RAQUEL DA SILVEIRA MAIA
ALTERAÇÕES COGNITIVAS EM RATOS INFECTADOS COM
Toxoplasma gondii
ALTERAÇÕES COGNITIVAS DE RATOS INFECTADOS COM
Toxoplasma gondii
Natal/RN 2012
Dissertação apresentada à Universidade Federal do Rio Grande do Norte, para obtenção do título de mestre em psicobiologia.
Orientador: Prof. Dr. Antonio Pereira Junior
Maia, Raquel da Silveira.
Alterações cognitivas em ratos infectados com Toxoplasma Gondii / Raquel da Silveira Maia. – Natal, RN, 2012.
57 f.: il.
Orientador: Prof. Dr Antônio Pereira Júnior. Coorientador: Profª. Dra. Regina Helena da Silva.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro de Biociências. Programa de Pós-graduação em Psicobiologia.
1. Toxoplasma Gondii – Dissertação. 2. Protozoários – Dissertação. 3. Memória operacional – Dissertação. I. Pereira Júnior, Antônio. II. Silva, Regina Helena da. III. Universidade Federal do Rio Grande do Norte. IV. Título.
RN/UF/BSE-CB CDU 616.993.1
“ Há homens que lutam um dia e são bons. Há outros que lutam um ano e são melhores. Há os que lutam muitos anos e são muito bons.
Porém, há os que lutam toda a vida. Esses são os imprescindíveis”.
Ao meu Orientador, Prof. Dr Antônio Pereira Júnior por acreditar e tornar possível a realização deste projeto.
À minha Co-orientadora, Prof Dra. Regina Helena da Silva pelos importantes apontamentos que fez e que tornaram possível a realização dos experimentos.
À Dra. Alicia Cabral por ter auxiliado na disponibilização de espaço para a realização dos experimentos.
Ao Prof. Dr Valter Ferreira de Andrade Neto por ter disponibilizado espaço e reagentes para a realização dos experimentos.
Ao técnico de laboratório Edson pela amizade e por ter auxiliado no aperfeiçoamento dos aparatos experimentais.
À Capes pelo apoio financeiro
Às companheiras de laboratório Ywlliane e Valeria pela contribuição nos experimentos e por possibilitar a realização deste projeto.
À minhas amigas Danyelle, Norma e Samira que me apoiaram desde o inicio da minha jornada e me animaram nos momentos difíceis.
Ao meu querido namorado, Haroldo Watson, pelo companheirismo e carinho.
Aos colegas do laboratório de biologia da malária e toxoplasmose: Claúdio Bruno, Graciele, Cecilia, Bruno, Camila, José Fernandes, Mariane e Thales
pela amizade.
O Toxoplasma gondii é um protozoário parasito que induz alterações comportamentais em roedores. O objetivo do presente estudo foi avaliar o efeito da infecção pelo T.gondii durante a fase crônica na memória operacional e na impulsividade de roedores, bem como o efeito de antipsicóticos em reverter as eventuais alterações comportamentais decorrentes da infecção. Ratos wistar fêmeas (n=40) foram infectadas com 25 cistos da cepa ME-49 do T. gondii, após 4 meses os animais foram submetidos aos testes comportamentais: tolerância ao retardo de gratificação, na qual o animal deve escolher entre duas recompensas, uma menor imediata e uma maior, mas com retardo e o teste de alternância espontânea, na qual o animal deve utilizar pistas espaciais para recordar braços previamente visitados. Os antipsicoticos foram administrados via intraperitoneal durante a fase teste dos experimentos comportamentais, sendo o antipsicótico haloperidol (1,5 mg/kg) administrado 60 min antes do inicio das sessões e o antipsicótico clozapina (2,5 mg/kg) 30 min antes. Os animais infectados com o parasito não apresentaram déficits de memoria operacional, e não desenvolveram prejuízo motor, no entanto foi observado prejuízo motor apenas nos animais tratados com haloperidol. Foi verificado que a administração de clozapina e haloperidol aumentou a porcentagem de alternação em grupos controle e infectado na tarefa de alternância espontânea.Não existe distinção entre animais do grupo controle e infectado no teste de tolerância ao retardo de gratificação em relação a porcentagem de escolhas pela maior recompensa, durante as fases de pré-treinamento e treinamento, no qual existe um retardo de 15 s para o acesso da grande recompensa, no entanto foi verificado que animais infectados preferem a maior recompensa, quando existe um retardo de 30 s quando comparado ao grupo controle. A administração da clozapina possibilitou que os animais infectados escolhessem a maior recompensa, no retardo de 30s na fase teste. Estes dados sugerem que ratos infectados não apresentam déficits de memória operacional e que a clozapina apresenta eficácia terapêutica em melhorar o desempenho cognitivo de ratos infectados.
Palavras-chave: Toxoplasma gondii, impulsividade, antipsicoticos, memória operacional.
Toxoplasma gondii is a protozoan parasite that induces behavioral changes in rodents. The aim of this study was to evaluate the effect of infection by T. gondii during the chronic phase in working memory and impulsivity in rodents as well as the effect of antipsychotics to reverse any behavioral changes resulting from infection. Female Wistar rats (n = 40) were infected with 25 cysts of the strain ME-49 T. gondii after 4 months the animals were subjected to behavioral tests: tolerance to delay gratification, in which the animal must choose between two rewards, a smaller and more immediate, but delayed and the test of spontaneous alternation, in which the animal must use spatial cues to remember previously visited arms. Antipsychotic drugs were intraperitoneally administered during the testing of the behavioral experiments, the antipsychotic is haloperidol (1.5 mg / kg) administered 60 min before the start of the session and the antipsychotic clozapine (2.5 mg / kg) 30 min before. Animals infected with the parasite did not show operating deficits of memory, and motor impairment did not develop, however motor impairment was observed only in animals treated with haloperidol. It was found that administration of clozapine and haloperidol increased the percentage of alternation in infected and control groups in task switching espontânea.Não no distinction between control animals and infected the test of tolerance to delay gratification in relation to the percentage of choices greatest reward, during the pre-training and training, in which there is a delay of 15 s to access the great reward, however it was observed that infected animals prefer the greatest reward, when there is a delay of 30 s when compared to control group. The administration of clozapine possible that infected animals chose the greatest reward in the delay of 30 seconds during the test. These data suggest that infected mice do not exhibit deficits in working memory and that clozapine has therapeutic efficacy in improving cognitive performance of mice infected.
Lista de Figuras
Figura 1: Formas infectantes do Toxoplasma gondii ... 15
Figura 2: Ciclo biológico do Toxoplasma gondii... 18
Figura 3: Processos envolvidos na memoria operacional... 25
Figura 4: Esquema do Teste de Hemaglutinação Indireta... 30
Figura 5: Esquema do labirinto em cruz elevada... 32
Figura 6: Representação esquemática do labirinto em T... 34
Figura 7: Peso corporal dos animais experimentais... 35
Figura 8: Mudanças do peso corporal dos animais durante os experimentos... 36
Figura 9: Porcentagem de escolhas pela grande recompensa ao final das sessões de pré- treinamento... 37
Figura 10 Desempenho médio dos animais durante as primeiras 8 sessões de pré-treinamento, quando o retardo para a obtenção da recompensa maior está ausente... 37
Figura 11: Porcentagem de escolhas pela grande recompensa ao final das primeiras 5 sessões de treinamento, na qual existe um retardo de 15s para o acesso da grande maior ... 38
Figura 12: Desempenho médio dos animais ao escolher a recompensa maior durante as primeiras 5 sessões de treinamento, quando existe um retardo de 15s... 39
Figura 13: Número de escolhas pela grande recompensa durante sessões de treinamento, na qual existe um retardo de 30s para o acesso à grande recompensa:... 40
escolhas pela grande recompensa... 41 Figura 15: Efeito dos antipsicoticos Haloperidol e Clozapina sobre a alternância espontânea no labirinto em cruz elevada... 42 Figura 16:Efeito dos antipsicóticos Clozapina e Haloperidol sobre o número de entradas nos braços do labirinto em cruz elevada... 43
Lista de Tabelas
AIDS: Síndrome da imunodeficiência adquirida CAE: Comportamento de alternância espontânea GC: Inteligência cristalizada
GF: Inteligência fluida
MCF: Modelo dos cinco fatores de personalidade
T.gondii: Toxoplasma gondii
TMT: Teste das trilhas
WCST: Teste de Wisconsin de classificação de cartas
SUMÁRIO
1.INTRODUÇÃO... 13
1.1 Histórico... 13
1.2 Ciclo de vida... 14
1.3 Alterações comportamentais induzidas por parasitos... 18
1.4 Toxoplasma gondii e Esquizofrenia... 21
1.5 Função Executiva... 21
1.6 Impulsividade... 23
1.7 Memória Operacional... 21
1.8 Os cinco fatores de personalidade... 26
2. Justificativa e Objetivos... 27
3. METODOLOGIA... 28
3.1 Sujeitos experimentais... 28
3.2 Protocolo de infecção... 29
3.3 Teste Sorológico... 29
3.4 Tratamento farmacológico... 30
3.5 Teste de alternância espontânea... 31
3.6 Teste Tolerância ao retardo da gratificação... 32
3.7 Análise estatística... 34
4 RESULTADOS... 35
4.1 Perfil sorológico e Peso corporal... 35
4.2 Teste de Tolerância ao retardo da gratificação... 36
4.3 Teste de Alternância Espontânea... 40
5 DISCUSSÃO... 44
6 CONCLUSÃO... 48
7 Perspectivas ... 49
1-INTRODUÇÃO 1.1- Histórico
O Toxoplasma gondii é um protozoário parasita intracelular altamente prevalente (TENTER, A. M. et al., 2000; PAPPAS, G. et al., 2009) e com um ciclo de vida indireto, onde os felinos são os hospedeiros definitivos e qualquer vertebrado homeotérmico pode ser o hospedeiro intermediário (HUTCHISON, W. M. et al., 1969). O T. gondii foi inicialmente descrito em 1908 por dois pesquisadores franceses, Charles Nicolle e Louis Manceaux, quando estudavam o parasito leishmania no roedor Ctenodactylus gundi no Instituto Pasteur em Tunis, na Argélia (MORRISSETTE, N. S. e AJIOKA, J. W., 2009). No mesmo ano e de maneira independente, Alfonso Splendore, trabalhando no Brasil, descobriu o mesmo parasito em coelhos (DUBEY, J. P., 2008). Charles Nicolle e Louis Manceaux mais tarde denominaram o parasito de Toxoplasma gondii, baseado em sua morfologia (Toxo=arco) e no animal onde foi inicialmente descrito (gondii= gundi).
