion Coefficient
CBC Complete Blood Count CC Corpus Callosum
CCA Corpus Callosal Abnormality(ies) CNS Central Nervous System
CSF Cerebrospinal Fluid DTI Diffusion Tensor Imaging DTT Diffusion Tensor Tractography FA Fractional Anisotropy
FeLV Feline Leukaemia Virus FIV Feline Immunodeficiency Virus
FLAIR Fluid-Attenuated Inversion Recovery GM Gray Matter
GW Glial Wedge
HC Hippocampal Commissure HPE Holoprosencephaly IGG Indusium Griseum Glia LR Lamina Reuniens
LSD Lysosomal Storage Disease(s) LT Lamina Terminalis
MD Mean Diffusivity
MRI Magnetic Resonance Imaging MZG Midline Zipper Glia
PNS Peripheral Nervous System RC Rostral Commissure
1
1. Introduction
The Corpus Callosum (CC) is the most prominent telencephalic commissure in placental mammals and is responsible for connecting neocortical homotopic areas of the two cerebral hemispheres, exchanging information between the right and left sides of the brain (Aboitiz & Montiel, 2003; Raybaud, 2010; Tole, Gutin, Bhatnagar, Remedios, & Hébert, 2006). Once thought of as the ‘seat of the soul’ (Di Leva, Tschabitscher, & Rodriguez y Baena, 2007), the CC is now known to be essential for functional integration of cognitive and sensory interhemispheric information, although the manner through which this process occurs is not completely elucidated (Bénézit et al., 2015; van der Knaap & van der Ham, 2011). In human medicine the CC is classically divided into five segments (rostrum, genu, body, isthmus and splenium) in a rostro-caudal direction (Fabri, Pierpaoli, Barbaresi, & Polonara, 2014; Vannucci, Barron, & Vannucci, 2017), whereas in veterinary medicine the genu, body and splenium are considered the main divisions (DeLahunta, Glass, & Kent, 2015), but the rostrum, although much smaller than in humans (Rakic & Yakovlev, 1968), is also considered as a segment by several authors thorough the literature.
Abnormalities of the CC can be categorized into acquired or congenital (Krupa & Bekiesinska-Figatowska, 2013). Corpus Callosal Abnormalities (CCA) are one of the most frequent congenital cerebral anomalies in humans, usually associated with other cerebral or extra-cerebral malformations, but also existing as isolated entities, however this case is less common (Palmer & Mowat, 2014; Romaniello et al., 2016; Schell-Apacik et al., 2008). Extensive research about CCA has been conducted over the years, but because of the CC’s complex multisteped embryology in addition to the strong relationship with the other two interhemispheric great commissures, the potential factors and mechanisms that can interfere with the CC’s neurodevelopment are immense (Dobyns, 1996; Raybaud, 2010; Sztriha, 2005). The Magnetic Resonance Imaging (MRI) is the preferred imaging method to evaluate the CC, especially in midsagittal T2-weighted images (Battal, Kocaoglu, Akgun, Bulakbasi, & Tayfun, 2010; Hasegawa et al., 2013).
CCA in cats and dogs have been mostly reported as secondary features in a group of cerebral malformations, some of them included in the spectrum of lobar Holoprosencephaly (HPE) (Shimbo, Tagawa, Yanagawa, & Miyahara, 2018; Shimokawa Miyama et al., 2009; Sullivan, Harmon, Purinton, Greene, & Glerum, 2003). Only recently, a retrospective MRI study in fifteen dogs was performed, in which the authors highlighted CCA as the main cerebral abnormality and described the associated clinical presentations and concomitant brain malformations associated with this condition (Gonçalves et al., 2014). Since this study, interest in reporting
2
CCA in these species as well as in understanding the inherent pathophysiology and its consequences in interhemispheric connectivity through new imaging techniques has increased (P. J. Johnson, Barry, Luh, & Davies, 2019; Wang-Leandro, Dennler, & Beckmann, 2018). Nonetheless, CCA in cats and dogs have only been sporadically described and reported in the veterinary medical literature, hence why the clinical implications and imaging features are still poorly characterized. According to current knowledge, Staffordshire Bull Terriers and Miniature Schnauzers seem to be predisposed breeds to this malformation and to the most frequently associated clinical presentation, hypodipsic/adipsic hypernatremia (Gonçalves et al., 2014; Shimokawa Miyama et al., 2009; Wang-Leandro et al., 2018). In addition, when partial agenesis occurs, the rostral portion of the CC seems to be the most frequently affected area in cats and dogs (Gonçalves et al., 2014; P. J. Johnson et al., 2019; Shimbo et al., 2018; Wang-Leandro et al., 2018) in opposition to humans, where the caudal portion of this commissure is usually affected (Raybaud, 2010).
In this dissertation a complete state of the art review about the CC’s embryology, evolutionary importance, structure and function, as well as abnormalities of this structure in cats and dogs, with special emphasis in congenital malformations, was made. The goal was to properly contextualize and analyse three distinct clinical cases presented to the Hospital Veterinari Mollins in which the animals were submitted to MRI exams and findings of an abnormal CC’s morphology were described. Further, with this work we hope to raise interest in future research in CCA in cats and dogs and to increase its recognition among veterinary clinicians.
3
2. State of the Art
2.1. The cerebral hemispheres
The Central Nervous System (CNS), comprised by the brain and the spinal cord, and the Peripheral Nervous System (PNS), the set of all the cranial, spinal and named nerves of the body, are considered to be the two main divisions of the nervous system (DeLahunta et al., 2015) although some authors consider the Autonomic Nervous System (ANS) as the third main component (Thomson & Hahn, 2012; Uemura, 2015). The coordination and integration of the different parts of the nervous system are the brain’s responsibility, the control organ of the body (König & Liebich, 2007).
