Toxoplasma gondii in vitro
Texto
Imagem
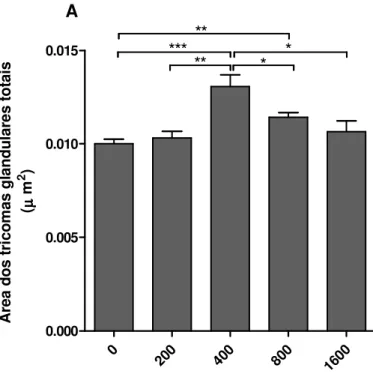
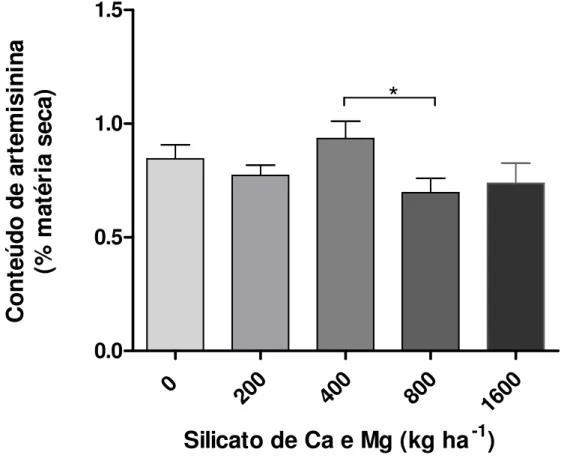
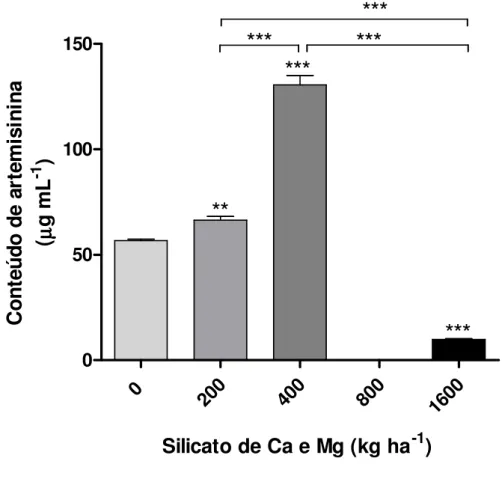
Documentos relacionados
Refere-se a fundamentação teórica, organizado de maneira a transmitir os conceitos utilizados como referência para o desenvolvimento desta pesquisa através de uma
Foram utilizados os marcadores moleculares ISSR, utilizando os primers GACA4 e GTG5, e os marcadores ITS e RAPD, com o objetivo de caracterizar geneticamente linhagens de
Asumiendo el compromiso con una formación en esta línea consideramos que desarrollar el liderazgo situacional supone hacer posible que quien como sujeto se problematiza, y quien
Toxoplasma gondii and Neospora caninum in domestic cats from the Brazilian semi-arid: seroprevalence and risk factors.. [Toxoplasma gondii e Neospora canimum em gatos no
iii) Construir linha em tensão mais elevada no mesmo percurso. iv) Construir nova linha na mesma tensão segundo outro percurso. v) Construir nova linha noutra tensao
Mas que fosse um espaço político sim, mas de discussão de melhoria da ação docente do lado pedagógico (Lenira Paz, F, 2017). Eu penso que esse período todo foi um período
Considering MODIS’s temporal resolution and its potential application to the study of temporal dynamics of the phytoplankton biomass, we aimed at selecting an algorithm