O potencial patogênico do T. gondii começou a ser revelado na década de 30, com a descrição da encefalomielite decorrente da infeção congênita em fetos em desenvolvimento (WOLF, A. e COWEN, D., 1937). Em seguida, foi descrita a tríade de sintomas associados com a toxoplasmose congênita em humanos, a hidrocefalia, a retinocoroidite e a encefalite (WOLF, A. et al., 1939; INNES, E. A., 2010). O diagnóstico da infecção em seres humanos foi facilitado pelo desenvolvimento de um teste baseado em corante por Albert Sabin e Harry Feldman em 1948 (SABIN, A. B. e FELDMAN, H. A., 1948). A introdução deste teste permitiu demonstrar as altas taxas de prevalência da infecção em seres humanos e animais e estimulou as pesquisas sobre as possíveis rotas de transmissão do parasito, tendo em vista que apenas a transmissão congênita dificilmente resultaria em taxas tão elevadas (INNES, E. A., 2010).
O T. gondii é um parasita extremamente bem sucedido, infectando cerca de 1/3 da população mundial e com ocorrência generalizada em várias regiões do mundo (TENTER, A. M. et al., 2000; MONTOYA, J. G. e LIESENFELD, O., 2004; PAPPAS, G. et al., 2009). Existem várias explicações para o sucesso do T. gondii, entre eles a capacidade de formar oocistos ambientalmente estáveis, a capacidade de infectar uma variedade de hospedeiros homeotérmicos, a capacidade de manipular o sistema imunológico dos hospedeiros, para evitar detecção e resistir à eliminação e, mais intrigante, a capacidade de manipular o comportamento dos seus hospedeiros intermediários.
O impacto da doença em seres humanos é dependente da fase da infecção. A fase aguda da doença decorre da ingestão dos oocistos do parasito ou através da via transplacentária, podendo ser responsável na gravidez pelo aborto espontâneo ou morte fetal. O recém-nascido exposto ao T gondii no útero pode desenvolver toxoplasmose congênita, com consequências neurológicas graves. O ônus da toxoplasmose congênita pode ser representado através do cálculo do número de anos de vida útil perdidos por incapacidade ou mortalidade precoce (DALY, do inglês disability-adjusted life years). É o maior dos patógenos associados com alimentos (HAVELAAR, A. H. et al., 2007). Entretanto, o parasita persiste no interior de células do cérebro e de outros tecidos na forma de bradizoítos é a infecção latente ou crônica que pode ser reativada em casos de imunossupressão, como na síndrome da imunodeficiência adquirida (AIDS), causando encefalite grave (LUFT, B. J. e REMINGTON, J. S., 1992).
A descrição do ciclo de vida do T. gondii ocorreu apenas na década de 1970 (DUBEY, J. P., 2008). Na década seguinte, a epidemia de AIDS e o aumento no número de estudos indicando uma grande variedade de patologias associadas com a toxoplasmose crônica, provocou uma reavaliação da significância clínica da toxoplasmose e do seu impacto na sociedade (DUBEY, J. P., 2008).
O T.gondii apresenta um ciclo de vida heteróxeno no qual a fase assexuada ocorre nos hospedeiros intermediários e a fase sexuada ocorre no intestino de felinos (TENTER, A. M. et al., 2000; DUBEY, J. P., 2009). O T. gondii possui três estágios infectantes: i) o oocisto (10 por 12µm) é a forma proveniente da reprodução sexuada do parasita (gametogonia) no interior das células do epitélio intestinal dos felídeos e possui em seu interior 8 esporozoitos infectantes; ii) o taquizoíto (2 por 6 µm) é uma forma de multiplicação e metabolismo mais rápido sendo produzido pelo ciclo assexuado do parasito em hospedeiros intermediários, é encontrado na fase aguda da infecção; iii) os bradizoítas são formas assexuadas, com metabolismo lento (7 por 1,5 µm) localizados no interior dos cistos teciduais principalmente, durante a fase crônica da infecção (ver Figura 1) (TAKASHIMA, Y. et al., 2008; UNNO, A. et al., 2008; UNNO, A. et al., 2009).
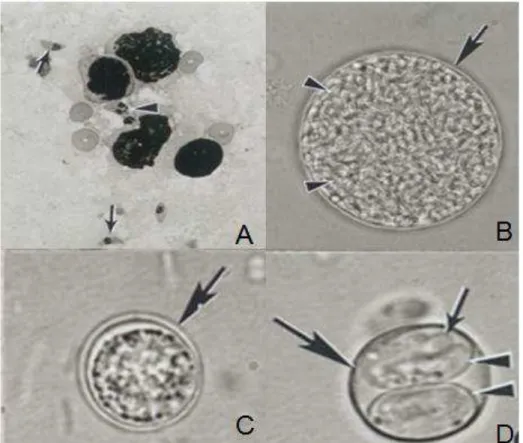
Figura 1: Formas infectantes do Toxoplasma gondii. Em A: taquizoítas presentes no pulmão, B:
cisto tecidual presente no cérebro, C: oocistos esporulados em fezes de felinos, D: oocistos esporulados com esporocistos e no interior de cada um dois esporozoítas.
O ciclo assexuado ocorre quando um hospedeiro intermediário, tal como um roedor selvagem, ou qualquer outro hospedeiro de sangue quente, ingere (i) oocistos esporulados presentes no ambiente, ou (ii) cistos presentes em carne crua ou mal passada, ou (iii) contaminado por via transplacentária (TENTER, A. M. et al., 2000). Mesmo em países desenvolvidos, como o Reino Unido, estudos relatam que 38% das amostras de carne para consumo humano estão infectadas com o T.gondii (ASPINALL, T. V. et al., 2002).
Nos hospedeiros intermediários, o T gondii apresenta duas fases de desenvolvimento assexuadas. Na primeira fase os oocistos e bradizoítas, ao serem liberados no tubo digestivo, sofrem intensa multiplicação intracelular e como taquizoítas, invadirão vários tipos de células dos hospedeiros onde formarão o vacúolo parasitoforo e se multiplicarão rapidamente por endodiogenia repetida formando novos taquizoítas (TENTER, A. M. et al.,2000, NEVES, D.P. et al., 2005).
Posteriormente, com a ação do sistema imunológico, os parasitos extracelulares desaparecem do sangue, da linfa e dos órgãos viscerais, enquanto outros parasitos evoluem para a formação de cistos que contêm em seu interior bradizoítas que permanecem quiescentes, se dividindo lentamente (Neves, D.P. et al., 2005).
A fase cística caracteriza a fase crônica da doença. Os cistos se localizam predominantemente nos músculos e no sistema nervoso central (NATH, A. e SINAI, A. P., 2003). Recentemente, foi demonstrado que os cistos parasitados são encontrados quase exclusivamente no interior de neurônios, não em células de glia, onde podem permanecer por toda a vida do hospedeiro protegidos da ação do sistema imunológico (MELZER, T. C. et al., 2010).