The prosencephalon (forebrain), the most rostral primitive brain vesicle, gives rise to the telencephalon (rostrally) and diencephalon (caudally) (McGeady et al., 2017). The telencephalon later expands into two telencephalic vesicles that grow dorsally and caudally above the diencephalon and mesencephalon, originating two cerebral hemispheres that comprise the cerebrum and are connected to each other by the CC (DeLahunta et al., 2015; McGeady et al., 2017). The longitudinal cerebral fissure, in the dorsal midline of the telencephalon, separates the two hemispheres and contains the falx cerebri, a fold of dura mater (Skerritt, 2018). Although structurally symmetrical, each cerebral hemisphere contributes in a unique way to the brain’s global function (Uemura, 2015).
2.1.1. The cerebral cortex
Excluding the rodents, lagomorphs, birds and most marsupials, which have a smooth hemisphere surface (lissencephalic), most animal species of veterinary interest have a convoluted hemisphere surface (gyrencephalic), although carnivores and primates remain lissencephalic throughout gestation for a considerable period (Thomson & Hahn, 2012). The cerebral convolutions are the outward folds (gyri) between the inward folds (sulci), and each gyrus is made of gray matter (GM) superficially (cerebral cortex) and white matter (WM) in its core (corona radiata) containing the axons of neurons that connect to and from the cortex (Butler & Hodos, 2005; DeLahunta et al., 2015). This anatomical configuration is due to the accelerated growth of the cortex compared to the underlying WM, allowing a larger cortex’s surface (Uemura, 2015). While the cerebral cortex is the superficial GM that covers the cerebral hemispheres, the septal and basal nuclei and the hippocampus are GM structures within the hemispheres (Thomson & Hahn, 2012).
4
Despite correspondence differences between species exist, one can divide the cerebral cortex into four main lobes according to the cranium’s bones that topographically relate to the corresponding areas of the brain into frontal, parietal, occipital and temporal lobes, although this division is not exact (DeLahunta et al., 2015; McGeady et al., 2017). The anatomical limits of each lobe also differ between different authors. For example, the inclusion of the postcruciate gyrus in the frontal lobe is defended by Uemura (2015) but according to Evans & De Lahunta (2010) this gyrus is part of the parietal lobe. Nevertheless, conciliating both authors (first Uemura (2015) and second Evans & DeLahunta (2010)), the four main neocortical lobes in each hemisphere might be grossly identified as: frontal lobe – includes all gyri rostral to the postcruciate gyrus (included) or cruciate sulcus (postcruciate gyrus not included); temporal lobe – includes the sylvian and ectosylvian gyrus or the ventrolateral aspects of the hemispheres; parietal lobe – includes the mid portion of the suprasylvian gyrus and the rostral endomarginal and marginal gyri or caudal to the cruciate sulcus until the caudal third of the hemisphere, dorsal to the sylvian gyrus; occipital lobe – dorsocaudal to the temporal lobe and caudal to the parietal lobe or caudal third of the hemispheres (Figure 1).The piriform lobe, a ventral bulge medial to the temporal lobe, belongs to a different phylogenetically portion of the brain (paleopallium), therefore it is considered as a distinct entity from the other neocortical lobes.
Another classification or division system of the cerebral cortex is based on its phylogenetical origin, in which there are three major divisions: the paleopallium (paleocortex), the archipallium (archicortex) and neopallium (neocortex), with ‘pallium’ and ‘cortex’ as synonyms although pallium is more comprehensive (includes the ependyma and pia mater). In mammals, the Figure 1 – Schematic representation of the four main cerebral lobes according to Evans & De Lahunta
(2010) (right hemisphere) and Uemura (2015) (left hemisphere): frontal lobe (pink), temporal lobe (blue), parietal lobe (gray) and occipital lobe (green) [Adapted from Uemura (2015)].
5
paleocortex and archicortex (the oldest regions) give rise to the olfactory region of the brain and the hippocampus (an internal gyrus), respectively (DeLahunta et al., 2015; Skerritt, 2018). The neocortex is separated from the other phylogenetical portions by the splenial sulcus (archicortex) medially, and rhinal sulcus (paleocortex) ventrally, comprising the largest portion of the cerebrum (Thomson & Hahn, 2012). The neocortex has two types of areas regarding their function: the projection areas (primary cortical areas) and the association areas. The projection areas are responsible for receiving sensory information (primary somesthetic area, primary visual area and primary auditory area) from the brainstem and spinal cord, providing awareness of sensation, and they also initiate motor activity (primary motor area). The association areas receive information from the above, recognize the sensations, interpret it, compare it with previous experiences and select the most adequate response, while predicting its consequences (Butler & Hodos, 2005; Skerritt, 2018).
The primary motor area includes the precruciate, postcruciate and rostral suprasylvian gyri, the last two gyri being also part of the primary somesthetic area in addition to the rostral part of the ectosylvian gyri. In the occipital lobe, the caudal part of the marginal gyrus and the surrounding gyri form the primary visual area, while the primary auditory area is situated in the middle ectosylvian gyrus of the temporal lobe (Uemura, 2015). There are three types of association areas: cognitive, interpretative and the frontal association areas. In cats and dogs, cognitive association areas are in the parietal and occipital lobes, while the interpretative association area resides in the temporal lobe, with a presumable role in learning and memory. The frontal association area or premotor frontal association area isn’t well developed in these species, but contributes to alertness, intelligence and individual temperament (Skerritt, 2018). The visual association area extends from the occipital lobe to the temporal and parietal lobe, while somesthetic association areas reside in the upper part of the temporal lobe and in more posterior parts of the parietal lobe, and the auditory association area is in the temporal lobe (Butler & Hodos, 2005). Despite the percentage of association areas the cerebral cortex of cats and dogs is only of 20% compared to human’s 85%, it is enough to allow the above described processes to occur in an individual manner for each individual, enabling them with a characteristic personality (Skerritt, 2018).
