Fauna edáfica como indicadora de qualidade do solo em fragmentos florestais
e área sob cultivo do cafeeiro
Edaphic fauna as a soil quality indicator in forest fragments and coffee
growing area
DOI:10.34117/bjdv6n3-375
Recebimento dos originais: 10/02/2020 Aceitação para publicação: 24/03/2020
Danilo Cândido da Silva
Graduado em Tecnologia de Gestão Ambiental pelo Instituto Federal de Educação, Ciência e Tecnologia do Sul de Minas, Campus Inconfidentes-MG
Instituição: Instituto Federal de Educação, Ciência e Tecnologia do Sul de Minas, Campus Inconfidentes-MG
Endereço: Praça Tiradentes, 416, Bairro Centro, 37576-000, Inconfidentes – MG E-mail: danilocandido456@gmail.com
Jamil de Morais Pereira
Doutorado em Microbiologia Agrícola pela Escola Superior de Agricultura “Luiz de Queiroz” Tecnologia do Sul de Minas, Campus Inconfidentes-MG
Instituição: Instituto Federal de Educação, Ciência e Tecnologia do Sul de Minas, Campus Inconfidentes-MG
Endereço: Praça Tiradentes, 416, Bairro Centro, 37576-000, Inconfidentes – MG E-mail: jamilmpereira@gmail.com
Danielle Cristina Ortiz
Mestre em Ciência do Solo pela Universidade do Estado de Santa Catarina Instituição: Universidade do Estado de Santa Catarina
Endereço: Av. Luiz de Camões, 2090 – Bairro Conta Dinheiro, 88520-000, Lages – SC E-mail: ortiz.dco@gmail.com
Luís Carlos Iuñes de Oliveira Filho
Doutor em Manejo do solo pela Universidade do Estado de Santa Catarina Instituição: Universidade do Estado de Santa Catarina
Endereço: Rua Beloni Trombeta Zanin 680E – Bairro Santo Antônio, 89815-630, Chapecó – SC E-mail: iunes1981@gmail.com
Lilian Vilela Andrade Pinto
Doutora em Engenharia Florestal pela Universidade Federal de Lavras Tecnologia do Sul de Minas, Campus Inconfidentes-MG
Instituição: Instituto Federal de Educação, Ciência e Tecnologia do Sul de Minas, Campus Inconfidentes-MG
Endereço: Praça Tiradentes, 416, Bairro Centro, 37576-000, Inconfidentes – MG E-mail: lilian.vilela@ifsuldeminas.edu.br
Dilmar Baretta
Doutor em Solos e Nutrição de Plantas pela Escola Superior de Agricultura “Luiz de Queiroz” Instituição: Universidade do Estado de Santa Catarina
Endereço: Rua Beloni Trombeta Zanin 680E – Bairro Santo Antônio, 89815-630, Chapecó – SC E-mail: dilmar.baretta@udesc.br
RESUMO
A redução da cobertura de florestas naturais, principalmente pela concorrência com a atividade agropecuária, bem como o seu baixo nível de conservação, pode diminuir a diversidade dos organismos da fauna do solo, os quais atuam na decomposição da matéria orgânica e ciclagem de nutrientes, por exemplo, funções fundamentais para a manutenção da qualidade do solo. Desta forma, o objetivo deste estudo foi verificar o potencial da fauna edáfica como bioindicadora da qualidade do solo e/ou ambiental em Fragmento de Floresta Estacional Semidecidual secundária (MC), Floresta Estacional Semidecidual em regeneração (MI) e área sob cultivo do cafeeiro (CA). A coleta de invertebrados foi realizada por duas metodologias: monólitos de solo (TSBF) e armadilhas de queda. Para isso, em cada área, foram retirados 10 monólitos de solo (25 x 25 cm e 0-20 cm de profundidade) espaçados de 30 metros entre si. Nos mesmos pontos, foram instaladas as armadilhas de queda, as quais permaneceram na área por três dias. Foram também coletadas amostras de solo, para determinação de atributos químicos e umidade, além de serapilheira. Considerando a metodologia TSBF a maior densidade (m2) de organismos edáficos foi observado na área de MC, diferindo estatisticamente de CA. Já para as armadilhas de queda a maior abundância foi encontrada na área de MI, sem diferir de MC. Em relação aos índices ecológicos, pelo TSBF a riqueza e a diversidade máxima foram maiores na área de MC, diferindo significativamente das demais áreas. Em relação as armadilhas de queda não foram observadas diferenças em nenhum índice ecológico avaliado. Os grupos Formicidae e Isoptera foram os mais abundantes pela metodologia TSBF enquanto que Collembola e Formicidae pelas armadilhas. Os indivíduos do grupo Araneae obtiveram os maiores valores de abundância, densidade e frequência relativa nos dois métodos de coleta. Destacando-se como bons indicadores da qualidade, sendo encontrados em ambientes mais conservados. Através da Análise de Componentes Principais foi possível verificar nítida separação entre as áreas estudadas, e em ambas as metodologias de coleta a área de MC foi a que apresentou associação com o maior número de grupos de organismos da fauna edáfica. Sendo assim, as áreas de florestas (MC e MI) apresentaram maior qualidade biológica do solo em comparação a CA, destacando a importância da manutenção dessas áreas para a conservação da fauna edáfica e manutenção dos serviços ambientais por eles exercidos.
Palavras-chave: Invertebrados de solo; Atributos químicos; Mata Atlântica; monólitos de solo;
Armadilhas de queda.
ABSTRACT
The reduction of natural forest cover, mostly due to the competition with the agricultural activity, as well as its low conservation level, can diminish the diversity of the soil fauna organisms, which act in the decomposition of organic matter and nutrient cycling, for example, key functions for maintaining soil quality. Thus, the objective of this study was to verify the potential of edaphic fauna as a bioindicator of soil and / or environmental quality in Secondary Semideciduous Seasonal Forest (MC), Regenerating Semideciduous Seasonal Forest (MI) and coffee growing area (CA). Soil invertebrates were collected by the TSBF methodology, in which 10 soil monoliths (25 x 25 cm and 0-20 cm profundity) were retired from each area, 30 m apart. At the same collection points, the litter was sampled and 10 fall traps were installed, which remained buried in the ground for three days. Soil was collected for chemical attributes analysis and soil moisture, and litter. Considering the TSBF methodology the highest density (m2) of edaphic organisms was observed in the MC area, differing statistically from CA. For the fall traps the highest abundance was found in the MI area, without
differing from MC. Regarding the studied indexes, by TSBF the richness and maximum diversity were higher in the MC area, differing significantly from the other areas. For the fall traps no difference was observed in any evaluated ecological index. The Formicidae and Isoptera groups were more abundant by TSBF methodology and the fall traps were Collembola and again Formicidae. The individuals of the Araneae group obtained the highest values of abundance, density and relative frequency in both collection methods, demonstrating to be good bioindicators of soil quality, being found in more conserved environments. Through Principal Component Analysis it was possible to verify clear separation between the studied areas, and in both collection methodologies the MC area was the one that was associated with the largest number of edaphic fauna groups. Thus, the forest areas (MC and MI) showed higher soil biological quality compared to CA, highlighting the importance of maintaining these areas for the conservation of edaphic fauna and maintenance of environmental services performed by them.
Keywords: Soil invertebrates; Chemical attributes; Atlantic forest; soil monoliths; Fall Traps.
1 INTRODUÇÃO
As atividades humanas, mediante as práticas agrícola e florestais, interferem na abundância e diversidade da biota do solo, afetando os serviços do ecossistema a curto e a longo prazo (CROTTY et al., 2015), resultando em alterações na qualidade do solo, inclusive sua perda (TESFAHUNEGN, 2016). A qualidade do solo está relacionada com a sua capacidade de desenvolver suas funções, dentro dos limites de cada ecossistema, para sustentar a produtividade biológica e manter ou melhorar a qualidade da água, ar e saúde de plantas, animais e do Homem (DORAN & PARKIN, 1994).