1.2.1- Ciclo sexual
fase assexuada (merogonia) e outra sexuada (gamogonia) (TENTER, A. M. et al., 2000) (ver Figura 2).
Os bradizoítas, taquizoítas ou cistos penetram nas células epiteliais intestinas e se multiplicam por endodiogenia (o parasito se reproduz por um tipo de brotamento, formando duas novas células filhas dentro de cada célula mãe) e merogania (divisão nuclear seguida de divisão do citoplasma, constituindo vários indivíduos isolados simultaneamente), dando origem a vários merozoítas. O conjunto destes merozoítos formados dentro do vacúolo parasitóforo da célula é denominado meronte ou esquizonte maduro. O rompimento da célula parasitada libera os merozoítos, que penetram em novas células epiteliais intestinais e se transformam nas novas formas sexuadas femininas ou masculinas. Esta fase ocorre dois dias após a ingestão de cistos (DUBEY, J. P., 1998).
Os gametas são encontrados no intestino, especificamente no íleo, 3 a 15 dias após a inoculação. Após a fertilização um oocisto é formado, as células epiteliais infectadas se rompem e descarregam os oocistos no lúmen intestinal. O oocisto alcança o meio exterior com as fezes e sua maturação ocorrerá por um processo denominado esporogonia. Após um período de cerca de quatro dias apresentará dois esporocistos, contendo 4 esporozoítas cada (DUBEY, J. P., 1998).
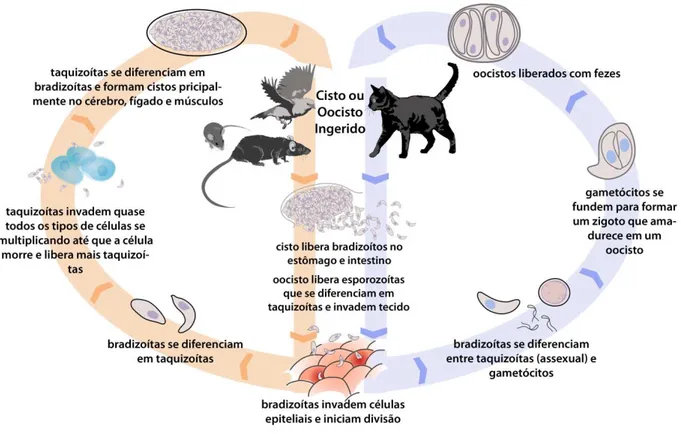
Figura 2: Ciclo biológico do T.gondii.
1.3- Alterações comportamentais induzidas por parasitos
Os parasitas sobrevivem através da exploração de hospedeiros de uma espécie diferente. Esse estilo de vida é provavelmente o mais bem sucedido do planeta, tendo em vista que para cada organismo existe um parasita que pode infectá-lo. Hospedeiros e parasitos co-evoluem em uma disputa perpétua envolvendo adaptações e contra-adaptações de modo a manter suas adequações aos ambientes no qual compartilham. Este raciocínio é capturado na hipótese evolucionária denominada de princípio da rainha vermelha, proposta por Van Valen (VAN VALEN, L., 1973) e que pode ser resumido como: “para um sistema evolucionário, o desenvolvimento contínuo é necessário apenas para manter a sua adequação relativa aos sistemas com os quais co-evolve”. No caso de seres humanos, esse princípio pode ser ilustrado pelo fato de que apesar da imensa alocação somática para o sistema
imunológico no nosso organismo, ainda existe bastante vulnerabilidade às doenças infecciosas.
Vários parasitas evoluíram ciclos de vida complexos, que envolvem múltiplos hospedeiros e são transmitidos para o hospedeiro definitivo de maneira trófica. Aparentemente, existem vantagens adaptativas na introdução de pelo menos um hospedeiro intermediário para a transmissão do parasita até seu alvo final (CHOISY, M. et al., 2003).
Alguns parasitas que são transmitidos de maneira trófica são capazes de alterar o comportamento do hospedeiro intermediário para aumentar chances desse ser predado pelo hospedeiro definitivo (THOMAS, F. et al., 2005). O fenômeno de alteração fenotípica induzida em animais parasitados permite que o parasito passe de um hospedeiro intermediário para o hospedeiro definitivo e desta forma possa completar o seu ciclo de vida (CEZILLY, F. et al., 2010). Existem vários exemplos de manipulação parasitária de hospedeiros disponíveis na literatura, ocorrendo em vários organismos, desde vírus, fungos e bactérias até mamíferos (LEFEVRE, T. et al., 2009). Por exemplo, formigas infectadas com o tremátodo Dicrocoelium dendriticum se deslocam para o topo da vegetação e assim aumentam suas chances de predação pelo hospedeiro definitivo do parasita, as ovelhas (CARREY, W. P., 1969), ou o aumento da atratividade de seres humanos infectados pelo parasito Plasmodium falciparum, causador da malária, para o mosquito Anopheles gambiae, hospedeiro definitivo do parasito e permitindo que o mesmo possa completar o seu ciclo de vida (LACROIX, R. et al., 2005). As modificações fenotípicas induzidas por parasitos são bem variadas, tanto em magnitude quanto em diversidade. Na maioria dos casos, são vários os traços fenotípicos modificados pelo parasito no hospedeiro, em efeito transformando-os em organismos profundamente modificados (THOMAS, F. et al., 2005; THOMAS, F. et al., 2010).
KANNAN, G. et al., 2010; HOUSE, P. K. et al., 2011). Essa alteração comportamental é especifica para a aversão inata aos feromônios do gato presentes na urina, já que não ocorrem alterações em outros comportamentos, tal como medo aprendido ou ansiedade (VYAS, A. et al., 2007).
As indicações iniciais dos mecanismos responsáveis pelas mudanças comportamentais induzidas pelo T. gondii sugeriam a participação de vias dopaminérgicas. Foi demonstrado, por exemplo, que a administração do antipsicótico haloperidol e do ácido valpróico, um estabilizador do humor, poderiam evitar o aparecimento dessas mudanças comportamentais (WEBSTER, J. P. et al., 2006). Adicionalmente, foi demonstrado que a administração de um bloqueador seletivo de autoreceptores dopaminérgicos, a vanoxerine, (GBR- 12909) também causava supressão dos comportamentos atípicos associados com a infecção crônica pelo T.gondii em roedores (SKALLOVA, A. et al., 2006). Também demonstraram que antipsicóticos e estabilizadores de humor têm ação inibitória sobre a replicação de taquizoítas in vitro (JONES-BRANDO, L. et al., 2003).
Os cistos de T.gondii se localizam em regiões estratégicas para a manipulação do comportamento no sistema nervoso central de mamíferos, como no sistema límbico (HOLLIMAN, R. E., 1997), sistema olfatório, córtex pré-frontal, hipotálamo, amigdala lateral e basolateral (VYAS, A. et al., 2007). O sistema límbico é uma região envolvida na mediação de vários comportamentos sociais, tal como: agressividade, reprodução e comportamentos parentais (KLEIN, S.L., 2003). Recentemente foi demonstrado que ratos infectados cronicamente com T.gondii ao serem expostos ao odor do gato apresentam aumento da atividade neural localizada numa região da amigdala normalmente ativada quando ratos não infectados são expostos a uma fêmea no estro (HOUSE, P. K. et al., 2011).
indicando um possível mecanismo para a manipulação comportamental observada em hospedeiros intermediários do parasito (PRANDOVSZKY, E. et al., 2011).
1.4- Toxoplasma gondii e esquizofrenia
Vários estudos disponíveis na literatura têm sugerido que o risco do desenvolvimento de esquizofrenia é significativamente aumentado com a exposição pré-natal/perinatal a vários insultos ambientais, como desnutrição (BROWN, A. S. et al., 1996), complicações obstétricas, exposição materna ao estresse e infecção (BROWN, A. S. et al., 2009), etc. Esses estudos sugerem que os insultos ambientais interferem com o desenvolvimento normal do cérebro precoce e dão suporte à teoria de que a esquizofrenia e outros distúrbios psicóticos associados derivam de uma combinação de susceptibilidade genética e agentes ambientais (LEWIS, D. A. e LEVITT, P., 2002).
Estudos demonstram uma associação entre a infecção por Toxoplasma gondii e esquizofrenia. Foi demonstrado que alguns casos de toxoplasmose aguda estão associados em humanos adultos estão associados com sintomas psiquiátricos, tais como: desilusão e alucinações (TORREY, E. F e YOLKEN, R.H., 2003), além disso análises preliminares indicam um aumento dos anticorpos anti- T.gondii (IgG e IgM) em mulheres durante a gestação e que posteriormente a sua prole desenvolveu esquizofrenia na fase adulta (BUKA, S. L. et al., 1999).