Histologically, the neocortex of mammals is essentially constituted by six layers of neuronal cell bodies in its cortical plate (Richards, 2002), which differs from the archicortex and paleocortex that have fewer layers (DeLahunta et al., 2015). The molecular layer (layer I) is the most superficial of all and is mainly composed by dendritic ramifications of deeper layers’ neurons and afferent axons. The external granular layer (layer II) has small neurons with short axons, the granule cells, while in the external pyramidal layer (layer III) the pyramidal cells
6
spread apical dendrites to the cortex’s external surface. Both cell types project axons to deeper layers and different ipsilateral cortical regions or to contralateral cortical regions through the CC. The internal granular layer (IV) is formed by granule cells, the internal pyramidal layer (V) by pyramidal cells and the multiform layer (VI) by fusiform cells (Butler & Hodos, 2005). The correct neurodevelopment of the six layers is a crucial point for the CC’s development, since the majority of callosal fibres originates from neurons within these layers (Richards, Planchez, & Ren, 2004).
2.1.2. The cerebral commissures
There are three types of axons from telencephalic neurons that form the WM in the cerebral hemispheres according to their destination: association, projection and commissural axons. The first type are interneurons that connect cortical areas of the same hemisphere, and they can be short (e.g.: arcuate fibres) or long (e.g.: superior longitudinal fasciculus). The second type of axons connect the cerebral hemisphere to further elements of the CNS and are introduced into the brainstem via the internal capsule. Finally, the commissural axons are the axons that communicate one cerebral hemisphere with the other, across the midline. In the corona radiata, the three types of axons blend together and form the gyri’s WM. (DeLahunta et al., 2015; Skerritt, 2018).
Commissural pathways in the telencephalon are cortico-cortical bundles of WM that symmetrically connect the cortices of both hemispheres. While decussations associate different structures, commissures associate homologous ones in the CNS and are required for bilateral integration and good body coordination (Raybaud, 2010). In able to connect the two hemispheres, three great interhemispheric commissures arise, one for each phylogenetic division of the cerebrum: the Rostral Commissure (RC) (paleocortex), the Hippocampal Commissure (HC) (archicortex) and the CC (neocortex) (Raybaud, 2010; Tole et al., 2006). The first two are common to all mammals, including marsupials and monotremes, while the CC is only present in some marsupials and in all placental mammals (Butler & Hodos, 2005), hence why some authors acknowledge the CC as a characteristic of placental mammals as the placenta itself (Aboitiz & Montiel, 2003).
The RC develops in the lamina terminalis (LT), dorsal to the optic chiasma, and crosses the midline rostral to the hypothalamus, interconnecting the ventrolateral parts of the telencephalon: the bilateral rhinencephalic structures such as the olfactory lobes, piriform lobes and the amygdaloid bodies (Butler & Hodos, 2005; DeLahunta et al., 2015). In animals like the marsupials and monotremes, apart from the paleocortical ones, fibres that arise from the neocortex cross through this commissure (Aboitiz & Montiel, 2003), being considered the
7
brain’s most evolutionarily conserved commissure (Lodato & Arlotta, 2015). The HC is a small triangular structure that connects the hippocampus of both hemispheres through commissural axons that cross the midline between the fornical crura (part of the fornix) dorsally to the caudal aspect of the diencephalon (DeLahunta et al., 2015; Raybaud, 2010).
2.2. The Corpus Callosum
The CC (L. corpus, body; callosum, hard) is the most prominent commissure in the placental mammals’ brain and is responsible for connecting neocortical regions of each hemisphere (Mihrshahi, 2006; Uemura, 2015), enabling functional integration of cognitive and sensory information between them (van der Knaap & van der Ham, 2011). The fact that CC’s ancestral structures do not exist in nonplacental mammals, suggests that the CC results from an abrupt and complex evolutionary origin in placental mammals (Aboitiz & Montiel, 2003) as a phylogenetically new feature (Raybaud, 2010). Each neocortical area projects callosal axons across the CC to the contralateral hemisphere in order to connect mostly homotopic (homologous) areas, but also heterotopic (heterologous) ones (Aboitiz & Montiel, 2003; Innocenti, 1986). The neocortical callosal axons are mostly projected from cortical layers II, III and V (Richards, 2002; Richards et al., 2004).
2.2.1. Embryology
In 1968 Rakic & Yakovlev carried out a study in order to clarify the sequence of events that lead to the development of the cerebral commissures, including the CC and the structures related to it (e.g.: the cavum septi and septum pellucidum), in several whole human embryos, human cerebra and in additional cerebra of animals (e.g.: domestic animals like cats, dogs and mice) for comparison purposes. The work from these authors is still considered a fundamental and cardinal study, being one of the most frequently quoted articles on the subject of cerebral commissures development (Raybaud, 2010), so a brief summary of their work will be described next.
