Vila Real, 2013
Gemelaridade em vacas leiteiras: incidência e a
sua influência no desempenho produtivo em
explorações do Litoral Norte de Portugal
Dissertação de Mestrado em Medicina Veterinária
Jorge Carneiro Lopes
II
Agradecimentos
Esta página é dedicada a todas as pessoas que, de alguma forma, contribuíram para a elaboração desta dissertação. Pela sua valiosa contribuição, gostaria de expressar os meus sinceros agradecimentos.
Ao Professor Doutor João Carlos Caetano Simões, orientador da dissertação, por tu-do o que fez para me auxiliar neste longo e árduo trabalho, pela sugestão e orientação tu-do tema, pela enorme disponibilidade que sempre manifestou, pela motivação e incentivo que me transmitiu, pela ajuda fornecida no tratamento estatístico dos dados e análise dos resul-tados, bem como, por todos os conselhos, correções e ensinamentos que me proporcionou ao longo deste trabalho.
A todos os produtores com os quais tive a oportunidade de falar, em especial ao Sr. José António Santos, pela disponibilidade que mostrou em fornecer-me os dados, que tão importantes e valiosos foram na realização deste trabalho, pois sem eles nada disto teria sido alcançado.
Ao Nélson Bogas, colega de universidade, pela disponibilização completa dos índi-ces produtivos e reprodutivos da exploração leiteira da sua família, sem os quais não seria possível apresentar, no presente trabalho, os resultados sobre a duração da gestação e o intervalo entre partos, além da contribuição para os restantes resultados.
À Doutora Andreia Santos pela disponibilidade que manifestou durante o meu está-gio, assim como pela ajuda na obtenção dos dados e pelos ensinamentos que me transmitiu na prática clínica.
À minha família, especialmente aos meus pais e irmã, e a todos os meus amigos pe-lo apoio incondicional e por toda a ajuda prestada na elaboração deste trabalho.
III
Resumo
O presente trabalho teve como principal objetivo determinar a incidência de partos gemelares em vacas leiteiras, assim como avaliar a sua influência na produção em explora-ções do Litoral Norte de Portugal. Pretendeu-se ainda, avaliar a incidência de diversas do-enças incluindo refugos e/ou mortes em vacas com partos gemelares, assim como a viabili-dade dos vitelos ao parto e nos dias subsequentes. Foram recolhidos dados de 1109 partos ocorridos em 18 explorações leiteiras, durante os anos de 2010, 2011 e 2012. Os animais foram divididos em 2 grupos consoante apresentassem parto gemelar ou simples, corres-pondendo estes últimos ao grupo controlo, quando usado. A incidência de partos gemelares foi de 4,2% (47/1109; P<0,001), com variações (P<0,05) do n.º de fetos por parto entre ex-plorações. A incidência de partos gemelares esteve positivamente correlacionada (r=0,73; n=18; P<0,01) com o n.º de partos totais em cada exploração. A maioria dos gémeos nasci-dos foi do sexo masculino (61,7%; 58/94; P=0,001). De tonasci-dos os partos gemelares ocorrinasci-dos, 51,0% (24/47) correspondeu a partos em que os vitelos eram de sexos opostos, 36,2% (17/47) eram ambos machos e 12,8% (6/47; P<0,001) eram ambos fêmeas. Destes gémeos, 63,8% (60/94) foram nados-vivos e 36,2% (34/94; P<0,001) foram nados-mortos ou morre-ram nos primeiros 3 dias. A duração da gestação gemelar foi de 268,8 ± 7,7 dias (± D.P.; n=12) e a gestação simples de 278,1 ± 6,6 dias (n=200; P<0,001). No entanto, o intervalo entre partos foi similar entre gestações gemelares (389,8 ± 31,6 dias; n=10) e simples (403,5 ± 79,7 dias; n=133; P>0,05). Em 70,2% (33/47; P<0,001) dos casos, a vaca apresentou alte-rações patológicas, morte ou refugo até um mês pós-parto, sendo que 69,7% (23/33; P=0,001) destas anomalias corresponderam a retenção placentária. Em vacas de 2ª lacta-ção, a produção de leite aos 305 dias, foi maior quando estas apresentavam gestação ge-melar (12621,6 ± 4036,7 kg; n=11) do que simples (10185,1 ± 2533,6 kg; n=159; P<0,01). No entanto, foi observado, que independentemente da paridade, vacas com partos gemela-res apgemela-resentaram uma menor produção diária, durante o 1º mês de lactação após o parto, (34,2 ± 11,9 kg; n=31) do que vacas com partos simples (39,9 ± 11,7 kg; n=131; P<0,01). Já a produção acumulada, obtida a partir de cada um dos 5 primeiros contrastes leiteiros após o parto, foi similar (P>0,05) entre grupos, mesmo considerando o efeito da paridade. Conclu-ímos que a incidência da gemelaridade em vacas leiteiras é afetada por fatores relacionados com a dimensão (n.º de partos) da exploração. A elevada ocorrência de partos gemelares com pelo menos um macho coloca restrições, destas gestações, ao programa de reposição de fêmeas nas explorações. Embora tenha sido observada uma maior produção de leite durante a gestação gemelar, estima-se que vacas com partos gemelares produzam, durante o 1º mês após o parto, menos leite (kg/dia) do que vacas com partos simples,
provavelmen-IV
te devido à elevada ocorrência de doenças, principalmente do foro reprodutivo. No entanto, esta diminuição não parece afetar a produção acumulada posterior.
Palavras-chave: Epidemiologia; Partos Gemelares; Produção de Leite; Retenção
V
Abstract
The present study aimed to determine the incidence of twinning in dairy cows, as well as to evaluate its influence on production in farms of the North Coast of Portugal. It also in-tended to assess the incidence of several diseases including culling and/or deaths in cows with twin births, as well as the viability of calves at birth and on subsequent days. Data were collected from 1109 births in 18 dairy farms during the years 2010, 2011 and 2012. The ani-mals were divided into 2 groups according to presentation of twin or simple birth, where the last ones corresponded to the control group, when used. The incidence of multiple births was 4.2% (47/1109; P<0.001), with variations (P<0.05) on the n.º of fetuses per calving between farms. The incidence of twin births was positively correlated (r=0.73; n=18; P<0.01) with the total n.º of births in each farm. Most twins born were male (61.7%; 58/94; P=0.001). Of all twin births occurred, 51.0% (24/47) corresponded to births in which the calves were of oppo-site sexes, 36.2% (17/47) were both males and 12.8% (6/47; P<0.001) were both females. It was observed that 63.8% (60/94) of these twins were live births and 36.2% (34/94; P<0.001) were stillbirths or died within the first 3 days. The gestation length of twin pregnancy was 268.8 ± 7.7 days (± SD; n=12) and single pregnancy was 278.1 ± 6.6 days (n=200; P<0.001). However, the calving interval was similar between twin pregnancies (389.8 ± 31.6 days; n=10) and single pregnancies (403.5 ± 79.7 days; n=133; P>0.05). In 70.2% (33/47; P<0.001) of the cases, the cow presented pathological changes, died or was culled until one month postpartum, and 69.7% (23/33; P=0.001) of these abnormalities corresponded to re-tained placenta. In cows of 2nd lactation, milk yield at 305 days was higher when they pre-sented a twin pregnancy (12621.6 ± 4036.7 kg; n=11) than a simple pregnancy (10185.1 ± 2533.6 kg; n=159; P<0.01). However, it was observed that, regardless of parity, cows with twin births had a lower daily production during the 1st month of lactation after parturition (34.2 ± 11.9 kg; n=31) than cows with simple births (39.9 ± 11.7 kg; n=131; P<0.01). The accumulated production, obtained from each of the first 5 postpartum dairy contrasts were similar (P>0.05) between groups, even considering the effect of parity. We conclude that the incidence of twinning in dairy cows is affected by factors related to the size (number of births) of the farm. The high occurrence of twin births with at least one male puts restrictions of these pregnancies to the program of replacement heifers on farms. Although there was a higher milk yield during twin pregnancy, it is estimated that cows with twin births produce less milk (kg/day) during the 1st month after parturition, than cows with single births, probably due to the high occurrence of diseases, especially reproductive diseases. However, this decrease does not seem to affect further cumulative production.