1.5- Funções Executivas
al., 2000) são: inibição de interferência ou de resposta prepotente (controle da impulsividade), flexibilidade de controle mental e memória operacional.
Na clínica, as funções executivas são avaliadas com tarefas como o Teste Wisconsin de Classificação de Cartas (WCST, do inglês Wisconsin Card Sorting Test), a Torre de Hanoi ou a sua variante, Torre de Londres e pacientes com lesão no lobo frontal tendem a apresentar deficiências no desempenho dessas tarefas (MIYAKE, A. et al., 2000).
Estudos clínicos demonstram a existência de modulação das funções executivas por parasitos. Estudo com pacientes esquizofrênicos infectados com citomegalovírus e HSV1 apresentam uma maior taxa de erro no Teste de Trilha (TMT, do inglês Trail Making Test) (SHIRTS, B. H. et al., 2008). Outro estudo com pacientes esquizofrênicos demonstra que a exposição intrauterina às infecções, com o Toxoplasma gondii ou Influenza está relacionada com uma maior taxa de erro nos testes de WCST (BROWN, A. S. et al.,2009).
O comportamento inteligente apresenta várias demandas importantes de controle de sub-processos cognitivos para o alcance dos seus objetivos. Como esperado, vários estudos demonstram que o córtex frontal/pré-frontal proporciona um substrato unificado para a inteligência humana (DUNCAN, J. et al., 2000; BISHOP, S. J. et al., 2008; ROCA, M. et al., 2010).
Foi demonstrado que seres humanos infectados com Toxoplasma gondii apresentam menor QI se comparado com indivíduos não infectados, bem como apresentam uma menor probabilidade de atingir o ensino superior (FLEGR, J. et al., 2003). Foi verificada uma associação entre toxoplasmose congênita e diminuição do QI em crianças infectadas não tratadas, quando comparado ao grupo controle (SAXON, S. A. et al., 1973).
1.6- Impulsividade
A impulsividade é um construto psicológico muito importante dos sistemas teóricos de personalidade (DICKMAN, S. J., 1990; WHITESIDE, S. P. e LYNAM, D. R., 2001) e desempenha um papel de destaque no diagnóstico de várias psicopatologias, como transtorno de personalidade borderline, transtorno de personalidade antissocial, transtorno de déficit de atenção/hiperatividade, mania, demência e bulimia nervosa (ASSOCIATION, A. P., 1995). A impulsividade pode ser definida como um comportamento de ação rápida sem planejamento ou julgamento consciente e inadequado ao contexto (MOELLER, F. G. et al., 2001). Um outro componente importante no comportamento impulsivo é a intolerância ao retardo de recompensa (ARCE, E; SANTISTEBAN, C., 2006; BIZOT, J. C. et al., 2007).
Pessoas impulsivas ou com baixo controle têm dificuldade em imaginar cenários que incluem consequências negativas (COOLS, R., 2010), o que pode explicar a relação entre a impulsividade e o abuso de substancias e outros comportamentos aditos (ALESSI, S.M e PETRY, N.M., 2003; DAWE, S e LOXTON, N.J., 2004).
valor de recompensas futuras, sendo responsável pela impaciência de curto prazo.
A paciência para recompensas de longo prazo é mediada pelo córtex prefrontal lateral e pelo córtex parietal posterior, que são capazes de avaliar as diferenças entre recompensas abstratas, incluindo recompensas no futuro mais distante (MCCLURE, S. M. et al., 2004).
Sabe-se que pacientes com lesões no córtex pré-frontal têm problemas em prever consequências negativas em uma tarefa, se limitando a aspectos imediatos (BECHARA, A. et al., 2000). Estudos em modelos animais demonstraram que lesões no córtex orbito frontal e na amígdala resultam na escolha sistemática por uma pequena recompensa, mas disponível de maneira mais imediata, do que uma grande recompensa com acesso retardado (WINSTANLEY, C. A. et al., 2004).
Existem evidências de que a quantidade de receptores dopaminérgicos em vias mesolímbicas pode estar relacionado com a impulsividade. Por exemplo, estudo demonstra a existência de uma correlação entre a disponibilidade de receptores dopaminérgicos D2 e D3 no córtex temporal no tálamo de humanos com o grau de impulsividade (BERNOW, N. et al., 2011). A relação entre a modulação das vias dopaminérgicas na expressão do fenótipo impulsivo pode ser demonstrado também em modelos murinos, no qual foi verificado que a administração de agonistas dopaminérgicos, tais como metilfenidato ou anfetamina aumentam a frequência de escolha por uma recompensa grande que apresenta um retardo, em relação a uma pequena recompensa imediata (GAALEN, M. M. V. et al., 2006)
A impulsividade é um processo multifacetado, com substratos neurais distintos que interagem para determinar o fenótipo impulsivo (KERTZMAN, S. et al,. 2006). A impulsividade em seres humanos está relacionada com polimorfismos nos genes que codificam o receptor de dopamina do tipo D4 (DRDA) (COMINGS, D. E. et al., 1999) e ao transportador de serotonina (5-HTTLPR, do inglês 5-HT transporter gene-linked polymorphic region) (KIRBY, L. G. et al., 2011).
1.7- Memória Operacional
A memória operacional é uma parte do sistema de memória que armazena temporariamente e manipula informações importantes para organizar a cognição, desempenhando um papel crucial no aprendizado, no raciocínio e na linguagem (BADDELEY, A., 2003) (Ver figura 3).
Figura 3: Processos envolvidos na memória operacional. A memória sensorial está associada percepções automáticas e geralmente desaparece em poucos segundos, a memória operacional depende da atenção e dos elementos da memória sensorial e é capaz de reter a informação por poucos segundos.
Fonte: thebrain.mcgill.ca
As aferências dopaminérgicas da via mesocortical para o córtex pré-frontal desempenham um papel crucial nos processos cognitivos normais e patológicos associados com as funções executivas. Em especial, a dopamina regula a função da memória operacional, estabilizando a atividade dos neurônios que codificam itens de memória até que uma resposta adequada seja executada (SEAMANS, J. K. e YANG, C. R., 2004).
baixos níveis de dopamina mesocortical (ABI-DARGHAM, A. et al., 2002). Resultados semelhantes também foram observados em macacos com depleção experimental de dopamina no córtex pré-frontal (BROZOSKI, T. J. et al., 1979). Estudos apontam que a infecção pelo Toxoplasma gondii esta relacionada com déficit de memória operacional em ratos e camundongos (WITTING,P.A.,1979; KANANN, G. et al.,2010), isto pode ocorrer devido a presença de cistos no córtex pré-frontal (VYAS, A. et al., 2007).
1.8- Os Cinco Fatores de Personalidade
O modelo dos cindo fatores de personalidade (MCF) é considerado por muitos psicólogos como a melhor representação da estrutura de traços de personalidade (DIGMAN, J. M., 1990). De acordo com o MCF, a maioria dos traços de personalidade podem ser descritos em termos de um conjunto de cinco grande fatores: extroversão, socialização, escrupulosidade, neuroticismo e abertura para novas experiências (DIGMAN, J. M., 1990; HUTZ, C. S. et al., 1998). Cada um desses fatores representa um padrão de covariância entre um grupo maior de traços de personalidades mais específicos. A universalidade desta estrutura de traços de personalidade foi testada em várias culturas, de cinco famílias de línguas distintas, com resultado positivo (MCCRAE, R. R. e COSTA, P. T., JR., 1997; HENDRYKS, A. A. J. et al., 2003).
Existem vários trabalhos que demonstram uma associação entre o traço de escrupulosidade e a performance profissional (SALGADO, J. F., 2001; WITT, L. A. et al., 2002) e acadêmica (HAIR, P. e HAMPSON, S. E., 2006; POROPAT, A. E., 2009). A impulsividade é um dos traços definidores mais importantes da escrupulosidade (HAIR, P. e HAMPSON, S. E., 2006). Além disso, a escrupulosidade é altamente correlacionada com a performance no treinamento de uma tarefa de memória operacional (STUDER-LUETHI, B. et al., 2012).
Toxoplasma gondii acarreta aumento de extroversão e diminuição da conscientização, quando comparado ao grupo controle (LINDOVA, J. et al., 2012).
2- Justificativa e Objetivos
Estudos anteriores têm demonstrado alteração do repertorio comportamental normal de ratos e camundongos infectados com Toxoplasma gondii. As alterações observadas podem estar associadas com alterações nas vias dopaminérgicas determinadas pela presença de cistos no tecido nervoso, tais como em áreas límbicas. A modulação dopaminérgica efetuada por cistos no córtex pré-frontal pode ocasionar alterações nas funções executivas, tais como memória operacional e impulsividade. Diante disto o presente trabalho teve como objetivo avaliar se a infecção por Toxoplasma gondii na fase crônica resulta em modificações nas funções executivas, particularmente impulsividade e memória operacional, bem como determinar o efeito de antagonistas dopaminérgicos em reverter alterações na memoria operacional e impulsividade em ratos. Especificamente foi avaliado:
Avaliar o efeito da infecção crônica com o T. gondii no teste de tolerância ao retardo de gratificação, que mede impulsividade.