The three cerebral commissures originate from the rostral wall of the telencephalon (‘telencephalon impar or medium’) in very early embryonic stages, through complex, coordinated and very rapid processes. The rostral wall begins as a homogeneous cell-mass that rapidly evolves into different portions: the dorsal and densely cellular lamina reuniens of His (LR) and the ventral and thin lamina terminalis (LT).The area praecomimissuralis arises from the ventral part of the LR, that later becomes the septal area with its nuclei, and through which fibre bundles from the ganglionic eminences cross the midsagittal plane, growing into a thick bundle that later becomes the definitive RC. Complex events occur in the dorsal part of
8
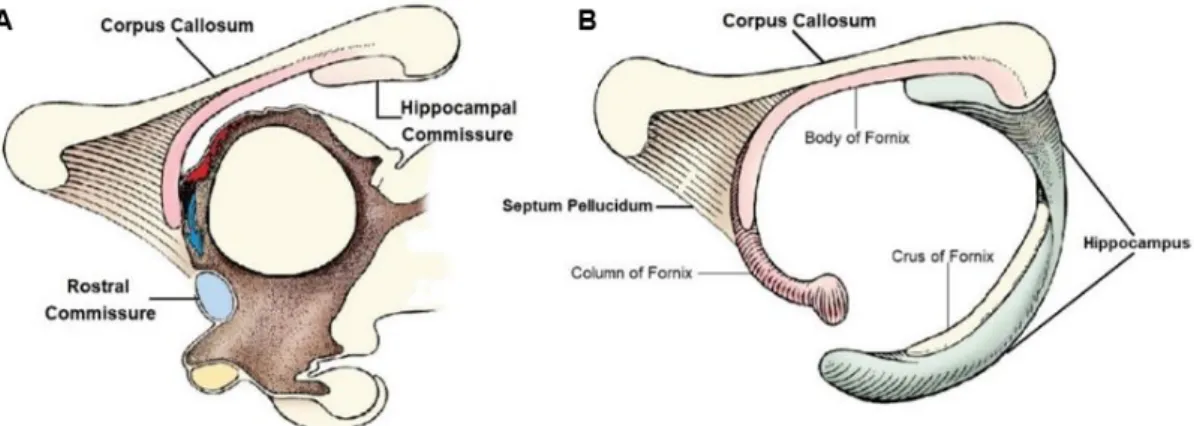
the LR that culminate with the development of several different structures, beginning with the emergence of axons of the two primordia of the fornix from the area praecomissuralis. Afterwards, axons cross between the two primordia of the fornix and originate the psalterium hippocampi and the HC. Meanwhile, a median groove (sulcus medianus telencephali medii) arises as the result of the infolding of the LR, immediately below the interhemispheric fissure, that ventrally progresses. Over time, the margins of the groove approximate, juxtapose and fuse into a massa commissuralis, the foundation through which the callosal fibres from the neocortex penetrate and grow to form the definitive CC. The septum pellucidum arises from the lateral margins of the sulcus medianus telencephali medii below the CC, connecting it to the fornix (Figure 2). In carnivores and rodents, the cavum septi remains open due to the CC’s rudimentary genuflexion and the CC’s rostrum is poorly developed compared to that of humans. During embryonic development, the form and position assumed by the CC is like that of the adult brain, posteriorly changing mostly in its thickness and rostro-caudal extent, which reflects the growth and surface changes of the cerebral hemispheres. The CC’s development is considered to be the same in all placental mammals (Rakic & Yakovlev, 1968).
The emergence of the nervous system’s midline, regulated by genetic patterning, is a vital process for the CC’s embryonic development. Without midline fusion, there is no substrate for the axons to thrive and cross to the other hemisphere (Richards et al., 2004). In 1982, Silver, Lorenz, and Wahlsten were the first to report the role of midline glial structures in this process (Nishikimi, Oishi, & Nakajima, 2013), stating that the margin apposition and fusion between the medial hemispheres’ walls described by Rakic & Yakovlev (1968) was only a prerequisite for the normal CC’s formation, but not enough for it to happen. According to the authors, a midline primitive ‘glial’ population from the ependymal zones migrated medially through the
Figure 2 – (A) Schematic representations of the development of the three main telencephalic
commissures; (B) The CC relationship with other midline structures [Adapted from de DeLahunta et al. (2015)].
9
fused walls to form a bridge or sling like structure (‘glial sling’), before the crossing of the first callosal axons just above it, only to later disappear after birth. It was also proposed that this primitive cellular had a positive influence in guiding the callosal axons through the midline (Silver et al., 1982) and could even rescue callosal axons from aberrant paired neuromas in chirurgical acallosal mice when presented in a scaffold (Silver & Ogawa, 1983). It was later clarified by Shu, Li, Keller & Richards (2003) that these cells were a migratory population of immature neurons from the subventricular zone, with no typical glial markers (e.g.: GFAP – glial gibrillary acidic protein), emerging only after the first callosal axons crossed the midline, therefore suggesting they could cross it without the sling. These authors redefined the sling as a neuronal migratory population whose function might include the guidance of callosal axons through the midline, that continued to migrate to populate other cerebral regions (Shu, Li, Keller, & Richards, 2003).
Glial cell structures have been described to participate in the compartmentalization of brain regions and in the organization of axon systems, acting as guides or barriers (Silver, Edwards, & Levitt, 1993). The guidance of axons through the midline occurs via signals at the corticoseptal boundary (Rash & Richards, 2001). Growth cones, the axons’ edge, have cell surface receptors that interpret cues in the environment as attractive/positive or repulsive/negative, in order to navigate over trajectories, creating wiring patterns (Fothergill et al., 2014; Kaprielian, Imondi, & Runko, 2000).These guidance cues are presented by midline specialized cells that function as intermediate targets (Kaprielian et al., 2000).There are three types of midline glial populations in the corticoseptal boundary, that probably act as key roles in the guidance of commissural axons: the glial wedge (GW), the indusium griseum glia (IGG) and the midline zipper glia (MZG) (Shu, Puche, & Richards, 2003; Shu & Richards, 2001; Silver et al., 1993; Tole et al., 2006).
All three midline glial populations mentioned above express the GFAP marker through immunohistochemistry and have different morphologies and relative locations in the midline (Shu, Puche, et al., 2003). The midline zipper glia (“massa commissuralis” of Rakic and Yakovlev), a specialized population of astroglia, helps to close (‘zipper’) the midline before its fusion. This cellular population also directs and promotes axon growth and helps remodel the basal lamina during the fusion process (Raybaud, 2010; Silver et al., 1993). The GW population has its cell bodies in the ventricular zone of the lateral ventricles and forms a wedge shape structure in their dorsomedial aspect, promoting an abrupt turning of the callosal axons towards the midline (Shu & Richards, 2001). The IGG originates in the indusium griseum (Shu & Richards, 2001), a part of the limbic system that consists of two thin GM bands that lie directly above the CC (Di leva, Fathalla, Cusimano, & Tschabitscher, 2015). Both of these glial
10
populations express the slit-2 molecule, with a repellent effect on neocortical axons by interacting with their slit-2 robo receptors, thus guiding callosal axons towards and across the midline through surrounding repulsion (Shu & Richards, 2001).