VI
Keywords: Dairy Cows; Epidemiology; Milk Production; Retained Placenta; Twin Births;
VII
Índice Geral
Agradecimentos ... II Resumo ...III Abstract ... V Índice de figuras ... X Índice de tabelas ... XI Índice de abreviaturas e siglas ... XII1. Introdução e objetivos ... 1
2. Revisão bibliográfica: a gemelaridade ... 3
2.1. Desenvolvimento folicular ... 3
2.2. Mecanismos responsáveis pela gemelaridade ... 4
2.3. Gémeos monozigóticos... 5
2.4. Gémeos dizigóticos ... 5
2.5. Fatores predisponentes ... 6
2.5.1. Raça ... 6
2.5.2. Paridade ... 7
2.5.3. Sazonalidade, temperatura e alimentação ... 8
2.5.4. Seleção genética ...10
2.5.5. Taxa de ovulação ...11
2.5.6. Produção leiteira ...12
2.5.7. Alimentação ...14
2.5.8. Somatotropina bovina recombinante (rbST) ...14
2.6. Impacto económico da gemelaridade ...16
2.6.1. Impacto da gemelaridade no embrião/vitelo ...16
2.6.1.1. Morte embrionária tardia e morte fetal ...16
VIII
2.6.1.3. Freemartinismo ...18
2.6.2. Impacto da gemelaridade nas vacas ...20
2.6.2.1. Produção de leite na subsequente lactação ...20
2.6.2.2. Duração da gestação ...21
2.6.2.3. Distócia ...22
2.6.2.4. Retenção placentária ...23
2.6.2.5. Metrite ...24
2.6.2.6. Fertilidade ...24
2.6.2.7. Doenças metabólicas no pós-parto ...25
2.6.2.8. Taxa de refugo ...26
3. Material e métodos ...27
3.1. Animais e dados clínicos ...27
3.2. Registos zootécnicos ...27
3.3. Análise estatística ...28
4. Resultados ...29
4.1. Incidência de partos gemelares nas explorações ...29
4.2. Caracterização dos partos gemelares ...31
4.2.1. Caracterização do sexo e da viabilidade dos fetos ...31
4.2.2. Paridade, duração da gestação e intervalo entre partos das mães ...32
4.2.3. Afeções, refugo e/ou mortalidade associadas a partos gemelares ...32
4.3. Influência da gemelaridade na produção leiteira. ...33
4.3.1. Produção na lactação anterior ao parto gemelar ...33
4.3.2. Produção na lactação subsequente ao parto gemelar ...34
5. Discussão ...35
5.1. Incidência de partos gemelares nas explorações ...35
5.2. Caracterização dos partos gemelares ...35
IX
5.2.2. Paridade, duração da gestação e intervalo entre partos das mães ...37
5.2.3. Afeções, refugo e/ou mortalidade associadas a partos gemelares ...38
5.3. Influência da gemelaridade na produção leiteira ...39
5.3.1. Produção na lactação anterior ao parto ...39
5.3.2. Produção na lactação subsequente ao parto gemelar ...40
6. Conclusão ...42
X
Índice de figuras
Figura 1 - Desenvolvimento folicular durante o ciclo éstrico em vacas leiteiras, segundo um
padrão de duas ondas foliculares, com a seleção de um único folículo dominante…………..4
Figura 2 - Percentagem de gemelaridade segundo a época do ano e paridade, para vacas da raça Frísia em clima temperado………9
Figura 3 - Incidência de múltipla ovulação em vacas com diferentes produções durante os 14 dias que precedem o estro………...11
Figura 4 - Distribuição do n.º de partos gemelares de acordo como n.º de partos totais em cada exploração………..29
Figura 5 - Regressão entre o n.º de partos totais e a percentagem de partos gemelares em cada exploração………..30
Figura 6 - Distribuição dos partos gemelares por exploração……….30
Figura 7 - Caracterização do sexo dos vitelos………..31
Figura 8 - Caracterização dos partos segundo o sexo dos vitelos……….31
XI
Índice de tabelas
Tabela 1 - Incidência de partos múltiplos segundo a raça de bovinos……….7 Tabela 2 - Efeito da paridade na taxa de gemelaridade (%) em vacas leiteiras………7 Tabela 3 - Valores médios da produção de leite, % e kg de gordura aos 305 dias em lacta-ção entre vacas com gestalacta-ção gemelar e simples.………...13 Tabela 4 - Efeito do tipo de parto na viabilidade dos vitelos (%)………18 Tabela 5 - Duração da gestação entre vacas com gestação gemelar e simples……….21 Tabela 6 - Incidência de retenção placentária em vacas com partos simples e gemelares..23 Tabela 7 - Dias em aberto e inseminações por conceção entre vacas com parto gemelar e simples………..25 Tabela 8 - Taxa de refugo em vacas com partos gemelares e simples………26 Tabela 9 - Ocorrência de doenças, refugo e/ou morte no pós-parto de vacas com gestações gemelares……….33 Tabela 10 - Produção diária de leite após parto simples e gemelar, de acordo com dados do contraste leiteiro, independentemente da paridade………..34
XII
Lista de abreviaturas e siglas
ABLN – Associação para o Apoio à Bovinicultura Leiteira do Norte ANOVA – Análise de Variância
DAE – Deslocamento de Abomaso à Esquerda EUA – Estados Unidos da América
F – Feminino
FDA – “Food and Drug Administration” FF – Feminino-Feminino
FSH – Hormona Folículo-Estimulante (“Follicle-Stimulating Hormone”) IA – Inseminação Artificial
JMP – “John’s Macintosh Project” kg – Quilogramas (“Kilograms”)
LH – Hormona Luteinizante (“Luteinizing Hormone”) M – Masculino
MARC – “Meat Animal Research Center” MF – Masculino-Feminino
MM – Masculino-Masculino mm – Milímetros
n.º - Número
SAS – “Statistical Analysis System” UE – União Europeia
USDA – Departamento de Agricultura dos Estados Unidos (“United States Department of Agriculture”)
1
1. Introdução e objetivos
Nos últimos anos, o setor leiteiro tem vindo a sofrer uma evolução significativa. Gra-ças ao desenvolvimento de novas técnicas e procedimentos intensivos, as explorações lei-teiras dos países mais desenvolvidos, aumentaram de forma significativa o número de ani-mais por exploração, assim como a produção de leite por animal, o que conduziu ao aumen-to da produtividade e rentabilidade. Todo este sucesso foi fruaumen-to, principalmente, de uma se-leção genética intensiva de animais de elevada produção, associado ao melhoramento da nutrição e do maneio geral dos animais. No entanto, tem sido registado uma diminuição da performance reprodutiva, com uma queda nos índices de fertilidade das vacas, e um aumen-to do intervalo paraumen-to-conceção, (Lucy, 2001; López-Gatius, 2003), principalmente, em explo-rações com animais de elevada produção de leite.