Avaliar o efeito da infecção crônica com o T. gondii no desempenho em uma tarefa de alternância espontânea, dependente de memória operacional
Verificar se a administração prévia dos antipsicóticos clozapina e haloperidol tem efeito no comportamento de tolerância ao retardo de gratificação em ratos controle e infectados.
3- METODOLOGIA
3.1- Sujeitos Experimentais
Foram utilizados 40 ratos Wistar fêmeas com 4 meses de idade (140-245 g) (Ver tabela 1). Todos os animais foram mantidos no biotério do laboratório de biologia da malária e toxoplasmose do Centro de Biociências da Universidade Federal do Rio Grande do Norte, sendo submetidos a um ciclo de claro e escuro de 12h/12h (início da fase clara: 06:00 h) e à temperatura ambiente constante de 22 ± 1°C, com fornecimento de água ad libitum.
Os animais submetidos ao teste de tolerância ao retardo de gratificação tiveram controle alimentar organizado da seguinte maneira: 10g/rato/dia de ração na primeira semana da fase de pré-treinamento e 12 a 15g /rato/dia de ração a partir da primeira semana de treinamento até o termino do experimento. Os animais utilizados nos testes de alternância espontânea tiveram acesso ad libitum ao alimento.
Tabela 1: Numero de animais por grupo utilizados durante as análises comportamentais.
3.2-Protocolo de Infecção
deste foi retirado 25µl que foi colocado em uma lâmina e visualizado ao microscópio para contabilizar os cistos. Posteriormente, foi feito uma diluição com PBS para adequar a quantidade de cistos ao número desejado e os animais experimentais foram infectados via oral por meio de gavagem. Os animais controle receberam via oral 200µl de solução salina.
3.3-Testes Sorológicos
Para o diagnóstico sorológico dos animais foi coletado 500µl de sangue por meio de punção da veia caudal lateral antes do processo de infecção, o mesmo procedimento foi repetido ao término das análises comportamentais, com a coleta de sangue sendo realizada a partir da punção intracardíaca.
O diagnóstico sorológico foi realizado por meio do teste qualitativo hemaglutinação indireta, que se baseia na aglutinação de hemácias sensibilizadas com o antígeno de T. gondii, em presença de soro contendo anticorpos contra este parasita. O teste é dividido em três etapas. Na etapa 1 adiciona-se a amostra, que sendo reagente, contém anticorpos anti-T. gondii. O suporte é uma placa de microtitulação com 96 cavidades. Na etapa 2 adiciona-se as hemácias adiciona-sensibilizadas com antígeno anti- T.gondii. A reação antígeno- anticorpo (etapa 3) é visualizada pela aglutinação das hemácias (BRASIL, 1998).
Figura 4: Testes de hemaglutinação indireta. A) Representação esquemática de uma reação de HAI, B) Visualização da amostra reagente, não reagente e indeterminada na reação de HAI.
Para a coleta de sangue, os animais foram anestesiados com 10mg/kg
de Cloridrato de Xilazina a 2% e 75mg/kg de Cloridrato de Cetamina a 10%, via intramuscular. O sangue coletado com heparina foi centrifugado à uma temperatura de 21°C à 3000 rpm por 15 min, para a obtenção de soro contendo os anticorpos, que foi armazenado a - 20°C até o momento de realização do teste de hemaglutinação indireta.
3.4- Tratamento Farmacológico
As drogas utilizadas foram a clozapina (2,5mg/kg) e o haloperidol (1,5mg/kg). A clozapina foi diluída em Tween a 10%, enquanto o haloperidol foi diluído em solução salina 9%. A solução controle consistia de salina 9%. Ambas as soluções foram administradas via intraperitoneal em um volume de 500µl. A administração da Clozapina ocorreu 30 min antes do inicio dos testes comportamentais, enquanto que a administração do haloperidol ocorreu 60 min antes.
A
B
3.5- Teste de Alternância Espontânea
O teste de alternância espontânea é um teste comportamental que avalia a memória operacional (HUGHES, R. N., 2004). O comportamento de alternância espontânea (CAE) é uma tendência de roedores e outros animais em alternar as escolhas não-reforçadas dos braços de um labirinto em cruz, T, ou Y em oportunidades sucessivas. O teste de alternância espontânea também avalia a locomoção do animal, que pode ser medida pelo número de entradas nos braços.
O teste de alternância espontânea foi realizado após 4 semanas de infecção com o T. gondii. Os animais foram colocados individualmente em um labirinto em forma de cruz (ver Figura 5) constituído de 4 braços fechados (0,47 x 0,16 x 0,34 m), com um telhado e um espaço central abertos (Plataforma central 0,16 x 0,16 cm). O labirinto era elevado a uma altura de 50 cm do chão.
Figura 5: Representação esquemática do labirinto em cruz elevada.
3.6- Teste de Tolerância ao Retardo da Gratificação
O teste de tolerância ao retardo de gratificação mede o comportamento impulsivo em animais experimentais, através da capacidade de esperar por uma recompensa alimentar (THIEBOT, M. H. et al., 1985; BIZOT, J. C. et al., 2007). Os ratos são treinados em um labirinto em T para escolher entre 2 recompensas: uma pequena, mas imediatamente disponível, e outra grande, mas com acesso retardado. Animais impulsivos apresentam uma tendência de escolher o braço com a menor recompensa, ao invés de esperar por uma recompensa maior, mas retardada.
recompensa é colocada na extremidade de cada um dos braços: um dos braços é associado com a recompensa pequena (2 pellets de ração) e o outro braço com a recompensa maior (10 pellets de ração). O procedimento experimental foi dividido em 4 etapas: Habituação, pré-treino, treino e teste (THIEBOT, M. H. et al., 1985; BIZOT, J. C. et al., 2007).
Durante a habituação os animais foram submetidos a 2 sessões (em dias distintos), na qual permaneciam no aparato por um período de 5 minutos e eram posteriormente recolocados na gaiola. Nesta etapa os animais podiam explorar livremente o aparato e se alimentar dos pellets em qualquer um dos braços.
Após a fase de habituação os animais foram submetidos à fase de pré- treino. Após a porta ser inserida na fenda C2, o animal é colocado na base do
braço vertical e quando escolhe um dos braços para se alimentar, a porta em C1 é inserida e a porta em C2 é removida (ver figura 4). O animal é removido do
aparato após se alimentar dos pellets de ração e é colocado na gaiola por um período de 2 a 3 min (intervalo entre testes). Após este intervalo o animal retorna para o aparato. Cada animal será submetido a este procedimento 4 vezes por dia, constituindo uma 1 sessão. Após o animal selecionar o braço com maior recompensa em 75% dos testes, será iniciada a fase de treino. Na fase de treino, após a porta ser inserida em C2, o animal é colocado
na base do braço vertical e quando escolhe um dos braços para se alimentar, a porta em C1 é posicionada de modo que o rato que seleciona o braço de maior
grupo recebeu, 60 minutos antes da sessão, também por via intraperitoneal, o antipsicótico haloperidol.
3.7-Análise Estatística
No experimento de tolerância ao retardo da gratificação, o efeito da infecção sobre a expressão do comportamento impulsivo foi avaliado pelo teste t com amostras não pareadas (etapas de pré-treinamento e treinamento), e o efeito entre as sessões foi realizado com análise de variância (ANOVA) de amostras de medida repetida seguida pelo teste bonferroni, enquanto que a análise dos efeitos dos antipsicóticos (clozapina e haloperidol) sobre o comportamento impulsivo e memória operacional foi realizada ANOVA, seguido pelo teste de Bonferroni. Para comparar o desempenho de ratos no teste de alternância espontânea, foi feita uma análise por meio do teste t com amostras não pareadas. Os resultados foram expressos em média ± E.P. O nível de significância considerado, em todos os testes foi de 5%.
4-Resultados
4.1- Perfil sorológico e Peso corporal
O teste de hemaglutinação indireta confirmou a presença de anticorpos anti-T.gondii em todos os animais que haviam sido infectados experimentalmente, no entanto não foi detectada a presença de anticorpos em animais controle. Foi observada que a média do peso corporal dos animais infectados é maior do que nos animais do grupo controle (p< 0,001) (Ver figura 7). Não foi observada diferença significativa no peso corporal dos animais do grupo infectado no decorrer dos experimentos, no entanto os animais do grupo controle tiveram um aumento do peso ao termino dos experimentos (p< 0,001) (Ver figura 8).
4.2- Teste de Tolerância ao Retardo de gratificação (Impulsividade).
