Elementary screening of lymph node metastatic- related genes in gastric cancer based on the co-expression network of messenger RNA, microRNA and long non-coding RNA
Texto
Imagem
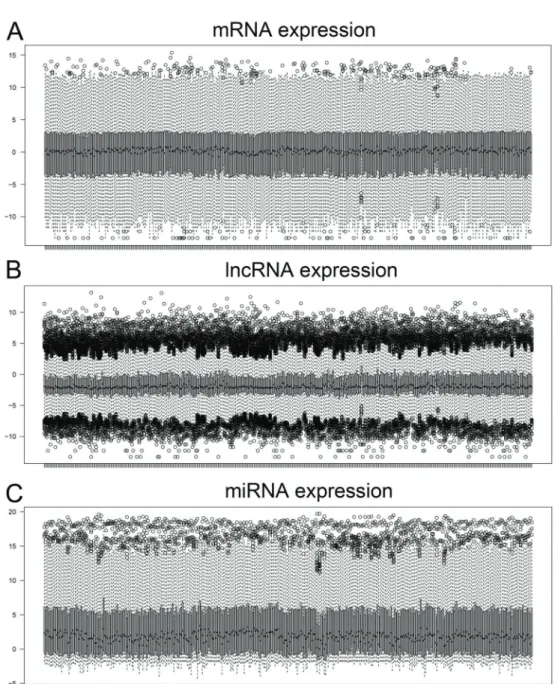

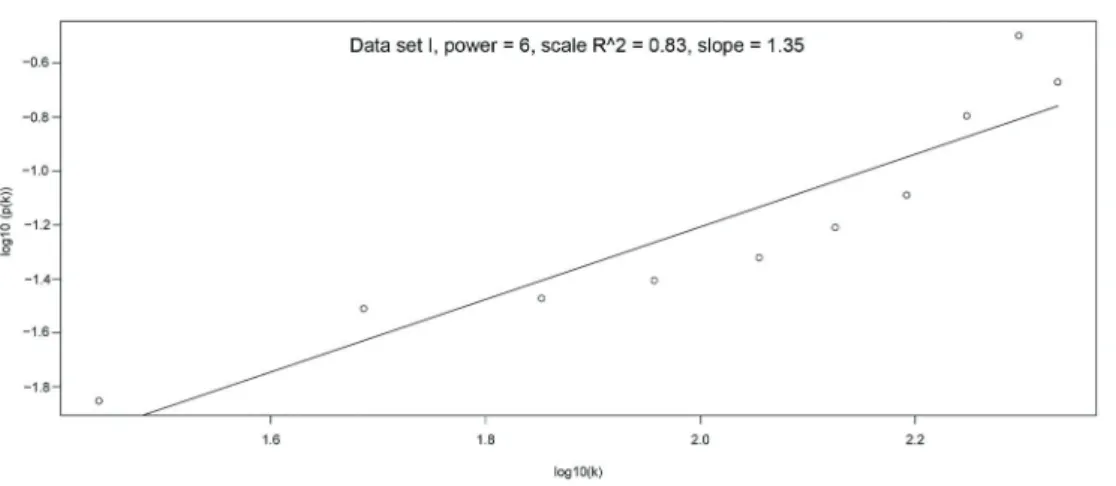

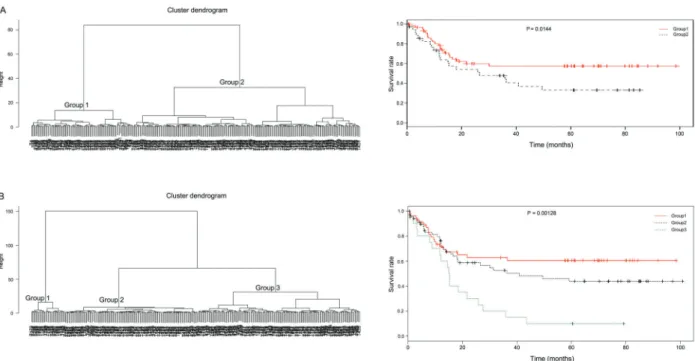
Documentos relacionados
The aim of this study was to investigate the levels of messenger RNA (mRNA) for activin-A on goat preantral and antral follicles and the effects of follicle stimulating
RNA-Seq analysis and identification of differentially expressed genes (DEGs) - We performed RNA-Seq anal- yses to evaluate spatiotemporal gene expression profiles along
E na visão dos beneficiários, os projetos desenvolvidos pelo Espaço Casa Viva possuem impacto em suas vidas, já que, ao se utilizar também o ponto de corte de 0,9 (n≥42)
gostava do seu pai, mas estava, a meu ver zangada pelo amor que gostaria de ter sentido da sua parte e que não sentiu como desejava, ciúme face à relação que o pai tinha
In this study, we used one of these ribozyme variants to target the HIV-1 RNA genome region in the tat gene, and investigated its activity in cleaving the target RNA sequence in
Expressed pseudogenes rather than intron retention or disrupted splice variants were selected (cases where a FANTOM stop codon faces a gap in the Swiss-Prot sequence were
To survey potential fusion genes in GBMs, we analysed RNA-Seq data from 162 GBM patients available through The Cancer Genome Atlas (TCGA), and found that 3 9 exons of
Desse modo, o reconhecimento do papel da escola como esse espaço de manutenção ou superação das desigualdades sociais pode ser um importante meio de construir e pensar sobre