À medida que se exploram os recursos dos ecossistemas de modo pouco racional, há a perda da qualidade ambiental e da biodiversidade. Admite-se que o modelo de implantação da lavoura cafeeira, no Sul de Minas Gerais, avançando sobre as áreas de fragmentos florestais conservados ou em recuperação, possam impor redução na atividade biológica do solo, principalmente sobre a abundância de organismos da fauna do solo, dependentes da cobertura vegetal natural como fonte de alimento e abrigo. Além disso, no Brasil, a grande maioria dos cultivos tradicionais de café adotam o modelo de monocultivo a pleno sol, como forma de aumentar os rendimentos da cultura, utilizando também espaçamentos cada vez mais adensados, com maiores populações de cafeeiros (COELHO et al., 2004). Consequentemente, esse tipo de modelo, muitas vezes têm efeito negativo sobre a qualidade do solo, perda de nutrientes minerais e redução da biota do solo (VASCONCELLOS et al., 2013).
Avaliar a qualidade do solo é uma tarefa bastante complexa, que exige o uso de um conjunto de indicadores, envolvendo as propriedades físicas, químicas e biológicas do solo. Diversos estudos, desenvolvidos em solos de ecossistemas florestais, mostram que o uso de indicadores biológicos de qualidade do solo são mais sensíveis, na caracterização das mudanças ambientais e/ou distúrbios antrópicos, comparados aos atributos físico-químicos possivelmente porque refletem uma situação
mais real (TÓTOLA & CHAER, 2002; SCHLOTER, DILLY & MUNCH, 2003; BARETTA et al., 2008).
Os sistemas de cultivos agrícolas e florestais, no geral, mudam a abundância e a diversidade da biota do solo, afetando os serviços do ecossistema a curto e a longo prazo (CROTTY et al., 2015). Estudo como o de Guimarães et al. (2016) avaliaram a influência de diferentes sistemas de produção de café, área de pousio e fragmento de vegetação nativa sobre a diversidade da fauna invertebrada epigea. A área de vegetação nativa mostrou-se mais diversificada e de maior complexidade vegetal, oferecendo melhores condições de desenvolvimento da fauna epigea, enquanto que, a área com cultivo de café exclusivo foi a que apresentou menor biodiversidade da fauna do solo. Corroborando com estes resultados, Santos et al. (2018) estudaram a macrofauna do solo em plantações de café orgânico comparado ao sistema convencional, e o cultivo orgânico apresentou maior diversidade de grupos taxonômicos.
Nesse sentido, o objetivo deste trabalho foi verificar o potencial da fauna edáfica como indicadora da qualidade do solo e/ou ambiental em fragmentos com diferentes estágios de conservação de Mata Estacional Semidecidual do bioma Mata Atlântica e área sob cultivo do cafeeiro no sul de Minas Gerais.
2 MATERIAL E MÉTODOS
2.1 CARACTERIZAÇÃO DA ÁREA DE ESTUDO
O estudo foi conduzido no mês de novembro de 2014, em uma propriedade agrícola localizada no município de Ouro Fino, Minas Gerais (22º 13’ 42,42” S e 46° 19’ 2,94” O, com altitude média de 1101 m). Para o estudo, foram selecionadas três áreas próximas entre si, sendo: mata conservada (MC), mata impactada (MI) e cultivo de cafeeiro (CA). Informações, características e históricos são apresentados na Tabela 1. O clima da região, segundo a classificação de Köppen, é o tropical úmido com duas estações definidas: seca (abril a setembro) e chuvosa (outubro a março), com precipitação pluviométrica média anual de 1600 mm e temperatura média anual de 18 °C.
Tabela 1. Características das áreas estudadas: mata conservada (MC), mata impactada (MI) e cultivo de cafeeiro (CA).
Área Tamanho (ha)
Tempo
de uso Histórico de manejo
MC 9,68 80 anos
Fragmento de floresta secundária Estacional Semidecidual pertencente ao Bioma Mata Atlântica. Presença de espécies, tais como Jacarandá (Dalbergia nigra),
Palmito-juçara (Euterpe edulis) e Canela-preta (Ocotea catharinensis). No
MI 14,52 50 anos
Fragmento de floresta em regeneração Estacional Semidecidual pertecente ao Bioma Mata Atlântica, com presença de espécies, tais como Cedro (Cedrela
fissilis), Embaúba (Cecropia angustifolia) e Caraguatá (Bromelia balansae). No
entorno há pastagem e outros fragmentos de mata. Há registros de entrada de gado.
CA 3,00 2 anos
O cultivo de cafeeiro, conduzido em sistema convencional com espaçamento de 2,40 x 0,60 m. Anteriormente, foi realizado o cultivo de batata no sistema convencional. Presença de plantas daninhas, tais como como a Braquiária (Brachiaria decumbens), Joá bravo (Solanum viarum) e Caruru (Amaranthus
retroflexus L.). No entorno já há fragmento de floresta.
2.2 CARACTERIZAÇÃO DO EXPERIMENTO
Para a avaliação da macrofauna edáfica foi utilizado o método TSBF (Tropical Soil Biology
and Fertility), descrito por Anderson & Ingram (1993). Em cada área estudada foram coletados 10
monólitos de solo de 25 × 25 × 20 cm, distantes 30 m ente si, com auxílio de um marcador de ferro. Antes da retirada do monólito de solo, a serapilheira foi amostrada nos mesmos pontos. As amostras de serapilheira e de monólito, foram acondicionados em sacos de polietileno e transportados para o laboratório. Em seguida, foi realizada a triagem manual dos indivíduos visíveis a olho nu, os quais foram armazenados em frascos com solução de álcool 75%. Posteriormente, os indivíduos foram identificados em grupos taxonômicos com o auxílio de um microscópio estereoscópico e literatura específica. Após a contagem dos indivíduos, determinou-se o número de indivíduos por m² (densidade), a riqueza (número de grupos taxonômicos), o índice de diversidade de Shannon-Weaver (H’), equabilidade de Pielou (J’) e diversidade máxima (Hmax).
Nos mesmos pontos de coleta dos monólitos, foram instaladas armadilhas de queda para avaliar a atividade de grupos da fauna do solo. Para isso, foram utilizados frascos de vidro de 6 × 12 cm (diâmetro × altura), contendo uma solução de 200 mL de água e detergente líquido neutro (2,5%). Após três dias (BARETTA et al., 2003) os frascos foram retirados e transportados para o laboratório, sendo o conteúdo dos frascos passados por peneira de 0,053 mm. Os indivíduos edáficos retidos na peneira foram recolhidos e armazenados em álcool 75%. Seguidamente, foram identificados e a caracterização da comunidade da fauna edáfica foi realizada com base no cálculo de abundância, riqueza de grupos taxonômicos, índice de H’, J’ e Hmax. Para os cálculos de riqueza, H’ e J’ foi utilizado o Software PAST 3.0, o Hmax foi calculado utilizando-se ln s, sendo s o número de grupos taxonômicos (riqueza).
2.3 DETERMINAÇÃO DOS ATRIBUTOS FÍSICO-QUÍMICOS
Para a determinação dos atributos físico-químicos retirou-se 10 amostras de solo, com o auxílio de um trado, na profundidade de 0-20 cm, nos mesmos pontos onde foram coletados os
monólitos de solo. As amostras de solo foram separadas, homogeneizadas e levadas para o laboratório. Foram determinados o pH, teor de fósforo (P), cálcio (Ca), magnésio (Mg), potássio (K), H+Al, Al e matéria orgânica do solo. A determinação dos atributos químicos ocorreu conforme metodologia descrita pela Embrapa (1999). A porcentagem de umidade do solo foi determinada por meio da pesagem de 10 g por amostra de solo, seguida de secagem em estufa a 105 °C por 48 h. A massa seca da serapilheira foi determinada a partir da secagem em estufa a 55 °C por 48h. Na Tabela 2, encontram-se os resultados das características químicas e físicas do solo.