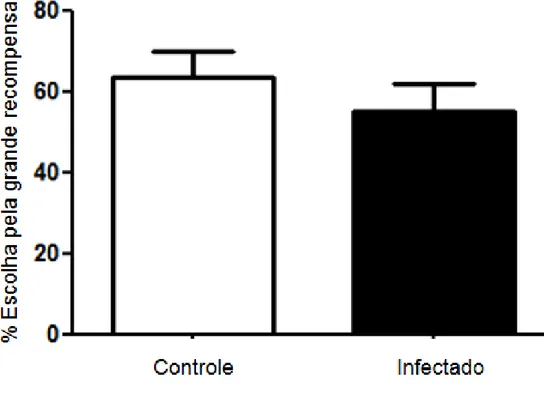
Figura 9: Porcentagem de escolhas pela grande recompensa ao final das sessões de pré-treinamento.
Figura 10: Desempenho médio dos animais durante as primeiras 8 sessões de
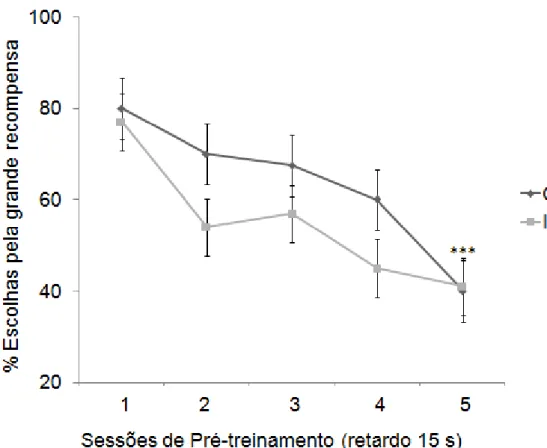
Ao final da fase de treinamento também não foi observada diferença significativa entre os grupos controle e infectado com relação à escolha pela recompensa maior, no retardo de 15s (p=0,09) (ver Figura 11), no entanto foi verificado um decréscimo significativo na escolha pela grande recompensa na 5 sessão pelo grupo controle (Ver figura 11). Foi observada diferença significativa entre os grupos controle e infectado (p< 0,0001) (ver Figura 12) com relação à escolha pela grande recompensa com um retardo de 30s, favorecendo o grupo infectado.
Figura 11: Porcentagem de escolhas pela grande recompensa ao final das primeiras 5 sessões
Figura 13: Número de escolhas pela grande recompensa durante sessões de treinamento, na qual existe um retardo de 30s para o acesso à grande recompensa ( ***p<0,001 quando comparado ao grupo controle).
Figura 14: Efeito dos antipsicoticos Haloperidol e Clozapina na porcentagem escolhas pela grande recompensa. ( *p<0,5 quando comparado ao grupo infectado tratado com salina, ** p<0,01, quando comparado ao grupo ao grupo infectado tratado com haloperidol).
4.3- Resultados do Teste de Alternância Espontânea
5-Discussão
Os nossos resultados demonstram que a infecção pelo T.gondii não modifica o desempenho comportamental de ratos infectados em tarefas que dependem de capacidade de memória operacional e do controle da impulsividade. Os resultados também indicam que os déficits observados não são de origem motora, já que a frequência de entradas nos braços do labirinto não difere entre os grupos analisados, ao contrario de um estudo prévio em um labirinto de três braços, onde camundongos infectados com a cepa ME- 49 apresentam déficits de memória operacional em uma tarefa de alternância espontânea (KANNAN, G. et al.,2010).’ Foi verificado que animais infectados erram com maior frequência em tarefas realizadas em um labirinto com 8 braços, no qual o animal deve recordar a localização de uma recompensa no final de três braços (HODKOVA, H. et al., 2007). No entanto foi demonstrado que camundongos infectados apresentam desempenho cognitivo normal e não desenvolvem déficits de memória espacial (GULINELLO,M. et al.,2010).
Como mencionado anteriormente o nosso estudo demonstrou a ausência de prejuízos motores decorrentes da infecção crônica com o T.gondii. Corroborando os nossos dados, outros estudos também verificaram a ausência de efeitos do T.gondii, no comportamento exploratório e na distância percorrida em um campo aberto em ratos infectados (VYAS, A. et al., 2007). Entretanto, outros estudos demonstram decréscimo na atividade motora de camundongos infectados durante tarefa de campo aberto (GULINELO, M. et al., 2010; KANNAN, M. J., 2010; SKALLOVA, A. et al., 2006).
Os nossos dados demonstram que a administração prévia dos antipsicóticos clozapina e haloperidol revertem as alterações na memória operacional causadas pela infecção crônica com o T. gondii. Essa modificação se reflete em um aumento na porcentagem de alternância entre os braços do aparato em cruz elevado. O efeito da clozapina sobre a memória operacional já foi demonstrada em estudos prévios em pacientes esquizofrênicos que apresentaram melhoras no desempenho de memória operacional verbal após 6 meses de tratamento (HAGGER, C. et al., 1993; LEE, M. A. et al., 1994). Estudos de ressonância magnética em pacientes esquizofrênicos que são tratados com o antipsicótico clorpromazina demonstram melhoras na memória operacional associado com um aumento de atividade no córtex pré-frontal dorso lateral, provavelmente relacionado com ação nos receptores dopaminérgicos D2 (HONEY, G. D. et al., 1999).
Não foi observado prejuízo motor nos grupos que receberam clozapina durante os testes de alternância espontânea (ver Figura 16). Os nossos dados corroboram estudos clínicos que demonstraram que a administração de antipsicoticos em pacientes esquizofrênicos está associada com baixa incidência de efeitos extrapiramidais (ROSENHECK, R. et al., 1997). Estudos com modelos murinos demonstram que a administração de baixas doses de clozapina, semelhantes àquelas utilizadas no presente estudo (2, 5 mg/ kg e 5mg/kg), não apresentam efeito cataléptico (BATOOL, F. et al., 2000).
Nossos resultados também demonstram que ratos infectados cronicamente com o T. gondii e que receberam haloperidol apresentaram deficit locomotor, demonstrado pelo menor número de entradas nos braços do labirinto em relação ao controle. Os nossos dados corroboram estudos clínicos anteriores com pacientes esquizofrênicos que relatam um aumento na incidência de sintomas parkinsonianos em indivíduos tratados com haloperidol (KNABLE, M. B. et al., 1997; SIKICH, L. et al., 2004).
A diferença observada entre os grupos tratados com haloperidol e clozapina com relação aos efeitos extrapiramidais pode ser explicada pela ocupação diferenciada de receptores dopaminérgicos D2. Antipsicoticos típicos
al.,2000), sendo que a probabilidade de efeitos colaterais, tais como efeitos extrapiramidais aumentam significativamente quando a ocupação de receptores D2 ultrapassa os 78% (KAPUR, S. et al., 2000). Já os antipsicoticos
atípicos apresentam maior afinidade por receptores 5HT2a e menor afinidade
por receptores D2 (MELTZER, H. Y. et al., 1989; e SCHOTTE, A. et al., 1996).
O teste de tolerância ao retardo da gratificação pode revelar traços de impulsividade em animais experimentais, tendo em visto que o animal deve escolher entre duas opções: uma recompensa pequena e imediata ou uma recompensa grande, mas com retardo para sua obtenção (BIZOT, J. C. et al., 2007).
Os resultados do nosso estudo demonstraram que não ocorre diferença significativa na frequência da escolha pela grande recompensa entre animais controle e infectados nas sessões de pré-treinamento, caracterizada pela ausência de retardo. Da mesma forma, não foi observada diferença significativa entre os dois grupos durante as primeiras sessões de treino, quando é imposto um retardo de 15s para o acesso à recompensa maior. Entretanto, a diferença entre os dois grupos se revela durante as sessões de treino, no qual é interposto um retardo de 30s para o acesso à grande recompensa: os animais infectados preferem o acesso a grande recompensa, mesmo com a presença do retardo de 30s, demonstrando que ratos infectados apresentam uma tendência para menor impulsividade. Este resultado corrobora estudos realizados com seres humanos infectados com o T. gondii que apresentaram escores significativamente menores do que sujeitos não-infectados em testes que avaliam procura de novidade, impulsividade, extravagância e desorganização (FLEGR, J. et al., 2003; SKALLOVA, A. et al., 2005).
Estudos anteriores demonstraram que pacientes esquizofrênicos tratados com clozapina apresentavam redução do comportamento impulsivo em relação ao grupo tratado com haloperidol. (BARUCH, S. et al., 2003; DURSUN, S.M. et al., 2000).