A eficiência reprodutiva está extremamente ligada ao sucesso económico das explo-rações (Briitt, 1985; Plaizier et al., 1997; Meadows et al., 2005). Assim sendo, torna-se claro a importância de um controlo reprodutivo dos animais, especialmente em explorações com um elevado sistema de produção, pois é a seguir ao parto que se inicia uma nova lactação.
Na produção leiteira, a gemelaridade é uma situação que afeta a eficiência reproduti-va e produtireproduti-va das reproduti-vacas. Esta ocorre com pouca incidência reproduti-variando entre 2,2 e 6,9% (Kin-sel et al., 1998; Silva del Rio et al., 2007). No entanto, nos últimos anos, o número de vacas com gestações gemelares parece ter vindo a aumentar, estimando-se que atualmente atinja os 9%, podendo mesmo ultrapassar os 12% em algumas explorações (Silva del Rio et al., 2007).
A elevada produção de leite tem sido descrita como sendo o fator mais importante para o recente aumento da gemelaridade (Kinsel et al., 1998), e tem sido relacionada com a incidência de dupla ovulação (Fricke e Wiltbank, 1999; López et al., 2005). Deste modo, é previsível que, nos próximos anos, a gemelaridade continue a aumentar, juntamente com a produção de leite.
Sendo a gemelaridade um fenómeno que resulta da capacidade reprodutiva dos animais, esta acaba por condicionar, em muito, as vacas e os vitelos recém-nascidos. Vacas com gestações gemelares acabam por estar sujeitas a determinados condicionalismos que irão determinar o sucesso da futura lactação. Assim sendo, há um aumento do risco de dis-tócia (Cady e Van Vleck, 1978; Gregory et al., 1990, 1996), levando ao aumento de retenção placentária (Turman et al., 1971; Bellows et al., 1974; Anderson et al., 1982; Guerra-Martinez et al., 1990), e ao aumento do intervalo parto-1ª inseminação (Erb e Morrison, 1959; Turman et al., 1971; Gregory et al., 1990).
2
Não só a vaca está suscetível aos efeitos de uma gestação gemelar. Os vitelos re-cém-nascidos acabam também por sofrer com isso, evidenciado pelo aumento da taxa de mortalidade, assim como pela diminuição da sua taxa de desenvolvimento (Gregory et al., 1996), que pode ser resultante duma diminuição do seu peso à nascença (Gregory et al., 1996; Echternkamp e Gregory, 1999).
Desta forma, torna-se claro que vacas com gestações gemelares acabam por enfren-tar grandes desafios após o parto, evidenciados pela elevada incidência de doenças ou dis-túrbios que interferem com a sua alta capacidade de produção de leite. Para além disto, os vitelos recém-nascidos têm maior probabilidade de sucumbir, representando um sério pro-blema a longo prazo para a exploração, no que diz respeito aos animais de reposição. Tudo isto acaba, inevitavelmente, por conduzir a enormes perdas económicas, as quais nos dias de hoje, podem determinar o sucesso ou o fracasso das atuais explorações leiteiras do nos-so país.
O objetivo principal deste trabalho foi determinar a incidência de gemelaridade em vacas leiteiras e determinar a sua influência na produção. Pretendemos ainda, avaliar a inci-dência de diversas patologias em vacas com partos gemelares, assim como a viabilidade dos vitelos ao parto e nos dias subsequentes.
3
2. Revisão bibliográfica: a gemelaridade
Os bovinos são uma espécie monotócica, o que significa que na maioria das vezes uma gestação com sucesso resulta no nascimento de um único vitelo. No entanto, nem sempre isto acontece, dando origem ao nascimento de mais que um vitelo no momento do parto. Para se compreender os mecanismos que levam ao aparecimento de gestações ge-melares, será necessário fazer uma breve abordagem ao desenvolvimento folicular.
2.1. Desenvolvimento folicular
Os ovários possuem várias estruturas no seu córtex que se designam por folículos. Cada folículo ovárico contém um único oócito, o qual é expelido em direção ao oviduto du-rante o processo de ovulação. No entanto, nem todos os folículos presentes no ovário vão crescer e desenvolver-se para depois ovularem. Existe assim, um mecanismo fisiológico responsável pela regulação do número de folículos que se tornam dominantes em cada on-da folicular, resultando normalmente, na seleção de um único folículo com capacion-dade ovu-latória.
Nos bovinos, o crescimento folicular que ocorre durante o ciclo éstrico, é regulado segundo um padrão de duas ou três ondas foliculares (Fig. 1). Normalmente, as vacas pri-míparas e multíparas exibem um padrão de duas ondas foliculares por ciclo éstrico, enquan-to as nulíparas (novilhas) têm maior tendência a exibir um padrão de três ondas foliculares por ciclo éstrico (Lucy et al., 1992). Em vacas leiteiras, o número de ondas foliculares por ciclo éstrico depende de alguns fatores que variam entre cada animal (Fortune et al., 1988). Alguns desses fatores incluem a quantidade de alimento ingerido (Murphy et al., 1991), a idade, a paridade e o momento da lactação (Lucy et al., 1992).
Assim, por volta do momento em que o folículo dominante (pré-ovulatório) do ciclo éstrico anterior ovula, inicia-se uma nova onda folicular, na qual, um pequeno grupo de folí-culos começa a crescer (Adams et al., 1992). Estes vão crescendo ao mesmo ritmo até que se atinge um ponto crítico em que apenas um folículo continua a crescer para se tornar do-minante, enquanto os outros entram em regressão (regressão dos folículos subordinados; Savio et al., 1988). Este processo de seleção é conhecido por desvio folicular (Ginther et al., 1996).
Deste modo, o folículo selecionado para ser dominante cresce e desenvolve-se, aca-bando por adquirir capacidade ovulatória. Com a presença de um estímulo ovulatório natural ou induzido, inicia-se o processo de ovulação, fazendo com que seja libertado um único
4
Dias após ovulação
D iâmet ro f ol icul ar (m m )
oócito, proveniente do folículo dominante. Se os eventos seguintes, que vão desde a fecun-dação até ao parto, ocorrerem normalmente, a gestação resultará no nascimento de um único vitelo.
Figura 1 – Desenvolvimento folicular durante o ciclo éstrico em vacas leiteiras, segundo um padrão
de duas ondas foliculares, com a seleção de um único folículo dominante. = Folículos em cresci-mento antes da seleção do folículo dominante; = Folículos dominantes; = Folículos em regressão. Fonte: Adaptado de Fricke e Shaver (2001).
De realçar que nestes processos de dinâmica folicular, intervêm complexos meca-nismos hormonais, cuja descrição não é do âmbito do presente trabalho, embora se encon-trem referências parciais nos subcapítulos subsequentes.
2.2. Mecanismos responsáveis pela gemelaridade
Em bovinos, a gemelaridade pode resultar de dois mecanismos diferentes. Consoan-te o mecanismo subjacenConsoan-te, esta pode ser classificada em dois tipos: monozigótica ou di-zigótica (Fricke, 2001).
O oócito, após rotura do folículo pré-ovulatório, vai ser fecundado no oviduto, por um espermatozóide, originando uma única célula, denominada de zigoto. No entanto, em raras situações, o zigoto acaba por, subsequentemente, sofrer uma divisão extra após fecunda-ção. Assim, originam-se dois embriões a partir de uma única célula, os quais vão
desenvol-5
ver-se durante o desenvolvimento uterino, dando origem a gémeos monozigóticos (Fricke, 2001).