Tabela 2. Características químicas e físicas do solo, na profundidade 0-20 cm, em mata
impactada (MI), mata conservada (MC) e Cultura de Café (CA). n=10
Variáveis MI(a) MC(a) CA(a)
pH (CaCl₂) 4,59 ± 034 4,37 ± 0,17 4,93 ± 031 P mg dm-3 1,76 ± 0,74 7,04 ± 2,94 8,98 ± 4,34 K mg dm-3 20,95 ± 5,19 33,96 ± 12,03 20,92 ± 8,01 Ca cmolc dm-3 0,73 ± 0,62 0,33 ± 0,28 2,59 ± 0,92 Mg cmolc dm-3 0,33 ± 0,22 0,18 ± 0,16 0,59 ± 0,23 H+AL cmolc dm-3 9,32 ± 1,29 7,35 ± 1,28 5,99 ± 1,98 Al cmolc dm-3 1,41 ± 057 0,85 ± 0,21 0,51 ± 0,37 CTC cmolc dm-3 10,45 ± 0,97 7,94 ± 1,17 9,24 ± 1,18 V % 10,81 ± 8,03 7,6 ± 6,14 36,26 ± 14,88 C-org g Kg-1 13,42 ± 0,97 14,44 ± 2,01 14,23 ± 1,41 Zn mg dm-3 0,25 ± 0,23 0,33 ± 0,25 0,6 ± 0,30 Fe mg Kg-1 78,3 ± 37,65 101,58 ± 33,51 58,69 ± 28,61 Mn mg dm-3 47,99 ± 26,17 19,78 ± 17,98 28,46 ± 9,24 B mg dm-3 0,13 ± 0,07 0,13 ± 0,04 0,11 ± 0,06 Umidade do solo % 26,37 ± 3,69 24,78 ± 2,85 31,08 ± 4,12 MSS(b) Kg m-2 0,81 ± 0,23 0,98 ± 0,28 —(c)
(a)média e desvio padrão, (c)massa seca da serapilheira, (c)ausência de serapilheira.
2.4 ANÁLISES ESTATÍSTICA
Os dados de densidade (indivíduos por metro quadrado, ind m-2), abundância, riqueza taxonômica, diversidade de H’, J’ e Hmax foram submetidos à análise de variância (ANOVA) de efeitos principais utilizando o teste post-hoc de Fisher LSD (p < 0,05) com auxílio do programa STATISTICA versão 7.
Ainda, com os resultados de densidade (TSBF) e de abundância total (armadilhas de queda) utilizou-se o teste de correlação de Pearson com as variáveis físicas: umidade do solo (Umid); massa
seca da serapilheira (MSS) e químicas do solo (pH, P, K, Ca, Mg, H+Al, Al, CTC, V%, C-org, Zn, Fe, Mn, Cu e B).
Os dados de densidade e abundância foram submetidos à análise estatística multivariada. Inicialmente a uma Análise de correspondência destendenciada (DCA). Considerando o resultado do comprimento do gradiente, com resposta linear (< 3), optou-se pela realização da Análise de Componentes Principais (ACP), para cada método de amostragem. A Análise de Redundância foi realizada para identificar e retirar as variáveis ambientais (atributos químicos e físico do solo) colineares, além de selecionar um conjunto mínimo de variáveis, com base em permutações por teste de Monte-Carlo. Somente as variáveis ambientais selecionadas nas RDAs foram posteriormente utilizadas na ACP como explicativas passivas para as mudanças observadas na fauna nas áreas estudadas de araucária e café. Para todas as análises estatísticas multivariadas utilizou-se o programa estatístico CANOCO versão 4.5.
3 RESULTADOS E DISCUSSÃO
3.1 DENSIDADE E FREQUÊNCIA RELATIVA DE INDIVÍDUOS DA FAUNA EDÁFICA Houve diferença significativa (p < 0,05) para densidade de indivíduos (ind. m-2) entre as áreas
estudadas pelo método de coleta TSBF (Figura 1A). A MC apresentou o maior valor de densidade, diferindo estatisticamente de CA. Este resultado, corrobora com os estudos realizados por Baretta et al. (2010) e Pereira et al. (2017) que obtiveram maior densidade de indivíduos em florestas nativas, comparados a reflorestamentos e áreas impactadas. Ecossistemas com maior diversidade de espécies vegetais, tais como encontrado em florestas nativas proporcionam um microclima mais favorável, com maior quantidade e qualidade de fontes de alimentos e abrigo para a fauna edáfica, possibilitando condições para que esses organismos possam sobreviver e se reproduzir (SZINWELSKI et al., 2012; PEREIRA et al., 2013).
Para a abundância total de indivíduos da fauna edáfica pelo método de armadilhas de queda, houve também diferença significativa (Figura 1B), em que, a MI apresentou a maior abundância de organismos, porém sem diferir da MC. A área de CA apresentou a menor abundância, sem diferir da MC. O fato de MI apresentar uma maior abundância e atividade de indivíduos da fauna, pode indicar que essa mata está evoluindo no seu estágio sucessional, passando de um sistema mais simples para mais complexo, aumentando sua diversidade de espécies e melhorando as condições ambientais para a presença de organismos edáficos.
Figura 1. (A) Densidade de indivíduos total ind m-² da fauna edáfica amostradas pela metodologia TSBF; (B) Abundância total da fauna edáfica amostradas por armadilhas de queda; Mata impactada (MI), mata conservada (MC) e cultivo do cafeeiro (CA). Médias seguidas de mesma letra não diferem entre si, pelo teste de Fisher LSD a 5% de probabilidade.
Estudos afirmam que a permanência da fauna edáfica, em determinado ambiente, é muito influenciada pela conversão de áreas nativas, seja agrícola ou florestal (ARAÚJO et al., 2010; CROTTY et al., 2015; POMPEO et al., 2016). Desta forma, a menor abundância/densidade de organismos da fauna do solo na área de CA já era esperada, visto que é um sistema de monocultivo convencional. No estudo de Santos et al. (2018) ao avaliarem a macrofauna do solo em cultivo de café orgânico e convencional, encontraram, embora sem diferença estatística, 50% mais indivíduos na área de cultivo orgânico do que no cultivo convencional.
Dentre os grupos edáficos encontrados (Tabela 3) e a frequência relativa (Figura 2A) pelo método TSBF, observa-se que os insetos sociais (Formicidae e Isoptera) foram os que apresentaram maior densidade nas três áreas estudadas, principalmente na área de MC. No estudo de Baretta et al. (2010) os insetos sociais também foram os mais abundantes em florestas conservadas de araucária, quando comparadas a área com o reflorestamento da espécie. Em suma, as formigas (Formicidae) juntamente com os cupins (Isoptera) são os grupos que mais dominam as florestas tropicais em abundância, biomassa e também em relação ao grande número de funções ecológicas por elas exercidas (KORASAKI, MORAIS & BRAGA, 2013).
ab a b 0 500 1000 1500 2000 2500 MI MC CA D e n si d ad e ( in d /m 2) a ab b 0 20 40 60 80 100 120 MI MC CA A b u n d ân c ia
Tabela 3. Densidade (ind m-2) e abundância total dos principais grupos taxonômicos da fauna
edáfica amostrados pelo método TSBF e armadilhas de queda, nas áreas de mata impactada (MI), mata conservada (MC) e cultivo do cafeeiro (CA).