The CC extends across the midline between the indusium griseum and glial wedge (Tole et al., 2006). Molecules expressed by midline cell populations (diffusible or attached to cellular surfaces or extracellular matrix) guide the axons through chemoattracting or repulsion of the growth cones to their innervation target. Some molecules are responsible for fasciculation and defasciculation (Raybaud, 2010; Richards, 2002). Fasciculation of growing axons to pioneering axons is another important mechanism of axon guidance in large tract formation (Richards, 2002). Pioneering axons are the first to form a specific pathway/trajectory and they facilitate the growth of later arriving axons (Rash & Richards, 2001), that use the first ones to find the correct path, while using guidance molecules for other purposes such as innervation of target points along the main trajectory (Richards, 2002; Richards et al., 2004).
The pioneering callosal axons originate from neurons within the cingulate cortex and are the first ones to cross the midline to the contralateral hemisphere, providing a means of growth and guidance for the neocortical axons within the new fibre tract (Koester & O’Leary, 1994). The cingulate gyri are telencephalic components of the limbic system in the hemispheres’ medial surface, and their corona radiata is called the cingulum, a long association tract that courses from the parahippocampal gyrus to the frontal lobe (Butler & Hodos, 2005). Nevertheless, some authors consider that the pioneering axons in the Koester & O’Leary study belonged to the perforating cingulate fibres that perpendicularly intersect the CC to the hippocampus (Ozaki & Wahlsten, 1998). They proposed that the first cortical neurons to send axons through the CC came from the dorsolateral portion of the frontal cortex (Ozaki & Wahlsten, 1992, 1998) but any callosal axon from every cortical region had the ability to cross the midline first (Ozaki & Wahlsten, 1993). In order to clarify this question Rash & Richards (2001) used a different technique in a similar study, and they concluded that the hypothesis proposed by Koester & O’Leary in 1994 was right: axons from the cingulate cortex are the callosal pioneering axons and callosal neocortical axons cross the midline within the bundle of cingulate fibres. Furthermore, some cingulate axons project laterally to the rostrolateral neocortex and guide neocortical axons towards the midline (Rash & Richards, 2001; Richards, 2002).
According to some authors, topographic information about rostro-caudal and medio-lateral organization exists in callosal fibres since early embryonic stages, with differences in the rostro-caudal projection contribution of callosal axons, from different cortical areas in the CC formation. This implies a rostral to caudal unidirectional CC formation gradient, from the frontal
11
to the occipital lobe, and that the genu formed first than the splenium (Ozaki & Wahlsten, 1992). This theory was supported by different studies from the same authors who concluded that CC’s growth followed the proliferation and maturation of neocortical cells, which occurs in a rostro-caudal and lateral-medial directions (Ozaki & Wahlsten, 1993, 1998). This proposal was against Rakic & Yakovlev (1968) who believed that the CC development was bidirectional, first caudal then rostral, reflecting the growth of the cerebral hemispheres, being the rostrum the last segment to form. Later, other authors supported the previous theory, stating that the CC’s development was indeed bidirectional. Initially, they affirmed that the rostral body was the first segment to form, followed by the genu, splenium and lastly the rostrum (Kier & Truwit, 1996). However, they later discovered that the lamina rostralis (a segment of the fetal rostrum) was present before the genu and splenium, rostrally anchoring the developing CC while the fornix anchored it caudally (Kier & Truwit, 1997).
A hypothesis that reconciles these two proposals affirms that the CC’s development may occur through different mechanisms in its rostral and caudal regions. The cingulate pioneering axons might be required for axonal crossing in the rostral and middle regions but in more caudal regions (such as part of the body and splenium) the HC might be enough to allow axonal crossing, without the need of cingulate pioneers (Richards et al., 2004). A correlation between the caudal CC with the hippocampal commissure exists, due to their proximity during embryonic development (Livy & Wahlsten, 1997). Thus, the callosal regions would fuse and grow through fasciculation in a rostro-caudal manner forming the complete CC. This would explain the reason why many syndromes that course with callosal abnormalities affect differently the CC’s rostral and caudal components, in human medicine (Raybaud, 2010; Richards et al., 2004). The complex embryonic development of the CC explains the wide heterogeneity of possible causes and mechanisms that can induce its malformation (Dobyns, 1996).
2.2.2. Structure
The CC is constituted by axons (myelinated and unmyelinated) but also by glial cells and blood vessels (Al Qattan et al., 2019). Several methods have been proposed to define regions and subregions within the CC, based on geometric criteria (Fabri et al., 2014; Hofer & Frahm, 2006; Witelson, 1989). The main CC’s segments considered for domestic animals are the genu (rounded rostral end), body (middle), and a splenium (rounded caudal end) (DeLahunta et al., 2015; Uemura, 2015) but a rostrum is also considered by many authors, despite its smaller development comparing to humans (Jacqmot et al., 2013; Rakic & Yakovlev, 1968). The classic segmentation followed in humans includes the rostrum/lamina rostralis, which connects
12
the genu with the LT (Vannucci et al., 2017), and a narrowing between the body and splenium called the isthmus, where the fornix joins the CC (Raybaud, 2010), but one can add additional divisions like an anterior, central and posterior midbody (Fabri et al., 2014).
The gross anatomy of the CC tends to be conservative between species, with some differences in the relative size of its different parts: small extremities with a big mid-third (mouse and rabbit); slender body with bulbous extremities (dog and human); a balanced distribution (cat, cow and horse) (Olivares, Michalland, & Aboitiz, 2000) (Figure 3). However, the relative size of the posterior third (splenium) is the most variable feature: bigger in frontally looking species (cat, dog and human) and smaller in laterally looking species (cow, horse, mouse and rabbit) (Olivares et al., 2000), which might be related to differences in the visual performance, since visual fibres tend to cross the midline in this area (Matsunami, Kawashima, Ueki, Fujita, & Konishi, 1994; Olivares et al., 2000; Phillips & Hopkins, 2012).