Em outras situações, dois folículos do mesmo ciclo éstrico são selecionados para continuarem a crescer a partir do grupo de folículos em crescimento durante uma onda foli-cular, resultando num fenómeno chamado de co-dominância. Se ocorrer um estímulo apro-priado para a ovulação, quer seja natural ou induzido, enquanto ambos os folículos domi-nantes estiverem presentes, serão libertados dois oócitos, um de cada folículo respetiva-mente. Se os eventos seguintes, que vão desde a fecundação até ao parto, ocorrerem nor-malmente para ambos os oócitos, então resultará no nascimento de dois vitelos, os quais serão chamados de gémeos dizigóticos (Fricke, 2001).
2.3. Gémeos monozigóticos
Os gémeos monozigóticos são vulgarmente chamados de gémeos verdadeiros. Es-tes são genotipicamente e fenotipicamente idênticos, e por isso, são sempre do mesmo se-xo. O mecanismo pelo qual surgem gémeos monozigóticos não é muito bem compreendido, mas pode ser considerado como a clonagem natural de um zigoto in vivo (Fricke e Shaver, 2001).
A percentagem de gémeos monozigóticos em bovinos é muito reduzida, com estima-tivas a rondar entre 7,4% (Erb e Morrison, 1959) e 13,6% (Ryan e Boland, 1991) de todos os partos gemelares, ou ainda menos de 0,3% de todos os partos (simples e gemelares). Johansson et al. (1974) descreveram uma taxa de gémeos monozigóticos a variar entre 13,5 a 25% dos gémeos homozigóticos, dependendo da raça e tendendo para aumentar com a paridade. Deste modo, gémeos monozigóticos ocorrem com pouca frequência, surgindo apenas em raros partos gemelares.
2.4. Gémeos dizigóticos
A maioria dos gémeos em bovinos é dizigótica (Erb e Morrison, 1959; Johansson et
al., 1974; Ryan e Boland, 1991). Os gémeos dizigóticos tanto podem ser do mesmo sexo
como do sexo oposto, e não são mais parecidos fenotipicamente ou genotipicamente do que irmãos, mantendo os mesmos progenitores, nascidos em diferentes gestações. Isto sugere que a gemelaridade e a taxa de ovulação são características fortemente associadas uma à outra (Morris e Day, 1986; Van Vleck et al., 1991).
6
2.5. Fatores predisponentes
A gemelaridade em bovinos é uma característica complexa causada por vários fato-res, que incluem componentes fisiológicos, ambientais e genéticos, podendo mesmo ser considerada como sendo multifatorial. O aumento da gemelaridade, que se tem vindo a re-gistar ao longo do tempo em vacas leiteiras, sugere uma mudança com concorrência de um ou mais destes fatores. Torna-se essencial ter um conhecimento claro destes fatores, para que possam ser elaboradas estratégias de maneio para explorações com elevadas taxas de gemelaridade.
Os fatores predisponentes para a gemelaridade em vacas leiteiras incluem a raça, a paridade, a sazonalidade, a seleção genética, a taxa de ovulação, a produção leiteira, a ali-mentação e a utilização de substâncias exógenas como a somatotropina bovina recombi-nante (rbST).
2.5.1. Raça
Desde há muito tempo, sabe-se que a frequência da gemelaridade varia entre as di-ferentes raças de bovinos. Contudo, estas variações segundo a raça, devem ser interpreta-das cuidadosamente, visto que o clima e a nutrição podem confundir esta relação.
Rutledge (1975) fez um estudo para comparar as diferenças nas taxas de gemelari-dade entre as diferentes raças de bovinos, acabando por descobrir diferenças significativas nas taxas de gemelaridade entre raças de carne (0,4 a 0,9%) e raças leiteiras (1,3 a 8,9%). A taxa de gemelaridade para raças leiteiras foi de 3,4% para a raça Holstein (n=25397), 1,3% para a raça Jersey (n=3537), 1,3% para a raça Guernsey (n=3263), 2,8% para a raça Ayrshire (n=889) e 8,9% para a raça Parda Suíça (n=305). Também Cady e Van Vleck (1978) analisaram dados de partos ocorridos em explorações leiteiras norte-americanas (Tabela 1), onde registaram uma incidência de gemelaridade de 4,7% para a raça Holstein (n=21174), 2,3% para a raça Guernsey (n=300), 1,8% para a raça Jersey (n=545), 2,6% para a raça Ayrshire (n=229) e 4,0% para a raça Parda Suíça (n=49). Deste modo, verificou-se que as vacas da raça Holstein exibem constantemente as maiores taxas de gemelaridade (Cady e Van Vleck, 1978; Foote, 1981).
Para além das variações registadas nos vários estudos, a taxa de gemelaridade varia amplamente entre vacas do mesmo efetivo. Kinsel et al. (1998) verificaram que no mesmo estudo, a taxa média de gemelaridade, em vacas norte-americanas da raça Holstein, foi de 2,4%, com diferenças entre efetivos a variar entre 0 a 9,6%.
7
Tabela 1 - Incidência de partos múltiplos segundo a raça de bovinos.
Raça Partos totais Partos
múlti-plos (%) Gémeos (n.º) Trigémeos (n.º) Quadrigémeos (n.º) Holstein 21174 4,75 997 7 1 Guernsey 300 2,33 7 0 0 Jersey 545 1,83 10 0 0 Ayrshire 229 2,62 6 0 0 Parda Suíça 49 4,08 2 0 0
Fonte: Adaptado de Cady e Van Vleck (1978).
2.5.2. Paridade
Alguns estudos realizados nas décadas de 40 e 50 revelaram a existência de uma associação positiva entre a gemelaridade e a paridade (n.º de parições) em vacas da raça Holstein-Frísia (Pfau et al., 1948; Erb e Morrison, 1959). Pfau et al. (1948) observaram que a gemelaridade em vacas leiteiras aumenta com a paridade, com cerca de 0,7% para as va-cas primíparas e 5,0% para vava-cas de segunda paridade. Já Berry et al. (1994) referem, para vacas de pelo menos terceira paridade, valores muito próximos de 10% (Tabela 2).
Tabela 2 - Efeito da paridade na taxa de gemelaridade (%) em vacas leiteiras. N.º de parições Referência N.º de partos 1 2 3 4 5 6 Pfau et al. (1948) 937 0,7 5,0 4,2 5,0 7,0 6,7a Erb e Morrison (1959) 7387 1,3 4,4 5,3 4,6 5,8 6,0 Nielen et al. (1989) 11951 0,8 2,7 4,1 4,5 4,9 4,8a Eddy et al. (1991) 19755 0,9 2,1 3,5 3,4 3,7 3,2 Ryan e Boland (1991) 24843 1,0 7,0 7,5 7,9 9,1a - Berry et al. (1994) 19497 1,3 6,0 9,4a - - - Kinsel et al. (1998) 52362 1,0 2,9 3,2 3,9 3,3 4,1a Fonte: Adaptado de Wiltbank et al. (2000). a Inclui todas as vacas com paridade igual ou superior à referida.
Da mesma forma, Erb e Morrison (1959) analisaram dados de 7387 partos, recolhi-dos durante 30 anos, e verificaram uma taxa de gemelaridade inferior para vacas primíparas (1,3%), do que para vacas de segunda parição (4,4%).
8
Um acentuado aumento da gemelaridade verifica-se entre as vacas primíparas e as vacas de segunda paridade. Esta continua a aumentar para paridades superiores, mas a uma taxa menor. O efeito da paridade na taxa de gemelaridade não é compreendido clara-mente, mas pode ser explicado pelo aumento da capacidade de as vacas, à medida que envelhecem, suportarem com mais sucesso uma gestação gemelar; pelo aumento da taxa de dupla ovulação ou ainda pela interação de ambos os fatores. Embora haja registo de que as vacas com gestação gemelar tenham um aumento da capacidade uterina para manterem a gestação (Ryan e Boland, 1991), vários estudos defendem que o aumento da taxa de ovu-lação é o fator mais importante para a explicação do efeito da paridade na ocorrência de gemelaridade (Wiltbank et al., 2000).