Método TSBF Grupos Taxonômicos MI MC CA Acarina 0 96 (±20,24) 32 (±5,76) Araneae 336 (±30,59) 336 (±21,93) 128 (±10,24) Blattodea 192 (±34,40) 96 (±11,19) 16 (±2,88) Chilolopoda 288 (±24,79) 400 (±27,46) 64 (±10,24) Coleoptera 208 (±25,07) 224 (±25,24) 432 (±24,96) Coleoptera - Staphylinidae 352 (±38,80) 272 (±20,03) 656 (±43,84) Collembola 32 (±6,75) 192 (±23,61) 272 (±37,12) Dermaptera 128 (±21,06) 64 (±11,19) 0 Diplopoda 320 (±36,95) 624 (±67,23) 16 (±2,88) Diplura 96 (±13,49) 80 (±15,55) 0 Formicidae 4256 (±455,88) 7136 (±999,29) 992 (±109,44) Hemiptera 320 (±41,31) 208 (±18,55) 192 (±17,92) Isoptera 1200 (±314,28) 3008 (±619,07) 960 (±166,4) Larva de Coleoptera 240 (±33,96) 864 (±75,12) 544 (±49,92) Larva de Diptera. 16 (±5,06) 208 (±28,27) 80 (±12,8) Larva de Lepidoptera 0 80 (±13,60) 80 (±12,8) Oligochaeta 192 (±29,02) 320 (±29,21) 528 (±30,4) Orthoptera 32 (±6,75) 32 (±6,75) 0 Outros* 640 (±86,48) 1728 (±121,26) 288 (±19,84) Armadilhas de queda Grupos Taxonômicos MI MC CA Acarina 4 (±0,52) 5 (±0,71) 0 Araneae 33 (±2,45) 44 (±3,66) 11 (±1,20) Blattodea 5 (±0,97) 11 (±1,60) 0 Chilopoda 0 1 (±0,32) 0 Coleoptera 51(±3,41) 21 (±2,18) 13 (±1,42) Coleoptera - Staphylinidae 97 (±10,40) 36 (±2,32) 48 (±4,13) Collembola 436 (±21,09) 341 (±16,80) 261 (±35,54) Diplopoda 3 (±0,67) 3 (±0,95) 0
Diptera 240 (9,27) 81 (±4,23) 39 (±1,29) Formicidea 62 (±2,49) 161 (±7,14) 240 (±29,34) Grylloblattodea 5 (±0,71) 12 (±1,69) 31(±3,28) Hemiptera 7 (±1,15) 1 (±0,32) 17 (±1,16) Isoptera 2 (±0,63) 0 10 (±1,33) Orthoptera 68 (±3,79) 35 (±2,17) 3 (±0,95) Outros* 10 (±1,34) 5 (±0,32) 54 (±12,51)
*Outros = Somatório de outros grupos
Para as armadilhas de queda foram Collembola e novamente Formicidae que apresentaram a maior abundância (Tabela 3) e frequência relativa (Figura 2B) de indivíduos. Estes grupos também foram observados como mais abundantes nos estudos de Antoniolli et al. (2006) e Brancher & Roza-Gomes (2012), estudando ambientes com sistemas de cultivo e mata nativa. Os maiores valores de abundância de Collembola foram encontrados nos fragmentos de mata MI e MC, ocorrendo o contrário para a abundância de Formicidae, que apresentou valores mais elevados na área de CA (Tabela 3). Resultado semelhante foi encontrado por De Lima et al. (2017), estudando a fauna edáfica como bioindicadora da qualidade do solo em áreas com e sem a adição de topsoil em plantios florestais para a recuperação de áreas de extração de cascalho, considerando a mata nativa da Caatinga como referencial.
Figura 2. Frequência relativa dos principais grupos da fauna edáfica (A) coletados com a
metodologia TSBF (B) coletados com armadilhas de queda. (n= 10). Ara= Araneae; Chilo= Chilopoda; Diplo= Diplopoda; Form= Formicidae; Isopt= Isoptera; Oligo= Oligochaeta; Coll=
Collembola; Griloblat= Grylobllatodea; Ortho= Orthoptera.
Os indivíduos do grupo Araneae obtiveram os maiores valores de abundância, densidade e frequência relativa nos dois métodos de coleta (métodos armadilhas de queda e TSBF). Em ambos os métodos, foram encontrados no solo de ambientes com maior densidade de cobertura vegetal MI e
MC comparados à área de cultivo do cafeeiro (CA) (Figura 2 e Tabela 3). As famílias de aranhas são afetadas pelo tipo de uso e manejo do solo, sendo mais abundantes em locais com menor intervenção humana, como relata o estudo de Rosa et al. (2019). Esses autores encontraram maior abundância de famílias de aranhas em áreas de floresta nativa, comparando-as com áreas de reflorestamento de eucalipto, integração lavoura-pecuária, plantio direto, pastagem e plantio convencional.
O grupo Chilopoda (Figura 2A) apresentou maior abundância e frequência relativa pelo método TSBF, em que, no ambiente mais conservado MC (90%) foi encontrado os valores mais elevados. Maior densidade e diversidade de Chilopoda foram encontradas em solos sob florestas de araucárias nativas, comparados a reflorestamentos e áreas com araucária que sofreram alguma intervenção antrópica, indicando uma relação entre ambientes florestais mais conservados e presença de grupos predadores (Chilopoda) à semelhança do que acontece com o grupo Araneae (BARETTA, 2007).
Os Diplopodas coletados pela metodologia TSBF (Figura 2A) apresentaram maior abundância e frequência relativa (90%) na área de mata conservada (MC) em comparação com a MI (60%) e CA (10%) (Figura 2). Esta diferença indica que nos ambientes mais preservados, a cobertura vegetal proporciona maior acúmulo de serapilheira em decomposição, aumentando a população de fungos, que servem de alimento para o grupo Diplopoda (COSTA NETO, 2007).
Ainda pelo método TSBF, a densidade e a frequência relativa de Oligochaeta foram diferentes nas três áreas (Tabela 3 e Figura 2A), em que, a área de CA teve uma maior abundância de indivíduos em relação às demais. Entretanto, a área mais conservada (MC) foi a que apresentou uma maior frequência relativa (100%) em comparação com a CA (90%) e MI (60%). Na MC foi identificada duas espécies nativas de minhocas, Urubenus brasiliensis e Fimoscolex sp., e no CA ocorreu predomínio da espécie Pontoscolex corethrurus (dados não apresentados). Estes resultados mostram que a espécie P. corethrurus, considerada muito agressiva e invasora, ainda se encontra restrita a área cultivada, não presente nos fragmentos florestais mais conservados.
Esse resultado é relevante porque indica a necessidade de o isolamento dos fragmentos de mata, visando a preservação das espécies de minhocas nativas ali presentes. Fernandes, Uehara-Prado & Brown (2012) encontraram espécies de minhocas exóticas e a peregrina P. corethrurus em maior abundância em ambientes mais impactados, enquanto as espécies de minhocas nativas nos ecossistemas mais preservados. Nesse sentido, destaca-se a importância do uso das minhocas como bioindicadoras da qualidade do solo e mudanças ambientais (BARETTA, 2007; BROWN, MASCHIO & FROUFE, 2009).
A ordem Orthoptera, apresentou valores de abundância (Tabela 3) e frequência relativa (Figura 2B) maiores nas áreas MI (100%) e MC (100%) em comparação com a CA (10%), pelo
método de armadilhas de queda. Rezende et. al. (2007) encontraram maior abundância de grilos, amostrados por armadilhas, após período de chuva e em área de mata com estádio de sucessão ecológica mais avançada, demonstrando o efeito positivo da umidade na sobrevivência dos grilos. A ordem Grylloblattodea coletada pelas armadilhas de queda foi mais abundante (Tabela 3) e frequente (Figura 2B) no solo sob cultivo do cafeeiro (CA). Esta ordem ainda é muito pouco estudada no Brasil, principalmente relacionando-a como bioindicadora de qualidade do solo (BARETTA et al., 2011), necessitando de mais estudos.