A ação de antipsicóticos sobre agentes infecciosos é bem conhecida há vários anos (JONES-BRANDO, L. et al., 2003). Por exemplo, foi observado que o haloperidol e o estabilizador de humor ácido valpróico inibem a replicação de taquizoítas do T. gondii em cultura de células (JONES- BRANDO, L. et al., 2003). Além disso, foi demonstrado que o haloperidol, o ácido valpróico, bem como a associação de piremitamina com dapsona, reduzem o comportamento de atração pela urina de gatos observada em por ratos infectados com o T. gondii (WEBSTER, J. P. et al., 2006).
A clozapina é um antagonista tanto de receptores serotoninérgicos (5HT2A) quanto dopaminérgicos (STAHL, S.M e MUNTNER, N., 2006). Estudos anteriores demonstraram que os antipsicóticos atípicos apresentam menor afinidade por receptores D2, assim como possuem uma cinética de dissociação rápida destes receptores. O efeito da clozapina sobre os receptores serotoninérgicos pode ser uma parte importante do mecanismo responsável pelo seu potencial terapêutico, relacionado com a redução do comportamento impulsivo. Pacientes esquizofrênicos tratados com clozapina apresentavam altas concentrações plasmáticas de 5-HT, bem como aumento de 5- HIAA, quando comparado aos grupos controle, além de maior impulsividade (DURSUN, S.M. et al., 2000).
6-Conclusão
7- Perspectivas
7-Referências
ABI-DARGHAM, A. et al. Prefrontal dopamine D1 receptors and working memory in schizophrenia. J Neurosci, v. 22, n. 9, p. 3708-19, May 1 2002.
ALESSI, S. M.; PETRY, N. M. Pathological gambling severity is associated with impulsivity in a delay discounting procedure. Behavioral Process, v. 64, p. 345-354, Jun 2003.
ARCE, E,; SANTISTEBAN, C. Impulsivity: a review. Psicothema, v. 18, n.2, p: 213-220, May 2006.
ASPINALL, T. V. et al. Prevalence of Toxoplasma gondii in commercial meat products as monitored by polymerase chain reaction--food for thought? Int J Parasitol, v. 32, n. 9, p. 1193-9, Aug 2002.
ASSOCIATION, A. P. Manual diagnóstico e estatístico de transtornos mentais - DSM IV. 4ª ed. Porto Alegre: Artes Médicas, 1995.
BADDELEY, A. Working memory: looking back and looking forward. Nat Rev Neurosci, v. 4, n. 10, p. 829-39, Oct 2003.
BADDELEY, A.; HITCH, G. J. Working memory. New York: Academic Press, 1974.
BADDELEY, A.; HITCH, G. J. Working memory. Scholarpedia, v. 5, n. 2, 2010.
BARUCH, S. et al. The effects of clozapine versus haloperidol on measures of impulsive aggresion on suicidality in chronic schizophrenia patients: an open, nonrandomized, 6 month study. The J Clin Psychiatry, v. 64, p. 755-760, 2003.
BRASIL, Ministerio da Saúde, Coordenação Nacional de doenças Sexualmente Transmissíveis e Aids. Doença de chagas. Triagem e diagnóstico sorológico em unidades hemoterapicas e laboratórios de sáude pública. Brasília, (DF);1998.
BROWN, M. D. et al. Maternal exposure to toxoplasmosis and risk of schizophrenia in adult offspring. Am J Psychiatry. v. 162, p. 767-773, 2005. BECHARA, A. et al. Characterization of decision- making defict of patients with ventromedial prefrontal cortex lesions. Brain, v.123, p. 2189-2202, 2000.
BERRIDGE, K. C.; ROBINSON, T. E. Parsing reward. Trends in Neurosci, v. 26, n. 9, p. 507- 513, Sep 2003.
BERNOW, N. et al. Dopamine D2/D3 receptor availability and venturesomeness. Psychiatry Research: Neuroimaging, v. 193, p. 80-84, Jul 2011.
BISHOP, S. J. et al. COMT val158met genotype affects recruitment of neural mechanisms supporting fluid intelligence. Cereb Cortex, v. 18, n. 9, p. 2132-40, Sep 2008.
BIZOT, J. C. et al. Effects of imipramine- like drugs and seretonin uptake blockers on delay of reward in rats. Possible implication in the behavioral mechanism of action of antidepressants. J Pharmacol and Exp Ther, v. 246, p. 1144-1151, Jun 1988.
BIZOT, J. C. et al. Seretonin and tolerance to delay of reward in rats. Psychophamacology, v. 146, p. 400-412, Jun 1999.
BIZOT, J. C. et al. Methylphenidate reduces impulsive behaviour in juvenile Wistar rats, but not in adult Wistar, SHR and WKY rats. Psychopharmacology (Berl), v. 193, n. 2, p. 215-23, Aug 2007.
BOOTOL, F. et al. Neurochimical and exptrapyramidal effects of atypical neuroleptic clozapine in rats. Pak J Pharm Sci, v. 31, n. 1, p. 47-55, Jan 2000. BROWN, A. S. et al. Neurobiological plausibility of prenatal nutritional deprivation as a risk factor for schizophrenia. J Nerv Ment Dis, v. 184, n. 2, p. 71-85, Feb 1996.
BROWN, A. S. et al. Prenatal exposure to maternal infection and executive dysfunction in adult schizophrenia. Am J Psychiatry, v. 166, n. 6, p. 683-90, Jun 2009.
BROZOSKI, T. J. et al. Cognitive deficit caused by regional depletion of dopamine in prefrontal cortex of rhesus monkey. Science, v. 205, n. 4409, p. 929-32, Aug 31 1979.
BUKA, S.L. et al. Viruses, fetal hypoxia and subsequent schizophrenia. A direct test of infectious agents using prenatal sera. Schizophr Res. p. 36-38,
CARREY, W. P. Behavioral and morphological changes in carpenter ants harboring dicrocoeliid metacercariae. American Midland Naturalist, v. 82, p. 605-611, 1969.
CEZILLY, F. et al. Host-manipulation by parasites with complex life cycles: adaptive or not? Trends Parasitol, v. 26, n. 6, p. 311-7, Jun 2010.
CHOISY, M. et al. Evolution of trophic transmission in parasites: why add intermediate hosts? Am Nat, v. 162, n. 2, p. 172-81, Aug 2003.
pathological gambling, and substance abuse. Am J Med Genet, v. 88, n. 4, p. 358-68, Aug 20 1999.
CONKLIN, H. M. et al. Working memory functioning in schizophrenia patients and their first-degree relatives: cognitive functioning shedding light on etiology. Neuropsychologia, v. 43, n. 6, p. 930-42, 2005.
COOLS, R. Roles of dopamine in the motivacional and cognitive control behavior. The Neuroscientist, v.14, n. 4, p: 381-394, 2008.
DAMASIO, H. et al. The return of Phineas Gage: clues about the brain from the skull of a famous patient. Science, v. 264, n. 5162, p. 1102-5, May 20 1994.
DAWE, S.; LOXTON, N. L. The role of impulsivity in the development of substance use and eating disorders. Neuroscience and Biobehavioral Reviews. v. 28, p. 343-351, 2004.
DEARY, I. J. et al. The neuroscience of human intelligence differences. Nat Rev Neurosci, v. 11, n. 3, p. 201-11, Mar 2010.
DICKMAN, S. J. Functional and Dysfunctional Impulsivity: Personality and Cognitive Correlates. Journal of Personality and Social Psychology, v. 58, n. 1, p. 95-102, 1990.
DIGMAN, J. M. Personality structure: emergence of the five-factor model. Annu Rev Psychol, v. 41, p. 417-440, 1990.
DUBEY, J. P. Advances in the life cycle of Toxoplasma gondii. Int J Parasitol, v. 28, n. 7, p. 1019-24, Jul 1998.
DUBEY, J. P. The history of Toxoplasma gondii--the first 100 years. J Eukaryot Microbiol, v. 55, n. 6, p. 467-75, Nov-Dec 2008.
DUBEY, J. P. History of the discovery of the life cycle of Toxoplasma gondii. Int J Parasitol, v. 39, n. 8, p. 877-82, Jul 1 2009.
DUNCAN, J. et al. A neural basis for general intelligence. Science, v. 289, n. 5478, p. 457-60, Jul 21 2000.
GAALEN, M. M. V. et al. Critical involvement of dopaminergic neurotransmission in impulsive decision making. Soc of Biol Psychiatry, v. 60, p. 66-73, Aug 10 2006.
GASKELL, E. A. et al. A unique dual activity amino acid hydroxylase in Toxoplasma gondii. PLoS One, v. 4, n. 3, p. 1-10, 2009.
GOLDMAN-RAKIC, P. S. The prefrontal landscape: implications of functional architecture for understanding human mentation and the central executive. Philos Trans R Soc Lond B Biol Sci, v. 351, n. 1346, p. 1445-53, Oct 29 1996.