When comparing some mammalian species, the relative size of the CC doesn’t keep up with the increase in brain size, in larger species: the relationship between callosal area and cortical surface is smaller than 1 (0.75) (Olivares et al., 2000) and the relationship between the number of splenium fibres and cortical cells is also smaller than 1 (2/3) across species (Olivares, Montiel, & Aboitiz, 2001). The previous facts, in addition to the conservation of the CC’s fibre composition among species (even with the caliber increase of a small group of fibres in larger brain size species), mean that interhemispheric transmission velocity doesn’t increase with brain size, suggesting an impairment in interhemispheric communication that may contribute to hemispheric isolation (Olivares et al., 2001).
In 2000, a study conducted on 21 dogs brains concluded that there are significative differences in the callosal anatomy of different sex dogs, since the total callosal area and length of male
Figure 3 – Diagram of some domestic animal species’ CC and the human CC. The posterior third
(splenium) is represented in all species and the posterior fifth in the human CC, delimiting the isthmus. The image is not to scale [(Adapted from Olivares et al. (2000)].
13
dogs were larger than the females (Aydınlıoglu et al., 2000). A similar dimorphism was also demonstrated in the cat (Tan & Kutlu, 1993). In the rat, sexual dimorphism has been demonstrated both in size (Fitch, Berrebi, Cowell, Schrott, & Denenberg, 1990), total axon density and myelinated and unmyelinated axon density (Al Qattan et al., 2019). Sexual dimorphism of the human CC is a debated theme about which there is still no consensus among researchers (Al Qattan et al., 2019; Ardekani, Figarsky, & Sidtis, 2013). Nevertheless recent studies with advanced imaging techniques have shown effective sex differences in the CC, including differences in cross-sectional area size, fibre organization and microstructural composition (Ardekani et al., 2013; Liu, Vidarsson, Winter, Tran, & Kassner, 2010). The interest in this subject, in humans and animals, is the possibility that sex differences in this structure may be related to differences in interhemispheric connectivity, therefore, related to behaviour functional differences between sexes (Al Qattan et al., 2019; Ardekani et al., 2013; Aydınlıoglu et al., 2000). This information would be useful in future studies in neurological disorders, in which sex influences the outcome (Liu et al., 2010).
The neocortical areas connected by the CC are topographically represented in its fibre organization in a rostro-caudal and dorsoventral manner, according to their fibre origins and their contralateral projection. This topographical organization varies among species (Aboitiz & Montiel, 2003; Hofer & Frahm, 2006; Pierce et al., 2014; Suárez et al., 2018; Zhou et al., 2013). Differences in the caliber and myelinization degree between callosal axons in different callosal segments were described in humans and primates (Aboitiz & Montiel, 2003; Aboitiz, Scheibel, Fisher, & Zaidel, 1992; Olivares et al., 2001), but these regional differences could not be observed in other mammals (like carnivores), at least in the CC’s caudal third (Olivares et al., 2001). According to Richards et al. (2004) the CC is mainly constituted by axons that project from neurons in layers II/III and V.
2.2.3. Function
It was once thought that the CC was the ‘seat of the soul’ (Di Leva et al., 2007). A positive correlation of its thickness with intelligence has been suggested in humans (Fabri et al., 2014) although this theory is still up to discussion (Westerhausen et al., 2018). A great part of today’s knowledge about the CC’s functions was achieved from studies in humans and animals, whom partial or total CC surgical resection was employed as a therapy for drug-resistant epilepsy, in order to stop seizure propagation between hemispheres (Bagley et al., 1995; Schaller & Cabrilo, 2016; van der Knaap & van der Ham, 2011). Another source of knowledge were neuropsychological and anatomical studies in humans with congenital abnormalities of the CC and from other types of developmental anomalies, like autism and schizophrenia (Paul et al.,
14
2007). Moreover, investigations designed with the purpose to determine de CC’s role in interhemispheric communication and brain lateralization, were conducted through specific learning skill’s tests, some of them using only split-brain animals (surgically resected CC) and others using split-brain animals and normal control animals (Glickstein, 2009; van der Knaap & van der Ham, 2011). This studies demonstrated that this commissure is crucial for information transference between the two hemispheres in several aspects, such as visual information (Myers, 1956), conditioned reflexes associated with somesthetic stimuli in one side of the body (Glickstein & Berlucchi, 2008), and transfer of intermanual learned somesthetic discrimination (Glickstein & Sperry, 1960; Stamm & Sperry, 1957). Furthermore, it has been suggested that a cerebellar pathway may also have a role in interhemispheric transfer of information, connecting visual areas in one hemisphere with motor areas in the contralateral hemisphere (Glickstein, 2009).
It is important to mention that CC’s transection does not completely mimic congenital agenesis of the CC: callosotomies performed in mature CCs (human adolescents and adults) may result in ‘disconnection syndromes’ in which interhemispheric communication is impaired, with sensory and bimanual motor coordination consequences; conversely, patients with CC’s agenesis or with an early performed callosotomy, may not exhibit any of these clinical sings, revealing some degree of neuronal compensation and plasticity (Palmer & Mowat, 2014). Disconnection syndromes have been extensively described in humans and animals (Catani & Ffytche, 2005; Geschwind, 1965). A recent study using diffusion weighted imaging and Diffusion Tensor Imaging (DTI), demonstrated for the first time, a subcortical extracallosal mechanism that may be responsible for preserving some interhemispheric communication in a human patient who had complete commissurotomy 40 years earlier, an only speculated hypothesis until now. This enhances the important contribution to nowadays knowledge of the brain’s structure-function relationships and connectivity changes throughout lifespan, of long-time follow up of these type of patients (Nomi et al., 2019).