3.2 RIQUEZA E ÍNDICES DE DIVERSIDADE
Pela metodologia de coleta TSBF (Tabela 4) foi encontrado diferença significativa para a riqueza de espécies e Hmax. A área de MC apresentou os valores mais elevados de riqueza e Hmax, diferindo de MI e CA, os quais não diferiram entre si. Para os demais índices de diversidade não foi observado diferença significativa. Já para o método de armadilhas de queda (Tabela 4) não foi observado diferença estatística para a riqueza e índices de diversidade utilizados.
Tabela 4. Valores de riqueza, diversidade de Shannon-Waver (H’), equabilidade de Pielou (J’) e diversidade máxima
(Hmax), nas áreas estudadas de mata impactada (MI), mata conservada (MC) e cultivo de cafeeiro (CA), pelos métodos de coleta TSBF e armadilhas de queda.
TSBF
Atributos da fauna edáfica MI MC CA
Riqueza 9,50b 12,20a 7,10b Shannon (H') 1,49ns 1,63ns 1,45ns Pielou (J) 0,52ns 0,48ns 0,68ns Hmax 2,23b 2,48a 1,87b Armadilhas de queda Riqueza 9,00ns 8,70ns 7,50ns Shannon (H') 1,58ns 1,58ns 1,30ns Pielou (J) 0,57ns 0,58ns 0,52ns Hmax 2,17ns 2,15ns 1,99ns
ns: não significativo. Médias seguidas de mesma letra na linha não diferem entre si, pelo teste de Fisher LSD a 5% de probabilidade.
No estudo de Rosa et al. (2015) no Planalto-Sul Catarinense, avaliando a macrofauna edáfica como bioindicadora da qualidade do solo, encontraram maiores valores de riqueza na floresta nativa quando comparada a sistemas de cultivos agrícola e área de reflorestamento com eucalipto. Os autores destacaram que a intensidade de uso do solo é uma das fontes de variação na distribuição dos grupos da macrofauna edáfica. Assim como, estudos de Baretta et al. (2010) e Pereira et al. (2017), obtiveram resultados semelhantes, pois ao comparar áreas de florestas nativas com áreas de reflorestamento ou
que sofreram algum tipo de intervenção antrópica, encontraram maior riqueza de grupos de invertebrados de solo em áreas mais conservadas.
3.3 ANÁLISE DE CORRELAÇÃO DE PEARSON
Para a metodologia TSBF, houve correlação significativa entre a densidade de indivíduos e atributos químicos do solo (Tabela 4). A densidade de indivíduos na mata impactada (MI) apresentou valores de correlação negativa: com fósforo, em que foram encontrados os menores valores para fósforo no solo (Tabela 4) e alumínio, onde a concentração de alumínio apresentou valores mais elevados. Positiva: com os atributos cálcio, saturação por base. Na mata conservada (MC) a única correlação significativa foi com carbono orgânico, onde os maiores valores foram encontrados em solo sob mata conservada (MC) (Tabela 2). Já na área sob cultivo do cafeeiro (CA), foram encontradas correlações positivas com: magnésio e capacidade de troca de cátions (Tabela 4). Esses resultados demonstram que há influência de diferentes atributos químicos do solo sobre a abundância de grupos da fauna nas diferentes áreas. Lourente et al. (2007), encontraram correlações positivas (Ca, Al e Carbono orgânico) e negativas (P e K) no solo sob diferentes sistemas de cultivo e grupos de invertebrados do solo.
Tabela 4. Coeficiente de correlação de Pearson (r) entra os atributos químicos do solo e
densidade de indivíduos pelo método TSBF nas áreas mata impactada (MI), mata conservada (MC) e cultivo do cafeeiro (CA).
TSBF Correlação MI MC CA P 0,821 -0,17 0,38 Ca 0,50 0,23 0,39 Mg 0,43 0,07 0,46 Al -0,45 -0,39 -0,16 CTC 0,01 -0,31 0,47 V (%) 0,49 0,19 0,20 C-org (g Kg-1) -0,06 -0,52 -0,34 Armadilhas de queda Correlação MI MC CA P (mg dm-3) 0,12 0,53 0,17 Ca (mg dm-3) -0,24 0,55 0,17 V (%) -0,32 0,47 0,18 C-org (g Kg-1) -0,37 -0,07 0,53 Umd. (%) 0,771 -0,31 0,12 M.S.S (Kg m-2) 0,14 0,62 -2
1Valores em negrito apresentam correlação significativa, pelo teste de t com 5% de probabilidade, 2Massa da serapilheira nula.
Houve correlação significativa também utilizando as armadilhas de queda, entre o número de indivíduos e atributos químicos e físicos do solo (Tabela 4). O número de indivíduos por armadilha na mata impactada (MI) apresentou valores de correlação positiva: com umidade do solo. Na mata conservada (MC) apresentou correlação positiva: com o Fósforo, saturação por base e massa seca da serapilheira. Já na área sob cultivo do cafeeiro (CA), ocorreu correlação positiva com o carbono orgânico. Esses resultados demonstram que há relação significativa com a atividade da fauna edáfica com os atributos químicos e físicos do solo.
3.4 ANÁLISES DE COMPONENTES PRINCIPAIS (ACP)
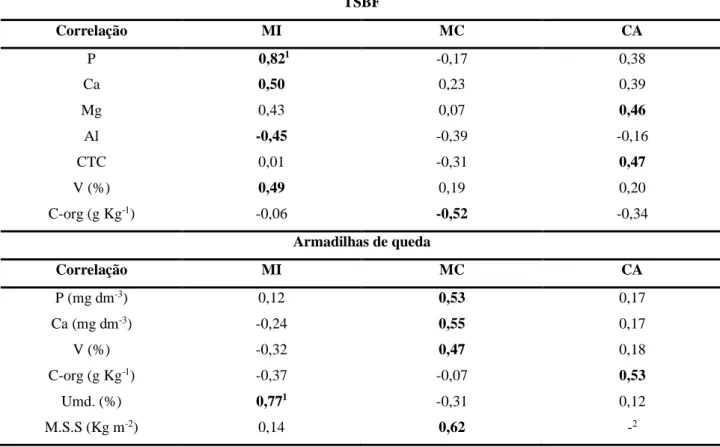
Na análise de componentes principais (ACP), para o método de coleta TSBF (Figura 3A), a variabilidade total dos dados da abundância foi explicada em 21,0% pela componente principal 1 (CP1) e 13,9% pela componente principal 2 (CP2), totalizando 34,9%. Observa-se através da ACP uma nítida separação das áreas, e também notavelmente a área de MC ficou relacionada ao maior número de grupos da fauna edáfica.
As variáveis ambientais utilizadas na ACP auxiliam na explicação da distribuição da fauna edáfica em cada área, por meio da associação que demonstram após serem projetadas a posteriori na análise. Desta forma, pelo método TSBF, H+Al e Mn contribuem para explicar a abundância da fauna edáfica na MI. Enquanto a MSS e K explicam os organismos associados à MC. Na área CA, a variável ambiental explicativa que mais contribuiu foi a umidade (Figura 3A).
A área de MC, associou o maior número de grupos de organismos edáficos (Ara, Derm, Form, Chi, Ter, Larvas, Diplo, Thy, Hem, Outros, Isop e Dipt). Para a área de MI, os organismos relacionados foram Blat, Hym e Dip. Já a área de CA se associou aos grupos Col e Oligo. No estudo de Pompeo et al. (2016) também encontraram maior número de grupos de organismos edáficos em área de floresta nativa, comparando com área de reflorestamento de pinus e campo nativo melhorado. Como destacado pelos autores, tais resultados podem estar relacionados a intensidade de uso do solo, que afeta os organismos edáficos e ao microclima encontrado em áreas de floresta nativa, o qual permite menores variações de temperatura, proporcionando ambiente mais favorável ao estabelecimento de vários grupos edáficos, assim como, a diversidade florística, que resulta em uma serapilheira mais diversa, com variedade de alimento e substrato para a fauna.