López-Gatius et al. (2005) observaram um aumento na dupla ovulação à medida que a paridade aumentava, desde 6,7%, 16,6% até 25,0% para vacas em 1ª, 2ª e ≥3ª paridade, respetivamente.
Segundo Kidder et al. (1952), Fricke et al. (1998) e Fricke e Wiltbank (1999) a inci-dência de dupla ovulação em vacas leiteiras ronda os 14%, e tal como a inciinci-dência de ge-melaridade, aumenta com a paridade (Labhsetwar et al., 1963).
2.5.3. Sazonalidade, temperatura e alimentação
A época ou o mês em que ocorre a conceção influencia a incidência de gemelaridade (Fig. 2). No início da década de 30, Johansson (1932) observou que durante o ano, existiam dois picos de gemelaridade em explorações suecas, correspondendo a conceções ocorridas no início da primavera e durante os meses de setembro e outubro. Apesar de vários estudos terem revelado os efeitos da sazonalidade na incidência de gemelaridade, outros não permi-tiram demonstrar esta relação (Hendy e Bowman, 1970). Por exemplo, na Holanda foi ob-servado um aumento sazonal de gemelaridade entre os meses de abril e setembro (Nielen
et al., 1989), e na Arábia Saudita foi observado desde maio até ao final de junho (Ryan e
Boland, 1991), enquanto que num estudo realizado na América do Norte não foram deteta-dos efeitos da sazonalidade sobre a gemelaridade (Kinsel et al., 1998).
Atualmente, especula-se muito sobre a influência deste fator na incidência de geme-laridade, verificando-se uma maior tendência nos meses mais quentes.
As diferenças de temperatura registadas em cada mês podem explicar os efeitos sa-zonais na incidência de gemelaridade. Segundo Roth et al. (2000), nos meses mais quentes, o aumento da temperatura promove o crescimento e desenvolvimento de folículos maiores
9
em cada onda folicular, mas com menor dominância entre eles. Isto pode ser devido a alte-rações nos níveis de gonadotropinas (FSH e LH) e de hormonas ováricas (estradiol e inibi-na) que se encontram em circulação, pois um folículo dominante de uma vaca em stress térmico acaba por produzir níveis mais baixos de estradiol (Wolfenson et al., 1997), assim como de inibina (Wolfenson et al., 1995), traduzindo-se num aumento dos níveis de FSH (Ingraham et al., 1974). Este ambiente endócrino permite que folículos subordinados cres-çam e adquiram capacidade para ovularem (Badinga et al., 1993; Wolfenson et al., 1995; Vasconcelos et al., 1998; Wilson et al., 1998; Roth et al., 2000), aumentando a probabilidade de ocorrer uma dupla ou múltipla ovulação, durante este período.
Figura 2 - Percentagem de gemelaridade segundo a época do ano e paridade, para vacas da raça Frísia em clima temperado. Fonte: Adaptado de Ryan e Boland (1991).
Para além disto, a atividade endócrina do aparelho reprodutor é sensível ao fotoperí-odo, especulando-se que este desempenhe um papel importante na incidência sazonal de gemelaridade (Rutledge, 1975; Nielen et al., 1989). O fotoperíodo altera a frequência pulsátil de LH e a capacidade de resposta ao feedback negativo do estradiol em ovinos (Robinson
et al., 1985). Segundo Hansen et al. (1982), novilhas expostas a um período de luz de 18
horas apresentaram uma maior resposta de LH ao estradiol do que novilhas expostas a um período de luz de 8 horas.
É importante referir que uma alimentação mais rica durante os meses de primavera e de outono poderá levar ao aumento da incidência de gemelaridade, assemelhando-se ao “flushing” realizado em ovinos.
Época do ano G emel arid ad e ( %) Paridade 1 Paridade 2 Paridade 3 Paridade 4 Paridade 5
Jan/Fev Mar/Abr Mai/Jun Jul/Ago Set/Out Nov/Dez zz
10
Por fim, Cady e Van Vleck (1978) e Nielen et al. (1989) verificaram uma diminuição da viabilidade embrionária nos meses mais quentes de verão, quando comparada com a viabilidade embrionária dos meses mais frescos de outono.
Serão necessários mais estudos para podermos quantificar os efeitos da sazonalida-de na gemelaridasazonalida-de, assim como os fatores responsáveis pelas flutuações sazonais sazonalida-de ge-melaridade.
2.5.4. Seleção genética
A seleção genética de animais com capacidade para aumentarem a taxa de gemela-ridade foi demonstrada durante um longo ensaio realizado no USDA MARC, em Clay Cen-ter, Nebraska, EUA. Verificou-se que a seleção genética de animais com elevada incidência de gemelaridade, aliada mais tarde, ao aumento da taxa de ovulação, conduziu a um au-mento de 4% em 1984 para 31% em 1995 (Gregory et al., 1997). Já em 1997, mais de 35% de todos os nascimentos registados no grupo experimental eram gemelares (Echternkamp e Gregory, 1999). Apesar do grupo experimental ser constituído por várias raças de bovinos de carne, e dos partos serem controlados para ocorrerem todos numa dada altura do ano, muitos dos animais selecionados para formar este grupo, no início da experiência, eram da raça Holstein, tendo sido adquiridos a explorações leiteiras locais, com base na sua história pregressa de gemelaridade (Gregory et al., 1990). Apesar das estimativas de heritabilidade e repetibilidade para a gemelaridade serem baixas (0,08 e 0.09), respetivamente (Van Vleck
et al., 1991; Gregory et al., 1997), uma história anterior de gemelaridade é um fator de risco
para o subsequente nascimento de gémeos (Bendixen et al., 1989; Nielen et al., 1989). Já que a gemelaridade é indesejável em vacas leiteiras, seria de esperar ver uma pressão de seleção contra esta. No entanto, a incidência de gemelaridade em vacas da raça Holstein é elevada, e tem vindo a aumentar ao longo dos tempos (Kinsel et al., 1998), visto que tem sido feita uma seleção genética de vacas com elevada capacidade de produção de leite, acabando esta por ser apontada como a responsável pelo aumento da incidência de gestações gemelares (Johanson et al., 2001), assim como as práticas de maneio das explo-rações de alta produção, pois também reduzem o risco de perda de gestações gemelares (López-Gatius e Garcia-Ispierto, 2010).
11
2.5.5. Taxa de ovulação
Em vacas leiteiras, a gemelaridade e a taxa de ovulação são características que es-tão fortemente associadas (Morris e Day, 1986; Ryan e Boland, 1991). Kidder et al. (1952) avaliaram 634 vacas por palpação transretal, registando uma incidência de dupla ovulação de 13%. Mais tarde, em estudos realizados em explorações leiteiras norte-americanas, ob-servou-se que a incidência de dupla ovulação em vacas leiteiras foi de 14,1% (28/199) para vacas sincronizadas com o programa Ovsynch (Fricke et al., 1998; Fricke e Wiltbank, 1999), e 20,5% para vacas com cios espontâneos (López et al., 2004). No mesmo sentido, López-Gatius et al. (2005) verificaram uma taxa de dupla ovulação de 15,5% (n=1795), não regis-tando diferenças entre vacas sincronizadas e vacas com cios espontâneos.