GUENTER, W. et al. Does Toxoplasma gondii infection affect cognitive function? A case control study. Folia Parasitologica, v. 59, p. 1-6, Jan 2012. GULINELLO, M. et al. Acquired infection with Toxoplasma gondii in adult mice results in sensorimotor deficits but normal cognitive behavior despite widespread brain pathology. Microbes and infection. v. 12, p: 528-537, Jan 25 2010.
HAIR, P.; HAMPSON, S. E. The role of impulsivity in predicting maladaptive behaviour among female students. Personality and Individual Differences, v. 40, n. 5, p. 943-952, 2006.
HAGGER, C. et al. Improvement in cognitive function and psychiatric symptoms in treatment- refractory schizophrenic patients receiving clozapine. Biological psychiatry, v, 34, p. 702-712, 1993.
HAVELAAR, A. H. et al. Disease burden of congenital toxoplasmosis. Clin Infect Dis, v. 44, n. 11, p. 1467-74, Jun 1 2007.
HENDRYKS, A. A. J. et al. The Five-Factor Personality Inventory: Cross-Cultural Generalizability across 13 Countries. European Journal of Personality, v. 17, n. 5, p. 347-373, 2003.
HOLLIMAN, R. E. Toxoplasmosis, behaviour and personality. J Infect, v. 35, n. 2, p. 105-10, Sep 1997.
HONEY, G. D. et al. Differences in frontal cortical activation by a working memory task after substituion of risperidone for typical antipsychotic drugs in patients with schizophrenia. PNAS, v. 96, n. 23, p. 13432-13437, Nov 9 1999. HOUSE, P. K. et al. Predator cat odors activate sexual arousal pathways in brains of Toxoplasma gondii infected rats. PLoS One, v. 6, n. 8, p. e23277, 2011.
HUGHES, R. N. The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci Biobehav Rev, v. 28, n. 5, p. 497-505, Sep 2004.
HUTCHISON, W. M. et al. Life cycle of toxoplasma gondii. Br Med J, v. 4, n. 5686, p. 806, Dec 27 1969.
HUTICHISON, W. M. et al. Chronic toxoplasma infections and familiarity- novelty discrimination in the mouse. Ann Trop Med Parasitol,v. 74, n.2, p. 145-150, Apr 1980.
HUTZ, C. S. et al. O desenvolvimento de marcadores para a avaliação da personalidade no modelo dos cinco grandes fatores. Psicologia: Reflexão e Crítica, v. 11, n. 2, p. 395-411, 1998.
INNES, E. A. A brief history and overview of Toxoplasma gondii. Zoonoses Public Health, v. 57, n. 1, p. 1-7, Feb 2010.
JONES-BRANDO, L. et al. Drugs used in the treatment of schizophrenia and bipolar disorder inhibit the replication of Toxoplasma gondii. Schizophr Res, v. 62, n. 3, p. 237-44, Aug 1 2003.
KANE, M. J. et al. Working memory capacity and fluid intelligence are strongly related constructs: comment on Ackerman, Beier, and Boyle (2005). Psychol Bull, v. 131, n. 1, p. 66-71; author reply 72-5, Jan 2005.
KANNAN, G. et al. Toxoplasma gondii strain-dependent effects on mouse behaviour. Folia Parasitol (Praha), v. 57, n. 2, p. 151-5, Jun 2010.
KNABLE, M. B. et al. Extrapyramidal side effects with risperidone and haloperidol at comparable D2 receptor ocupancy levels. Psychiatry Research:
Neuroimaging, v. 75, p. 91-101, Apr 1997.
KAPUR, S. et al. Relationship between dopamine D2 ocuppancy, clinical
response, and side effects: a double-blind Pet study of first- episode schizophrenia. Am J Psychiatry, v. 157, p. 514-520, 2000.
KAUFMAN, A. S. et al. How do educational attainment and gender relate to fluid intelligence, crystallized intelligence, and academic skills at ages 22-90 years? Arch Clin Neuropsychol, v. 24, n. 2, p. 153-63, Mar 2009.
KERTZMAN, S. et al. Computerized neuropsychological examination of impulsiveness: A selective revision. Isr Psychiatry Relat Sci, v. 43, n.2, p. 74-80, 2006.
KIRBY, L. G. et al. Contributions of serotonin in addiction vulnerability. Neuropharmacology, v. 61, n. 3, p. 421-32, Sep 2011.
LACROIX, R. et al. Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol, v. 3, n. 9, p. e298, Sep 2005.
LEE, M. A. et al. Effects of clozapine on cognitive function in schizophrenia. Journal of clinical psychiatry, v. 55, p. 8-14, 1994.
LEFEVRE, T. et al. Invasion of the body snatchers: the diversity and evolution of manipulative strategies in host-parasite interactions. Adv Parasitol, v. 68, p. 45-83, 2009.
LEWIS, D. A.; LEVITT, P. Schizophrenia as a disorder of neurodevelopment. Annu Rev Neurosci, v. 25, p. 409-32, 2002.
LINDOVA,J. et al. Higher Extraversion and lower conscientiounesness in humans infected with toxoplasma. European Journal of Personality. v. 26, p 285- 291, 2012.
LUFT, B. J.; REMINGTON, J. S. Toxoplasmic encephalitis in AIDS. Clin Infect Dis, v. 15, n. 2, p. 211-22, Aug 1992.
MCCRAE, R. R.; COSTA, P. T., JR. Personality trait structure as a human universal. Am Psychol, v. 52, n. 5, p. 509-16, May 1997.
MELZER, T. C. et al. Host Cell Preference of Toxoplasma gondii Cysts in Murine Brain: A Confocal Study. J Neuroparasitology, v. 1, 2010.
MELTZER, H. Y. et al. Classification of atypical antipsychotic drugs on the basis of dopamine D-1, D-2 and seretonin 2pki values. J Pharmacol Exp Ther, v. 251, p. 251- 246, 1989.
MCCLURE, S. M. et al. Separate Neural Systems Value Immediate and Delayed Monetary Rewards. Science. v. 306, p. 503-507, Jun 2004.
MILLER, E. K. The prefrontal cortex and cognitive control. Nat Rev Neurosci, v. 1, n. 1, p. 59-65, Oct 2000.
MIYAKE, A. et al. The unity and diversity of executive functions and their contributions to complex "Frontal Lobe" tasks: a latent variable analysis. Cogn Psychol, v. 41, n. 1, p. 49-100, Aug 2000.
MOELLER, F. G. et al. Psychiatric aspects of impulsivity. Am J Psychiatry, v. 158, n. 11, p. 1783-93, Nov 2001.
MORRISSETTE, N. S.; AJIOKA, J. W. The early years of Toxoplasma research: What's past is prologue. Int J Parasitol, v. 39, n. 8, p. 865-9, Jul 1 2009.
MURPHY, B. L. et al. Increased dopamine turnover in the prefrontal cortex impairs spatial working memory performance in rats and monkeys. Proc Natl Acad Sci U S A, v. 93, n. 3, p. 1325-9, Feb 6 1996.
NATH, A.; SINAI, A. P. Cerebral Toxoplasmosis. Curr Treat Options Neurol, v. 5, n. 1, p. 3-12, Jan 2003.
NEVES, D.P et al. Parasitologia humana. 11. ed. Atheneu. São Paulo. 2005. PAPPAS, G. et al. Toxoplasmosis snapshots: global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int J Parasitol, v. 39, n. 12, p. 1385-94, Oct 2009.
PAULESU, E. et al. The neural correlates of the verbal component of working memory. Nature, v. 362, n. 6418, p. 342-5, Mar 25 1993.
POROPAT, A. E. A meta-analysis of the five-factor model of personality and academic performance. Psychol Bull, v. 135, n. 2, p. 322-38, Mar 2009.
PRANDOVSZKY, E. et al. The neurotropic parasite Toxoplasma gondii increases dopamine metabolism. PLoS One, v. 6, n. 9, p. e23866, 2011.
ROCA, M. et al. Executive function and fluid intelligence after frontal lobe lesions. Brain, v. 133, n. Pt 1, p. 234-47, Jan 2010.
ROSENHECK, R. et al. A comparision of clozapine and haloperidol in hospitalized patients with refractory schizophrenia. New Engl J Med, v. 37, n. 12, p. 809-815, 1997.
SABIN, A. B.; FELDMAN, H. A. Dyes as Microchemical Indicators of a New Immunity Phenomenon Affecting a Protozoon Parasite (Toxoplasma). Science, v. 108, n. 2815, p. 660-3, Dec 10 1948.
SALGADO, J. F. Predicting job performance using FFM and non-FFM personality measures. Journal of Occupational and Organizational Psychology, v. 76, p. 323-346, 2001.
SAXON, A. S. et al. Intellectual deficits in children born with subclinical congenital toxoplasmosis: A preliminary report. The Journal of Pediatrics. v. 82, n. 5, p. 792-797, May 1973.