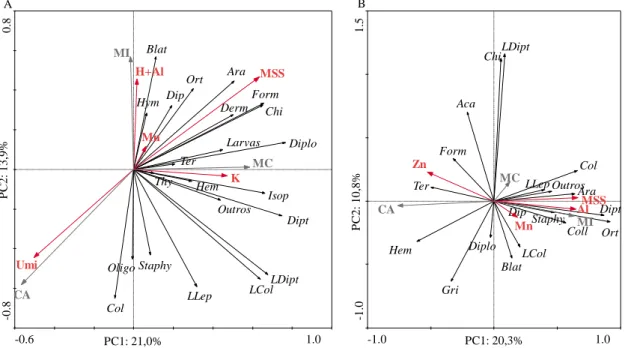
Figura 2. Relação entre a componente principais 1 (CP1) e componente principal 2 (CP2) da Análise de Componentes
Principais (ACP), dos grupos da fauna edáfica (setas pretas) e sua relação com as áreas de mata conservada (MC), mata impactada (MI) e cultivo de café (CA) (setas cinzas) e as variáveis ambientais (setas vermelhas) utilizadas como explicativas, no método de coleta TSBF (A) e metodologia armadilhas de queda (B). Abreviações - Aca: Acari; Ara:
Araneae; Blat: Blattodea; Chi: Chilopoda; Col: Coleoptera; Staphy: Coleoptera - Staphylinidae; Derm: Dermaptera; Diplo: Diplopoda; Dip: Diplura; Dipt: Diptera; Form: Formicidae; Gri: Gryllidae - Grilos; Hem: Hemiptera; Hym: Hymenoptera; Iso: Isopoda; LCol: Larva de Coleoptera; LDipt: Larva de Diptera; LLep: Larva de Lepidoptera; Larvas:
Outras; Oligo: Oligochaeta; Ort: Orthoptera; Outros; Ter: Termitoidae; Thy: Thysanoptera; Umi: Umidade; H+Al: Acidez potencial; MSS: Massa seca da serapilheira; Mn: Manganês; K: Potássio; Zn: Zinco; Al: Alumínio.
Além disso, a variável ambiental MSS e K ajudam a explicar a presença dos grupos taxonômicos encontrados na área de MC. Áreas de florestas nativas, devido a diversidade da cobertura vegetal possuem uma serapilheira rica, composta por folhas, ramos, flores, sementes e frutos, que servem de alimento e abrigo para a fauna edáfica. Estudos como de Maestri et al. (2013) ao compararem a diversidade dos organismos do solo em uma área de mata nativa e plantio de eucalipto, encontraram na área com mata uma maior diversidade, e atribuiu o resultado a maior heterogeneidade e complexidade estrutural do ambiente. Os autores relacionaram a variação na composição dos organismos edáficos devido a massa da serapilheira, visto que na mata nativa foi maior do nas áreas de eucalipto.
A MSS (Tabela 2) foi maior na área de MC e ausente na área de CA. No estudo de Silva et al. (2012) foi realizada a avaliação da fauna edáfica em diferentes sistemas de uso da terra, tais como: mata nativa, café convencional, manejo agroecológico com café e sistema agroflorestal com café. Verificou-se no café convencional a presença de uma serapilheira podre, contendo apenas algumas folhas de café e alguns brotos, enquanto que nos demais sistemas de uso, a cobertura vegetal foi
-0.6 1.0 -0.8 0.8 Ara Blat Chi Col Staphy Derm Diplo Dip Dipt Form Hem Hym Isop LColLDipt LLep Larvas Oligo Ort Outros Ter Thy K H+Al Mn Umi MSS MI MC CA PC1: 21,0% P C2: 13, 9% -1.0 1.0 -1.0 1.5 Aca Ara Blat Chi Col Staphy Coll Diplo Dip Dipt Form Gri Hem LCol LDipt LLep Ort Outros Ter Al Zn Mn MSS MI MC CA PC1: 20,3% P C2: 10 ,8% A B
diversa e espessa. Nesse sentido, uma serapilheira de maior qualidade e quantidade de massa fornece aos organismos edáficos um ambiente mais favorável à sua sobrevivência e permanência. Além disso, a MSS contribuiu para a maior disponibilidade de K na área MC, porque a serapilheira mais espessa e de qualidade, auxilia para o aumento da MO no solo, que juntamente com a atividade dos microrganismos e da fauna edáfica possibilitam maiores concentrações deste macronutriente no solo (RODRIGUES et al., 2010). E, consequentemente, o K influencia positivamente o número de organismos edáficos (CANTO, 1996).
Em relação a área CA, observa-se que ela ficou associada a um menor número de organismos edáficos (apenas Col e Oligo), o que também foi observado no estudo de Santos et al. (2018), os quais avaliaram a fauna edáfica em um sistema de cultivo de café orgânico comparado ao sistema de cultivo de café convencional. Adicionalmente, estes resultados corroboram os nossos porque os autores verificaram na ACP maior número de grupos taxonômicos associados a produção orgânica de café. Além disso, no mesmo estudo foi encontrada maior diversidade de espécies de minhocas no verão na área de cultivo convencional, porém, tanto no cultivo orgânico quanto convencional, foram identificadas apenas espécies de minhocas peregrinas ou exóticas. Estudos como o de Grosso, Jorge & Brown (2006) destacam que no manejo agrícola convencional, com intensidade de uso do solo, a comunidade de minhocas é composta majoritariamente por espécies exóticas devido à sua maior adaptabilidade. O fato das espécies de minhocas exóticas predominarem, representa um risco para a sobrevivência das espécies nativas (SANTOS et al., 2018).
Na área de CA, a umidade foi a variável ambiental explicativa que pode ter contribuído para a presença dos grupos Col e Oli. A fauna edáfica é influenciada pela umidade do solo, pois de modo geral preferem solos úmidos. Na área de CA, mesmo não sendo esperado, o teor de umidade do solo foi maior que a na MC e MI (Tabela 2). Entretanto, ressalta-se que a diversidade de organismos edáficos depende de inúmeros fatores, entre os quais estão o tipo de manejo, a disponibilidade de fontes de alimento, teor de matéria orgânica e umidade do solo (POMPEO et al., 2016).
Seguindo para a metodologia de armadilhas de queda (Figura 3B), a variabilidade total dos dados da abundância foi explicada em 20,3% pela CP1 e 10,8% pela CP2, totalizando 30,10%. Por meio da ACP, outra vez, foi possível observar uma nítida separação entre as áreas, e uma proximidade entre a MI e MC, que foram as que abrangeram o maior número de grupos de organismos edáficos. Os grupos edáficos Col, LLep, Outros, Ara, Chi e LDipt ficaram relacionados com a MC. Já para a área de MI os grupos relacionados foram Dipt, Dip, Staphy, Coll, Ort e LCol. Enquanto para CA Hem, Ter e Form foram os grupos relacionados. Os grupos edáficos Aca, Gri, e Diplo não se relacionaram a nenhuma área. Os atributos Al e Mn explicaram a presença de grupos edáficos na área
de MI. Enquanto a MSS melhor contribuiu para explicar a abundância dos grupos da fauna edáfica associados à MC e o Zn aos grupos associados à CA (Figura 3B).