A taxa de dupla ovulação, tal como a incidência de gemelaridade, aumenta com a paridade (Labhsetwar et al., 1963; Fricke e Wiltbank, 1999). Poucos estudos avaliaram os fatores que influenciam a dupla ovulação em vacas leiteiras. Contudo, uma elevada produ-ção leiteira, próxima do momento de ovulaprodu-ção, tem sido associada ao aumento da frequên-cia de dupla ovulação (Fricke e Wiltbank, 1999). Esta relação é descrita na figura 3.
Figura 3 – Incidência de múltipla ovulação em vacas com diferentes produções durante os 14 dias
que precedem o estro. Fonte: Adaptado de López et al. (2005).
Estes resultados devem ser interpretados com cautela, pois apesar da dupla ovula-ção preceder a formaovula-ção de gémeos dizigóticos, nem sempre resultará em gemelaridade. Mais pesquisa será necessária para compreender completamente a relação entre a dupla ovulação e a gemelaridade, assim como os mecanismos responsáveis pela dupla ovulação em vacas leiteiras. M úl ti pl a ovul açã o (% )
12
2.5.6. Produção leiteira
A possível relação entre a gemelaridade e a produção de leite tem sido estudada ao longo do tempo, pois as estratégias de maneio visam maximizar a produção de leite por va-ca. Se a gemelaridade está relacionada com a produção leiteira, então será expectável um aumento da mesma, tendo em conta o aumento anual da produção de leite por vaca que se tem vindo a registar nas últimas décadas, fruto de uma seleção genética apurada e do sur-gimento de novas técnicas de reprodução, nomeadamente a inseminação artificial.
Geralmente, a taxa de gemelaridade para a maioria das explorações de carne é infe-rior a 1% (Rutledge, 1975), enquanto que para as explorações leiteiras situa-se entre os 2,2 e 6,9% (Kinsel et al., 1998; Silva del Rio et al., 2007). Deste modo, percebe-se que a aptidão leiteira poderá levar ao aumento da gemelaridade.
Em estudos realizados nas décadas de 40 e 50, verificaram-se pequenas diferenças na produção de leite, por parte de vacas com gestações gemelares (Meadows e Lush, 1957). Em outros estudos, tem sido observado uma associação positiva entre a produção de leite e a incidência de gemelaridade em vacas leiteiras (Nielen et al., 1989; Kinsel et al., 1998), ao contrário de outros que não registaram qualquer relação entre estes (Pfau et
al.,1948; Kay, 1978; Deluyker et al., 1991).
Wood (1975) realizou um estudo na Grã-Bretanha, com a duração de dois anos, no qual avaliou a produção de leite de vacas da raça Frísia que tiveram um parto simples no primeiro ano de estudo e depois tiveram um parto gemelar no segundo (n=323), assim como vacas que tiveram partos simples nos dois anos de estudo (n=2009). Para as vacas que primeiro registaram um parto simples, e no ano seguinte tiveram um parto gemelar, verifi-cou-se um aumento significativo na produção de leite, do que em vacas com partos simples em ambos os anos.
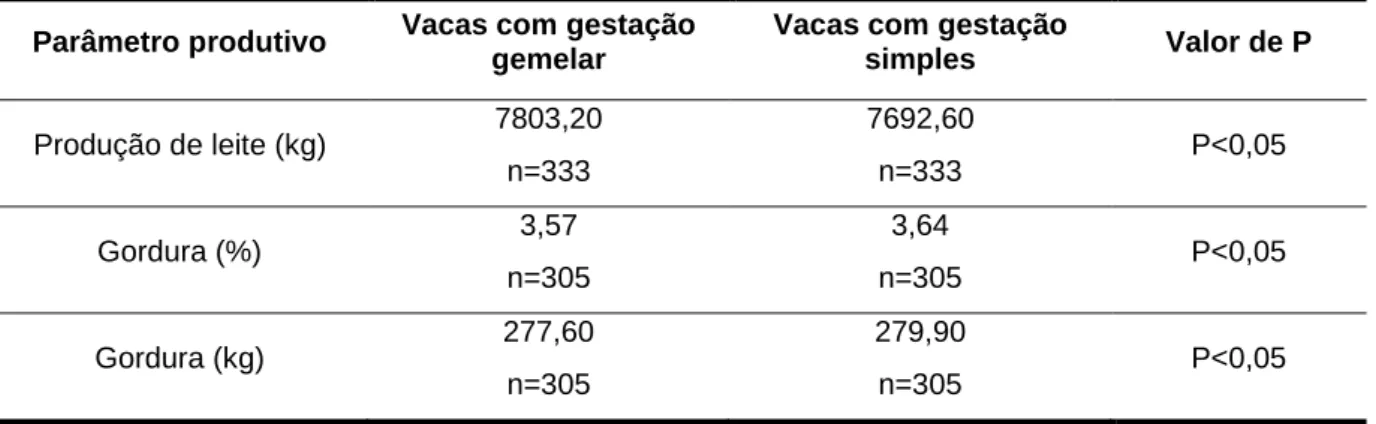
Dados recolhidos entre 1965 e 1975, indicaram que vacas com partos gemelares produziram mais leite durante a lactação em que se encontravam gestantes de gémeos do que em qualquer outra lactação (P<0,01; Kay, 1978). Do mesmo modo, Chapin e Van Vleck (1980) verificaram que vacas com gestações gemelares eram melhores produtoras, pois registaram produções entre 7705 e 7803 kg de leite por lactação, relativamente a vacas com gestações simples, cujas produções se situaram entre 7583 e 7692 kg de leite por lactação. Na tabela 3 é descrito, para além da produção de leite, a % de gordura no leite e os kg de gordura produzidos ao fim de 305 dias em lactação.
Nielen et al. (1989) verificaram em explorações alemãs, nas quais vacas leiteiras se encontravam gestantes de gémeos, um aumento da produção de leite até aos 100 dias em
13
lactação. Aos 270 dias em lactação, não foram observadas diferenças na produção de leite entre estas vacas. Do mesmo modo, Kinsel et al. (1998) observaram que vacas com partos gemelares produziram mais 2,7 kg de leite/dia no pico da lactação anterior, relativamente às vacas com partos simples, apesar de a produção total da lactação anterior não ter diferido.
Tabela 3 – Valores médios da produção de leite, % e kg de gordura aos 305 dias em lactação entre
vacas com gestação gemelar e simples.
Parâmetro produtivo Vacas com gestação
gemelar
Vacas com gestação
simples Valor de P Produção de leite (kg) 7803,20 n=333 7692,60 n=333 P<0,05 Gordura (%) 3,57 n=305 3,64 n=305 P<0,05 Gordura (kg) 277,60 n=305 279,90 n=305 P<0,05
Fonte: Adaptado de Chapin e Van Vleck (1980).
Kinsel et al., (1998) concluíram que o fator com maior contributo para o aumento da gemelaridade, durante um período superior a 10 anos, foi o constante aumento do pico de produção de leite. A elevada produção de leite e o facto de terem tido gémeos no passado aumentam o risco de gemelaridade (Nielen et al., 1989; Kinsel et al., 1998).
Em contrapartida, vacas com partos gemelares produzem menos leite na lactação subsequente, comparativamente às outras (Nielen et al., 1989). Esta redução na produção de leite pode ser resultante do aumento da incidência de desordens metabólicas, registadas na fase inicial da lactação.
Resultados de um estudo, indicam que existe um aumento na dupla ovulação asso-ciado ao aumento da produção de leite (López et al., 2005). Deste modo, vacas com produ-ções inferiores a 35 kg, entre 35-45 kg ou superiores a 45 kg por dia, durante os 14 dias antes do estro, revelaram taxas de dupla ovulação de 1,6% (2/128), 16,9% (32/189) e 47,9% (70/146; P<0,05), respetivamente, confirmando a relação entre a produção de leite e a dupla ovulação. No entanto, López-Gatius et al. (2005) não conseguiram encontrar resultados que confirmem esta relação num estudo realizado em Espanha, durante um período de cinco anos.
