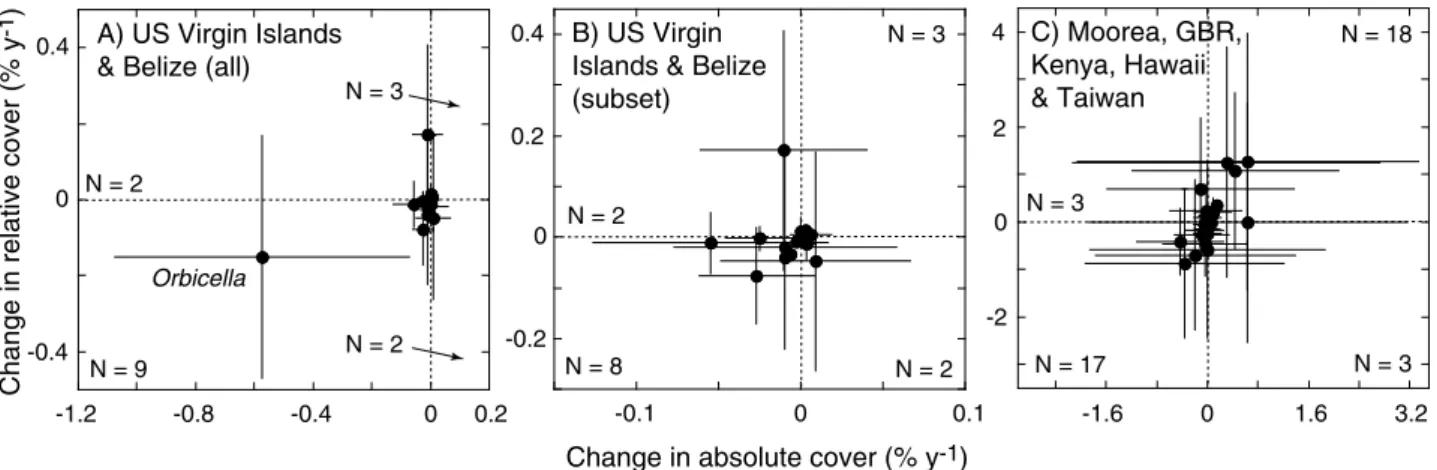
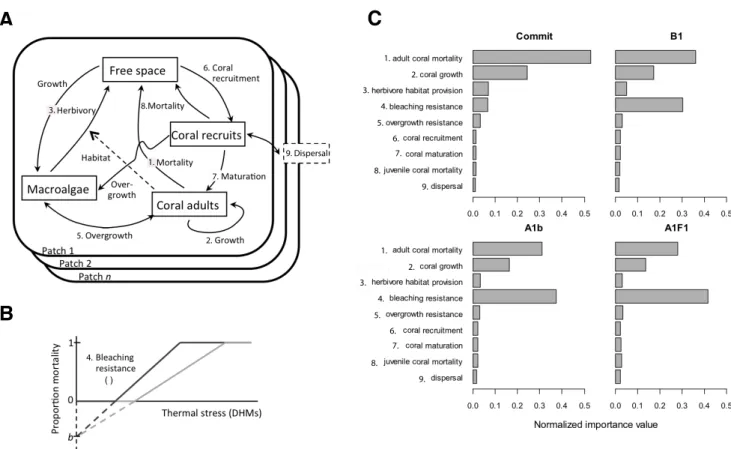
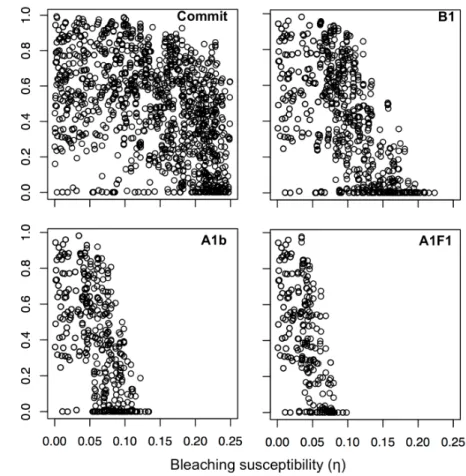
Persistence and change in community composition of reef corals through present, past, and future climates.
Texto
Imagem
Documentos relacionados
Diseases leading to accelerated decline of reef corals in the largest South Atlantic reef complex (Abrolhos Bank, eastern Brazil). Sequestration of a marine
To model the species’ current potential distribution areas and predict changes in species distributions as a result of both climate and habitat change, we first had to
In this area, there is an offshore coral reef formation, where corals were observed in loco; the water samples were collected to obtain the hydrological and plankton
Todavia, a falta de estudos sistematizados sôbre o assunto, a natureza necessàriamente fragmentária das deci sões judiciais a respeito, a existência dos textos legais calçados
Visto así, investigar la historia tanto de una palabra como de un concepto, incluyendo significados y significantes, se transforma en una tarea semejante a la del geólogo
This log must identify the roles of any sub-investigator and the person(s) who will be delegated other study- related tasks; such as CRF/EDC entry. Any changes to
To analyze the impact of nighttime oxygen-deprivation on coral health, this study evaluated changes in coral metabolism through the exposure of corals to chronic hypoxic conditions
We tested a novel biomimicry technique of animal-assisted cleaning on nursery corals prior to transplantation at a coral reef restoration site in Seychelles, Indian Ocean.. To