Novamente as áreas de MC e MI ficaram associadas a maioria dos grupos dos organismos edáficos. Isso, pode indicar que as características do solo encontradas nesses ambientes favoreceram para o estabelecimento da fauna edáfica, e que as condições encontradas na área de CA limitam o desenvolvimento de grupos restritos. A variável MSS associada à área de MC é essencial para criar um habitat ideal para a presença de grupos da fauna edáfica nesse ambiente.
Porém, há grupos da fauna edáfica que são mais ou menos restritivos a determinadas condições ambientais encontradas. O grupo Formicidae, relacionado a área de CA, está sendo comumente utilizado como indicador biológico da qualidade do solo (ROCHA et al., 2015; OLIVEIRA et al., 2016). Isto, porque é um grupo considerado de alta riqueza de espécies, por possuírem táxons especializados, pela grande sensibilidade em perceberam alterações ambientais e serem relativamente fáceis de coletar (ALONSO & AGOSTI, 2000; SOUZA et al., 2018).
No entanto, enquanto há espécies de formigas sendo prejudicadas por causa da degradação ambiental, outras podem ter sua abundância aumentada, por possuírem um elevado potencial de adaptabilidade (ROCHA et al., 2015). Desta forma, é possível pressupor que a espécie de, que a espécie de Formicidae associada na área de CA pode ser mais adaptada a condições limitadas de recursos, os quais seriam mais facilmente encontrados em áreas de floresta nativa, que possuem maior complexidade estrutural, alimento e abrigo, entre outros.
4 CONCLUSÃO
Os ambientes que apresentaram maior abundância e riqueza de grupos taxonômicos foram as áreas MC e MI, respectivamente, ressaltando a importância dos ecossistemas florestais para a conservação de grupos da fauna edáfica e manutenção da qualidade biológica do solo.
Dentre os organismos edáficos coletados durante a avaliação, os indivíduos do grupo Araneae obtiveram os maiores valores de abundância, densidade e frequência relativa nos dois métodos de coleta (método de armadilhas de queda e TSBF). E, em ambos, o grupo Araneae foi encontrado no solo de ambientes com maior densidade de cobertura vegetal (MI e MC) comparados à área de cultivo do cafeeiro (CA). Isso indica que esse grupo é afetado pelo tipo de uso e manejo do solo, sendo mais abundantes em locais mais conservados, onde haja menor intervenção antrópica.
A análise de ACP separou nitidamente as áreas, independentemente do método de coleta. Nesta, a área MC ficou relacionada ao maior número de grupos da fauna edáfica. As variáveis ambientais que explicaram a presença de grupos da fauna edáfica pelo método TSBF foram H+Al, Mn, MSS, K e umidade, enquanto pelo método de armadilhas de queda foram Al, Mn, MSS e Zn.
Assim sendo, a fauna edáfica demonstrou ser influenciada pelas práticas realizadas na lavoura cafeeira, confirmando que os organismos edáficos são bons bioindicadores da qualidade do solo, destacando principalmente o grupo Araneae.
AGRADECIMENTOS
Os autores agradecem ao Instituto Federal de Educação, Ciência e Tecnologia do Sul de Minas, Campus Inconfidentes-MG pelo suporte financeiro para publicação. L. C. I. O. F. é bolsista do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) – Brasil (155778/2018-8). D. B. agradece ao CNPq pela Bolsa de Produtividade em Pesquisa (305939/2018-1).
REFERÊNCIAS
ALONSO, L. E.; AGOSTI, D. Biodiversity studies, monitoring, and ants: an overview. In: AGOSTI, D.; MAJER, J. D.; ALONSO, L. E.; SCHULTZ, T. R. Ants standard methods for measuring and
monitoring biodiversity, p. 1-8, 2000.
ANDERSON, J. M.; INGRAM, J. S. I. Tropical soil biological and fertility: A Handbook of methods. 2. ed. Wallingford: C.A.B. International, 1993.
ANTONIOLLI, Z. I.; CONCEIÇÃO, P. C.; BÖCK, V.; PORT, O.; SILVA, D. M.; SILVA, R. F. Método alternativo para estudar a fauna do solo. Ciência Florestal, v. 16, n.4, p. 407-417. 2006. ARAÚJO, C. C. NOMELINI, Q. S. S.; PEREIRA, J. M.; LIPORACCI, H. S. N.; KATAGUIRI, V. S. Comparação da abundância de invertebrados de solo por meio da estimação intervalar encontrados em diferentes ambientes na cidade de Ituiutaba – MG. Bioscience Journal, v. 26, n. 5, p. 817-823, 2010.
BARETTA, D.; SANTOS, J. C. P.; MAFRA, À. L.; WILDNER, L. P.; MIQUELLUTI, D. J. Fauna edáfica avaliada por armadilhas de catação manual afetada pelo manejo do solo na região oeste catarinense. Revista de Ciências Agroveterinárias, v. 2, p. 97-106, 2003.
BARETTA, D. Fauna do solo e outros atributos edáficos como indicadores da qualidade ambiental em áreas com Araucaria angustifolia no Estado de São Paulo. 2007. 158 p. Tese (Doutorado)- Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 2007.
BARETTA, D.; MALUCHE-BARETTA, C. R. D. M.; CARDOSO, E. J. B. N. Análise multivariada de atributos microbiológicos e químicos do solo em florestas com Araucaria angustifolia. Revista
BARETTA, D.; BROWN, G. G.; CARDOSO, E. J. B. N. Potencial da macrofauna e outras variáveis edáficas como indicadoras de qualidade do solo em áreas com Aracaucaria angustifolia. Acta
Zoológica Mexicana, 135-150, 2010.
BARETTA, D.; SANTOS, J. C. P.; SEGAT, J. C.; GEREMIA, E. V.; OLIVEIRA FILHO, L. C. I.; ALVES, M. V. Fauna edáfica e qualidade do solo. In: KLAUBERG FILHO, O.; MAFRA, À. L.; GATIBONI, L. C. (Eds.) Tópicos em ciência do solo. Viçosa, MG, Sociedade Brasileira de Ciência do Solo, 2011. p. 141-192.
BRANCHER, D.; ROZA-GOMES, M. F. Survey of edaphic fauna in forest fragment in the municipality of Anchieta (SC, Brazil). Biota Neotropica, v. 12, n. 3, p. 96-98, 2012.
BROWN, G. G.; MASCHIO, W.; FROUFE, L. C. M. Macrofauna do solo em sistemas agroflorestais
e Mata Atlântica em regeneração nos municípios de Barra do Turvo, SP, e Adrianópolis, PR.
Colombo: Embrapa Florestas, 2009. 51 p. (Embrapa Floresta. Documentos, 184).
CANTO, A. C. Alterações da mesofauna do solo causadas pelo uso de cobertura com plantas leguminosas na Amazônia Central. Rev. UA. Série: Ciências Agrárias, v.4/5, n. 1/2, p. 79-94, 1996. COELHO, R. A.; RICCI, M. S. F.; ESPÍNDOLA, J. A. A.; COSTA, J. R. Influência do sombreamento sobre a população de plantas espontâneas em área cultivada com cafeeiro (Coffea canephora) sob manejo orgânico. Agronomia, v. 38, n. 2, p. 23-28, 2004.
COSTA NETO, E. M. The perception of diplopoda (Arthropoda, Myriapoda) by the inhabitants of the county of Pedra Branca, Santa Teresinha, Bahia, Brazil. Acta Biólogica Colombiana, v. 12, p. 123-134, 2007.
CROTTY, F. V.; FYCHAN, R.; SCULLION, J.; SANDERSON, R.; MARLEY, C. L. Assessing the impact of agricultural forage crops on soil biodiversity and abundance. Soil Biology and
Biochemistry, v. 91, p. 119-126, 2015.
DE LIMA, K. D. R.; CAMARA, R.; CHAER, G. M.; PEREIRA, M. G.; RESENDE, A. S. Soil fauna as bioindicator of recovery of degraded areas in the Caatinga biome. Revista Caatinga, v. 30, n. 2, p. 401-411, 2017.