2.2.3.1. The Corpus Callosum and brain lateralization
As mentioned before, in larger brain size species the interhemispheric transmission velocity does not proportionally increases. Not even a small group of bigger caliber fibres compensates the general increment in brain size and interhemispheric distance, which leads to a transmission delay that probably promotes brain lateralization and hemispheric isolation (Aboitiz & Montiel, 2003; Olivares et al., 2000, 2001; Ringo, Doty, Demeter, & Simard, 1994). In spite of the genetic constraints that may define hemispheric lateralization, the CC still plays a major role, or so it is assumed, in hemispheric function specialization (Bénézit et al., 2015). The only certainty about the CC regarding this theme, is that the CC mediates the transfer of
15
information between the two hemispheres. However, the nature of such information and its influence in the other hemisphere is still unknown (van der Knaap & van der Ham, 2011). There are two different well-defined lines of thought about this subject in the current literature, both of them supported by several authors and scientific evidence (Bénézit et al., 2015):
• The inhibitory theory - The CC is a means for one hemisphere to inhibit the other so that the first can dominate a certain brain function. This would mean that the CC favors the development of brain lateralization and hemispheric independency;
• The excitatory theory - The CC activates the un-stimulated hemisphere concerning a certain function, enhancing cerebral processing between the two hemispheres and consequently decreasing brain lateralization.
Both theories contribute to the evolutionary perspective of the CC. The possibility that this structure does not primarily have only an inhibitory or excitatory function is also considered (van der Knaap & van der Ham, 2011). Interhemispheric lateralization is still a topic of interest that has been a research subject for many years, with the CC as the main protagonist. A lot of different theories about the origin of this phenomenon exist, even though they are still uncertain and highly debated (Nowicka & Tacikowski, 2011).
2.2.4. Neuroimaging
In human medicine, ultrasonography and computed tomography can be used to evaluate the CC, but the modality of choice is the MRI (Battal et al., 2010). The basic physics behind MRI consists in the visualization of hydrogen atoms that exist in body tissues, after submitting them through a main magnetic field and emitting radiofrequency pulses, resulting in different T1 and T2 relaxation times, allowing tissue differentiation (Hetch, 2016). As a WM structure, the CC’s characteristics in MRI sequences consist of a high signal intensity in T1-weighted images and a low signal intensity compared to GM in T2 and proton density weighted images (Georgy, Hesselink, & Jerningan, 1993). Easily recognized in midsagittal MRI sequences, the normal CC overlies the fornix and third ventricle, like an eyebrow bold line structure in T2-weighted images (Hasegawa et al., 2013), with its four main segments: the rostrum, genu, body and splenium (Battal et al., 2010) (Figure 4).
16
In a study performed in 17 dogs’ brains, from one to twelve weeks of age, the CC could be clearly distinguished through a midsagittal T2-weigthed MRI by 8 weeks of age, acquiring only an adult appearance at 16 weeks of age (Gross, Garcia-Tapia, Riedesel, Ellinwood, & Jens, 2010). In cats, a similar study has not been made. However, in a DTI study in cats, the CC could be clearly visualized by 35 days postnatally (Takahashi et al., 2011) and was also identified in a 2 month-old clinically normal cat’s MRI (Hasegawa et al., 2013). It is very important to consider that image quality obtained in MRI sequences is greatly influenced by magnetic field strength, which can vary between three categories: low-field (0.2T – 0.4T), medium field (0.5T- 1.0T) and high field (>1.0T). The most popular in veterinary medicine practice are low-field magnetic strength MRI equipment, while high-field equipment predominate in academic veterinary centers, due to the first’s relatively low cost and easier installation/maintenance (W. Mai, 2018) (Figure 5).
DTI is an MRI modality that tracks axonal fibres through water molecules anisotropy diffusion in tissues. Inside axonal fibres, the water movement is delimited by axonal membranes and myelin sheets, therefore the preferential diffusion in WM tracts is parallel to them (Fabri et al., Figure 4 – Midsagittal T2-weighted image of an adult dog’s brain. The main segments of the CC are
highlighted (red arrows): R – rostrum; G – genu; B – body; S – splenium. The RC (RC) and fornix (F) are also indicated (black arrows) (Adapted from
http://vanat.cvm.umn.edu/mriHeadAtlas/sagHeadAtlas.html).
Figure 5 – Difference between two midsagittal T2-weighted images of two healthy Boxer dogs with different magnetic field strengths: 1.5T (A) and 0.25T (B) (Adapted from Mai, 2018).
17
2014). With the appropriate magnetic field gradients, almost any MRI sequence is able to detect water diffusion and with a diffusion tensor, a mathematic matrix, it is possible to quantitatively describe diffusion anisotropy in the three-dimensional space (Bihan & Johansen-Berg, 2012; Jacqmot et al., 2013). Some DTI metrics are the apparent diffusion coefficient (ADC), factional anisotropy (FA), radial diffusivity (RD) and axonal diffusivity. Regarding myelination, FA, RD and ADC are considered important factors (Leong et al., 2015). Fibre tractography is a technique that uses the diffusion ellipsoids (tridimensional representation of water molecules diffusion in a certain diffusion time) from DTI, to virtually delineate WM tracts in the three-dimensional space (Bihan & Johansen-Berg, 2012).