As taxas de gemelaridade registadas para vacas leiteiras em estudos mais recentes são superiores às de estudos mais antigos (Day et al., 1995), o que indica que a
gemelari-14
dade poderá aumentar ao longo do tempo nas explorações leiteiras, devido ao constante aumento da produção por vaca.
Até ao momento, dados que sustentem a relação entre gemelaridade, dupla ovula-ção e produovula-ção de leite são apenas de carácter associativo, não estabelecendo uma ligaovula-ção de causa-efeito, e os mecanismos fisiológicos que explicam o modo como a produção de leite pode influenciar a incidência de gemelaridade ainda não são claramente conhecidos.
2.5.7. Alimentação
Tem sido especulado que a alimentação possa ter um importante papel na incidência de gemelaridade em vacas leiteiras. O aumento dos níveis de proteína “bypass” na dieta, ou seja, a proteína que passa intacta do rúmen para o duodeno, permitem aumentar a taxa de ovulação e de gemelaridade em ovinos (Nottle et al., 1988). Apesar de ainda não ter sido provado, elevados níveis de proteína “bypass” na dieta de vacas leiteiras poderão influenciar o aumento da incidência de gemelaridade. Dados mais recentes (Vasconcelos et al., 1998) sugerem que o aumento da ingestão de alimento poderá aumentar o metabolismo hepático da inibina, como resultado de uma elevada ingestão de matéria seca e de um elevado fluxo de sangue hepático, semelhante ao que já foi descrito em ovinos (Thomas et al., 1987; Parr
et al., 1993; McEvoy et al., 1995). Com o aumento do metabolismo esteróide, poderá ocorrer
uma alteração a nível endócrino capaz de permitir que dois folículos se desenvolvam e se tornem dominantes, resultando no fenómeno de co-dominância (Wiltbank et al., 2000).
Kinsel et al., (1998) especularam que alimentando vacas leiteiras de alta produção com dietas ricas em energia poderia levar ao aumento da incidência de dupla ovulação, e por sua vez, ao aumento da taxa de gemelaridade. O efeito nutricional seria semelhante ao do “flushing” que se pratica nos ovinos (Thomas et al., 1987; Dunn e Moss, 1992), mas este mecanismo ainda carece de sustentação científica para vacas leiteiras.
2.5.8. Somatotropina bovina recombinante (rbST)
Nos EUA, desde Fevereiro de 1994, começou a utilizar-se rbST, após aprovação pe-la FDA (Bauman et al., 1999). Esta molécupe-la está descrita para ser utilizada em vacas sau-dáveis que se encontrem entre a 9ª e 10ª semana de lactação, com o objetivo de aumentar a produção de leite (em média, 5 kg/dia/vaca). Contudo, a utilização desta hormona não é permitida em Portugal ou em qualquer outro país da UE.
15
Desde a sua introdução no mercado, a relação entre rbST e gemelaridade tem gera-do alguma controvérsia entre os produtores de leite. Inicialmente, com base em estugera-dos realizados antes da sua aprovação para o mercado, foi descrito que a sua utilização poderia aumentar a taxa de gemelaridade. Burton et al. (1990) descobriram que vacas tratadas com rbST tinham uma maior incidência de gemelaridade, quando comparadas com o grupo con-trolo.
Em vários estudos, Cole et al. (1991), verificaram que as diferenças na incidência de gemelaridade dependiam da via de administração da hormona. Vacas que tinham sido inje-tadas com rbST por via intramuscular apresentaram uma maior incidência de gemelaridade (17,2%; 27/155) relativamente às vacas do grupo controlo (5,0%; 7/140; P<0,05). No entan-to, não foram detetadas diferenças na incidência de gemelaridade entre vacas do grupo con-trolo e vacas que tinham sido administradas por via subcutânea (Cole et al., 1991). Foi reali-zada mais investigação sobre este possível efeito da rbST na incidência de gemelaridade e, mais tarde, a FDA considerou que não existia o risco de aumentar a incidência de gemelari-dade devido à utilização de rbST (Collier, 1996). O mecanismo que explica o modo como cada via de administração influenciou a taxa de gemelaridade nestes ensaios não é conhe-cido.
A elevada produção de leite tem um importante efeito negativo na eficiência reprodu-tiva da vaca leiteira (Laben et al., 1982). Como já foi referido anteriormente, uma elevada produção de leite tem sido associada ao aumento da dupla ovulação e da gemelaridade em vacas leiteiras.
A rbST é, entre outros, uma substância para aumentar a produção de leite nas va-cas. Comparando a performance reprodutiva de vacas com produções semelhantes, em que um grupo é suplementado com rbST e o outro não, verificou-se que a capacidade de produ-ção de leite tinha mais influência na performance reprodutiva do que a administraprodu-ção de rbST (Cole et al., 1991). Assim, um grupo de vacas que seja suplementado com rbST deve-rá ter uma incidência de gemelaridade semelhante ao de um grupo de vacas que não te-nham sido suplementadas com rbST, mas de igual nível produtivo. Apesar dos efeitos nega-tivos da rbST na fertilidade de vacas leiteiras (Burton et al., 1990; Hemken et al., 1991; Mor-beck et al., 1991; Zhao et al., 1992; Esteban et al., 1994), tem sido revelado recentemente que melhora as taxas de conceção, quando aplicada conjuntamente com o protocolo hor-monal de sincronização da ovulação e inseminação artificial a tempo fixo (Moriera et al., 2000). Mais estudos serão necessários para se compreender os efeitos da rbST na eficácia reprodutiva e na gemelaridade em vacas leiteiras.
16
2.6. Impacto económico da gemelaridade
A incidência de gemelaridade em explorações leiteiras tende a aumentar ao longo do tempo (Day et al., 1995; Kinsel et al., 1998), e com isto as perdas económicas associadas. Eddy et al. (1991) e Beerepoot et al. (1992) estimaram que as perdas económicas associa-das à gemelaridade são entre 100 a 125$ por cada parto gemelar.
As gestações gemelares são indesejáveis nas explorações leiteiras, pois acarretam consequências negativas para os recém-nascidos e para a vaca. Vacas com gestações ge-melares estão sujeitas ao aumento do risco da perda de gestação, assim como acabam por ter uma diminuição da duração da gestação e uma maior incidência de distócia, o que acaba por se refletir num aumento dos nados-mortos, da morte perinatal dos recém-nascidos e da diminuição do peso dos vitelos à nascença (Pfau et al., 1948; Erb e Morrison, 1959; Foote, 1981; Nielen et al., 1989; Day et al., 1995; Marcusfeld-Nir, 1997; Echternkamp e Gregory, 1999). Para além disso, partos gemelares podem ser responsáveis pela maioria das doen-ças do pós-parto, como por exemplo, a retenção placentária, associada posteriormente à metrite, a qual afeta negativamente a fertilidade subsequente. Vacas que parem gémeos requerem tratamentos longos e dispendiosos no período de pós-parto e aumentam o inter-valo parto-conceção, o que acaba por se traduzir num maior risco de refugo (Nielen et al., 1989; Eddy et al., 1991; Bicalho et al., 2007). Por isso, vacas com partos gemelares repre-sentam um sério problema económico e de maneio, reduzindo a rentabilidade das explora-ções (Eddy et al., 1991; Beerepoot et al., 1992).