DORAN, J. W.; PARKIN, T. B. Defining and assessing soil quality. In: DORAN, J. B.; CLEMAN, D. C.; BEZDICEK, D. F. (Eds). Defining soil quality for a sustainable environment. Madison: Soil Science Society of America, 1994. p. 3-21. (SSSA. Special Publication, 35).
FERNANDES, J. O.; UEHARA-PRADO, M.; BROWN, G. G. Minhocas exóticas como indicadoras de perturbação antrópica em áreas de floresta atlântica. Acta Zoológica Mexicana, p. 211-217, 2010. GUIMARÃES, N. F.; FONTANETTI, A.; FUJIHARA, R. T.; GALLO, A. S.; SOUZA, M. D. B.; MORINIGO, K. P. G.; SILVA, R. F. Fauna invertebrada epigéica associada a diferentes sistemas de cultivo do cafeeiro. Coffee Science, v. 11, n. 4, p. 484-494, 2016.
GROSSO, E.; JORGE, G.; BROWN, G. G. Exotic and native earthworms in various land use systems of Central, Southern and Eastern Uruguay. Caribbean Journal of Science, v. 42, p. 294-300, 2006. KORASAKI, V.; MORAIS, J. W.; BRAGA, R. F. Macrofauna. In: MOREIRA, F. M. S.; CARES, J. E.; ZANETTI, R.; STURMER, S. L. (Eds.) O ecossistema solo: componentes, relações ecológicas e
feitos na produção vegetal. Lavras: Ed. UFLA, 2013. 352 p.
LOURENTE, E. R. P.; SILVA, R. F.; SILVA, A. D.; MARCHETTI, M. E.; MERCANTE, M. F. Macrofauna edáfica e sua interação com atributos químicos e físicos do solo sob diferentes sistemas de manejo. Acta Scientiarum. Agronomy. v. 29, n. 1, p. 17-22, 2007.
MAESTRI, R.; LEITE, M. A. S.; SCHMITT, L. Z.; RESTELLO, R. M. Efeito de mata nativa e bosque de eucalipto sobre a riqueza de artrópodos na serrapilheira. Perspectiva, v. 37, p. 31-40, 2013. OLIVEIRA, I. R. P.; FERREIRA, A. N.; VIANA JÚNIOR, A. B.; DANTAS, J. O.; SANTOS, M. J. C.; RIBEIRO, M. J. B. Diversidade de formigas (Hymenoptera: Formicidae) edáficas em três estágios sucessionais de Mata Atlântica em São Cristóvão, Sergipe. Agroforestalis News, v.1, n.1, 2016. PEREIRA, J. M.; SEGAT, J. C.; BARETTA, D.; VASCONCELLOS, R. L. F.; BARETTA, C. R. D. M.; CARDOSO, E. J. B. N. Soil Macrofauna as a Soil Quality Indicator in Native and replanted
Araucaria angustifolia Forests. Revista Brasileira de Ciência do Solo, v. 41, p. e0160261, 2017.
PEREIRA, G. H. A.; PEREIRA, M. G., ANJOS, L. H. C., AMORIM, T. A., MENEZES, C. E. G. Decomposição da serrapilheira, diversidade e funcionalidade de invertebrados do solo em um fragmento de floresta atlântica. Bioscience Journal, v. 29, n. 1, p. 1317-1327, 2013.
POMPEO, P. N.; SANTOS, M. A. B.; BIASI, J. P.; SIQUEIRA, S.; ROSA, M. G.; BARETTA, C. R. D. M.; BARETTA, D. Fauna e sua relação com atributos edáficos em Lages, Santa Catarina – Brasil. Scientia Agraria, v. 17, n.1, p. 42-51, 2016.
ROSA, M. G.; KLAUBERG FILHO, O.; BARTZ, M. L. C..; MAFRA, Á. L..; SOUSA, J. P. F. A.; BARETTA, D. Macrofauna edáfica e atributos físicos e químicos em sistemas de uso do solo no planalto catarinense. Revista Brasileira de Ciência do Solo, v. 39, n. 6, p. 1544-1553, 2015.
ROSA, M. G.; BRESCOVIT, A. D.; BARETTA, C. R. D. M.; SANTOS, J. C. P.; OLIVEIRA FILHO, L. C. I.; BARETTA, D. Diversity of soil spiders in land use and management systems in Santa Catarina, Brazil. Biota Neotropica, v. 19, n. 2, 2019.
RODRIGUES, A. B. C.; SCARAMUZZA, W. L. M. P.; SCARAMUZZA, J. F.; ROCHA, F. Atributos químicos em solo sob floresta nativa e capoeira. UNICiências, v. 14, n. 1, 2010.
ROCHA, W. O.; DORVAL, A.; PERES FILHOS, O.; VAEZ, C. A.; RIBEIRO, E. S. Formigas (Hymenoptera: Formicidae) bioindicadoras de degradação ambiental em Poxoréu, Mato Grosso, Brasil. Floresta e Ambiente, v. 22, n. 1, p. 88-98, 2015.
REZENDE, M. Q.; SOARES, L. G. S.; MÓL, A. P.; SPERBER, C. F. Interação entre processos ecológicos e geomorfológicos na diversidade e abundância de grilos (Orthoptera: Grylloidea) de serrapilheira florestal. Anais, VIII Congresso de Ecologia do Brasil. Caxambu. 2007.
SANTOS, J. B.; RAMOS, A. C.; AZEVEDO JÚNIOR, R.; OLIVEIRA FILHO, L. C. I.; BARETTA, D.; CARDOSO, E. J. B. N. Soil macrofauna in organic and conventional coffe plantations in Brazil.
Biota Neotropica, v. 18, n. 2, e20180515, 2018.
SCHLOTER, M.; DILLY, O.; MUNCH, J. C. Indicators for evaluating soil quality. Agriculture,
Ecosystems & Environment, v. 98, p. 255-262, 2003.
SILVA, J.; JUCKSCH, I.; MAIA, C. I.; FERES, A.; TAVARES, R. C. Soil faunal in management systems with coffee. Journal of Biotechnology and Biodiversity. v. 3 n. 2 p. 59-71, 2012.
SOUZA, K. K. F.; SOUSA, N. J.; SILVA, I. C.; LIMA, P. P. S.; MARQUES, E. N. Utilização de Formigas (Hymenoptera: Formicidae) como bioindicadoras em plantios de Pinus no Paraná. Revista
Brasileira de Agropecuária Sustentável, v.8, n.1, p.110-118, 2018.
SZINWELSKI, N.; ROSA, C. S.; SCHOEREDER, J.; MEWS, C. M.; SPERBER, C. F. Effects of forest regeneration on crickets: evaluating environmental drivers in a 300-year chronosequence.
International Journal of Zoology, v. 2012, p. 1-12, 2012.
TESFAHUNEGN, G. B. Soil quality indicators response to land use and soil management systems in Northern Ethiopia's catchment. Land Degradation & Development, v. 27, p. 438-448, 2016. TÓTOLA, M. R.; CHAER, G. M. Microrganismos e processos microbiológicos como indicadores da qualidade dos solos. In: ALVAREZ, V. V. H.; SCHAEFER, C. E. G. R.; BARROS, N. F.; MELLO, J. W. V.; COSTA, L. M. (Eds.) Tópicos em ciência do solo. Viçosa, MG, Sociedade Brasileira de Ciência do Solo, 2002. v. 2, p. 195-276.
VASCONCELLOS, R. L.; SEGAT, J. C.; BONFIM, J. A.; BARETTA, D.; CARDOSO, E. J. Soil macrofauna as an indicator of soil quality in an undisturbed riparian forest and recovering sites of different ages. European Journal of Soil Biology, v.58, p. 58-105, 2013.