In 2013, a study performed with the intent to visualize and quantitatively describe WM tracts in dogs’ brains in vivo, CC included, allowed the characterization of normal DTI metric parameters, as well as the representation of these WM tracts through tractography in healthy dogs (Anaya García, Hernández Anaya, Marrufo Meléndez, Velázquez Ramírez, & Palacios Aguiar, 2015). Other authors used DTI and tractography techniques to map WM tracts ex vivo in dogs’ and cats’ brains in order to build a brain atlas (Jacqmot et al., 2013, 2017). Pierce et al. (2014) studied the division of the canine CC through Diffusion Tensor Tractography (DTT), based on the axons’ destinations in the cortex’s major functional divisions. This author divided the dog’s CC into 6 major WM tracts (and segments) in the rostral to caudal axis: orbital frontal, anterior frontal, superior frontal, parietal, occipital and temporal. In the authors’ opinion, dividing the CC according to cortical destination of its fibres is more accurate and precise than to transpose the morphologic division criteria of the human CC to other species, which is difficult due to the different size and morphology of its segments (Figure 6).
Figure 6 - Lateral aspects of the six segmented tracts of the canine CC in the left hemisphere
represented through DTT superimposed on a parasagittal T2-weighted image. The callosal projections are Orbital Frontal (red), Anterior Frontal (orange), Superior Frontal (yellow), Parietal (green), Occipital
18
In addition to the characterization of dogs’ and cats’ WM tracts, DTI studies in these species support the future use of this non-invasive in vivo diagnostic technique as a complementary diagnostic method to conventional MRI. DTI can help to describe microstructural characteristics, morphology and connectivity of the brain’s main fibre bundles when affected by intracranial lesions and pathologies (Anaya García et al., 2015; Jacqmot et al., 2017). DTI usage in dogs is also useful to study WM pathologies that occur in humans, since the canine brain is an excellent model (Peterson, Chen, Calabrese, White, & Provenzale, 2016). DTI and tractography have also been used to describe malformations of the CC in dogs (P. J. Johnson et al., 2019; Wang-Leandro et al., 2018), a theme that will next be approached in this dissertation.
2.3. Corpus Callosal Abnormalities
In human medicine, the term CCA is used as a broad designation that includes all types of congenital morphology abnormalities in the CC (Hanna et al., 2011). Despite the numerous employed terms, in human medical literature, to characterize this entity as a congenital malformation (agenesis, dysgenesis, hypogenesis, hypoplasia, dysplasia) (Hanna et al., 2011; Hetts, Sherr, Chao, Gobuty, & Barkovich, 2006; Palmer & Mowat, 2014; Raybaud, 2010; Schell-Apacik et al., 2008), CCA includes them all (Hanna et al., 2011). By definition, cellular and tissular structural or functional abnormalities are defined as congenital malformations when they are present in an animal since birth (McGeady et al., 2017). However, in some medical literature, the expression ‘abnormalities of the CC’ is used in a more comprehensive manner to include congenital and acquired lesions (Krupa & Bekiesinska-Figatowska, 2013). Various articles review the different types of CC acquired lesions that can be evaluated through the MRI, like vascular and traumatic lesions, neoplasia, lipomas, infections, metabolic diseases, among others (Georgy et al., 1993; Krupa & Bekiesinska-Figatowska, 2013; Uchino, Takase, Nomiyama, Egashira, & Kudo, 2006). In this dissertation the concept CCA will be applied to refer to the spectrum of congenital abnormalities of the CC only, i.e. excluding the acquired ones.
CCA are one of the most common cerebral congenital malformations among humans, but they are highly unspecific, given that they can be part of the clinical manifestations of a congenital syndrome (some of them associated with specific chromosomal arrangements and gene mutations) and can occur associated with other brain malformations or as isolated entities, which leads to an uncertainty of its aetiology and outcome (Dobyns, 1996; O’Driscoll, Black, Clayton-Smith, Sherr, & Dobyns, 2010; Romaniello et al., 2016; Schell-Apacik et al., 2008). The real prevalence in humans is hard to know, even with prenatally CCA diagnostic with
19
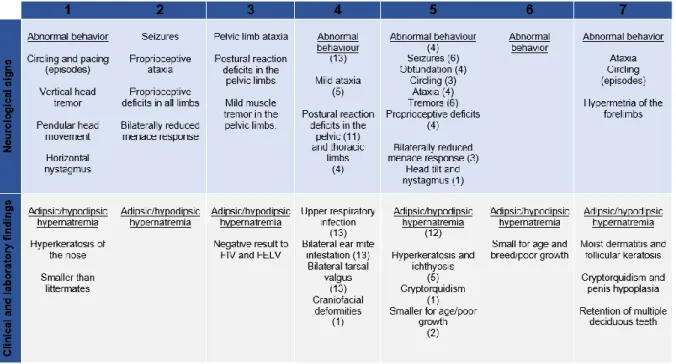
neuroultrasound and fetal MRI (Turkyilmaz et al., 2019), fallible methods like all others. In addition, CCA may be asymptomatic post-birth and a wide severity spectrum of clinical manifestations exists (Bénézit et al., 2015). Nevertheless the highest estimate of CCA prevalence is approximately 7 out of every 1000 births (Henderson, Beerse, Liang, Ferreira, & Wu, 2019) and 3% in the developmentally delayed population (Turkyilmaz et al., 2019). In veterinary medicine CCA are considered to be rare congenital malformations that can occur as isolated entities, although they are more frequently associated with other forebrain congenital abnormalities, including hydrocephalus, septum pellucidum agenesis and abnormal gyral patterns in opposing parasagittal midline gyri, which are also features of HPE (Gonçalves et al., 2014; Vandevelde, Higgins, & Oevermann, 2012). According to recent literature case reports, the most commonly affected dog breeds with CCA and HPE are Staffordshire Bull Terriers and Miniature Schnauzers but other dog breeds may be affected. Reported presentation ages range from 3 to 81 months of age in the most comprehensive study of CCA in dogs (Gonçalves et al., 2014), but this a very variable feature (Hecht, 2018; Shimokawa Miyama et al., 2009; Sullivan et al., 2003; Wang-Leandro et al., 2018). In Table 1, the signalment and respective citation of seven literature case reports of CCA in cats and dogs with CCA, are explicit.
Table 1 – Signalment of seven case reports of cats and dogs with CCA and their respective literature citation.