2.6.1. Impacto da gemelaridade no embrião/vitelo
2.6.1.1. Morte embrionária tardia e morte fetal
Depois de ocorrerem todos os acontecimentos necessários para o estabelecimento da gestação, a viabilidade embrionária é muito reduzida se estamos perante uma situação de gemelaridade. Day et al. (1995) realizaram um estudo em explorações leiteiras situadas na Califórnia, EUA, onde investigaram a ocorrência de mortalidade embrionária/fetal em 5309 gestações, por palpação transretal, entre os dias 30 e 70 após IA. Registaram 12% de morte embrionária para vacas com gestações simples e 27,9% para vacas com gestações gemelares (P<0,05). Já Eddy et al. (1991) verificaram em explorações leiteiras da Grã-Bretanha, um elevado número de abortos entre os dias 150 e 270 de gestação para vacas com gestações gemelares, comparativamente a vacas com gestações simples (8,3 % e 2,3%; P<0,01) respetivamente. Do mesmo modo, num estudo realizado em explorações
17
alemãs, a taxa de aborto foi superior para gestações gemelares (11%; n=308) do que para gestações simples (3,3%; n=958; P<0,05; Nielen et al., 1989). No entanto, em 10082 partos de nove explorações leiteiras irlandesas, com uma taxa de gemelaridade de 2,5%, a inci-dência de aborto não foi afetada pelo tipo de gestação, com 1,8% e 1,2% para gestações simples e gemelares, respetivamente (P>0,05; Mee, 1991).
López-Gatius et al. (2004) avaliaram a morte embrionária tardia e fetal precoce em vacas, através de ultrasonografia por via transretal. Começou por avaliar entre os dias 36-42 após IA, e acompanhou a gestação semanalmente, até se atingir mais de 90 dias após IA. A morte embrionária para gestações simples foi de 7,7% (n=1310), enquanto que para gesta-ções gemelares foi de 28,8% (n=132; P<0,05). Num outro estudo realizado pela mesma equipa de investigação, registaram-se 211 gestações gemelares, confirmadas entre os dias 36-42 após IA, sendo acompanhadas por ultrasonografia transretal semanalmente, até se atingir mais de 90 dias após IA, das quais 24,2% dessas gestações resultaram na perda de ambos os embriões, e em 6,2% (P<0,05) apenas um dos embriões sobreviveu (López-Gatius e Hunter, 2005).
Em bovinos, a maioria das gestações gemelares apresentam anastomoses dos va-sos sanguíneos do córion (Echternkamp, 1992). Deste modo, se um dos embriões morre, a probabilidade de o outro embrião sobreviver é reduzida (Echternkamp, 1992).
As perdas de gestações gemelares são afetadas pela localização dos embriões no útero, com elevada morte embrionária observada em gestações nas quais os embriões se encontram no mesmo corno uterino (35,2%; n=125), relativamente a gestações em que os embriões se encontram em diferentes cornos uterinos (8,1%; n=86; P<0,01; López-Gatius et
al., 2005). Do mesmo modo, Hanrahan et al. (1983) já tinham verificado que existia um
mai-or número de vacas gestantes, cuja implantação ocmai-orreu em ambos os cmai-ornos uterinos do que no mesmo corno uterino.
Um estudo recente revelou que novilhas manifestaram grandes perdas embrionárias em gestações gemelares com os embriões no mesmo corno uterino, do que em cornos ute-rinos diferentes. No entanto, o mesmo não se registou em vacas multíparas (Echternkamp et
al., 2007). Para além disto, Day et al. (1995) conduziram um estudo em explorações leiteiras
na Califórnia, EUA, onde verificaram não existir diferenças na taxa de aborto entre gesta-ções gemelares com os embriões no mesmo corno uterino (32,4%; n=187) ou em cornos uterinos diferentes (26,2%; P>0,05; n=71).
18
2.6.1.2. Mortalidade perinatal dos vitelos
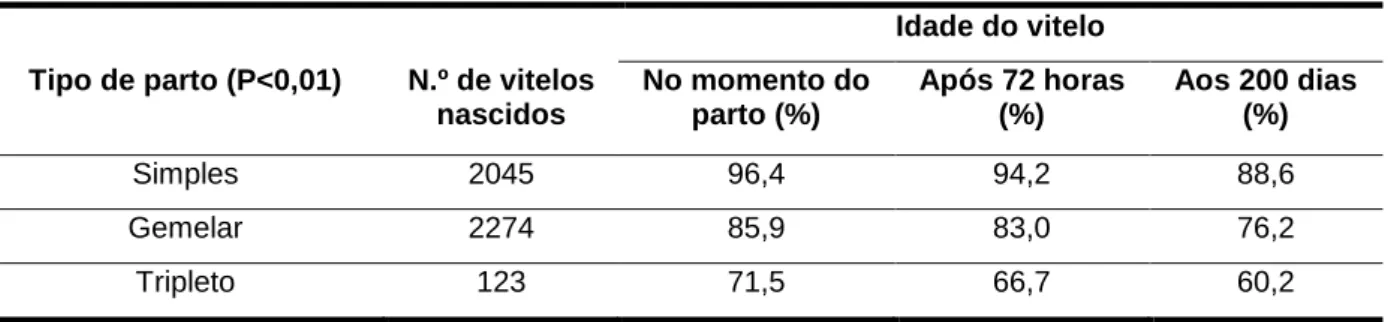
Vários fatores têm sido relacionados com a morte perinatal dos vitelos, nomeada-mente o tipo de touro, a paridade da vaca, a dificuldade de parto, o sexo, o tamanho do vite-lo e o número de vitevite-los (Wijeratne e Stewart, 1970; Hartman et al., 1974; Mee, 1991). As-sim, vários estudos revelaram que existe uma diferença significativa na mortalidade perinatal entre vitelos que nasceram dum parto simples, relativamente aos vitelos que nasceram dum parto gemelar. A probabilidade de ocorrer mortalidade perinatal é, aproximadamente, quatro vezes superior para vitelos nascidos de partos gemelares relativamente aos vitelos nascidos de partos simples, variando entre 3,2 a 6,2% para nascimentos simples, e 15,7 a 22,5% pa-ra nascimentos gemelares (Pfau et al., 1948; Erb e Morrison, 1959; Cady e Van Vleck, 1978; Nielen et al., 1989; Day et al., 1995). Este facto pode ser explicado pela maior incidência de distócia e pelo menor tamanho e peso dos vitelos (43,5 kg vs 30,6 kg), com consequente aumento da vulnerabilidade dos mesmos (Tabela 4).
Tabela 4 - Efeito do tipo de parto na viabilidade dos vitelos (%) a.
Idade do vitelo
Tipo de parto (P<0,01) N.º de vitelos
nascidos No momento do parto (%) Após 72 horas (%) Aos 200 dias (%) Simples 2045 96,4 94,2 88,6 Gemelar 2274 85,9 83,0 76,2 Tripleto 123 71,5 66,7 60,2
Fonte: Adaptado de Echternkamp e Gregory (2002). a Percentagem de vitelos vivos desde o momento do parto até aos 200 dias de vida.
2.6.1.3. Freemartinismo
Em bovinos, 90 a 95% das gestações gemelares apresentam anastomoses dos va-sos sanguíneos do córion (Jost et al., 1973). Deste modo, ocorre a fusão das membranas embrionárias de ambos os embriões, resultando numa troca de sangue entre eles. Isto acontece por volta do dia 35 de gestação, dias antes de ocorrer o desenvolvimento embrio-nário do trato reprodutivo. O desenvolvimento do trato reprodutivo ocorre primeiro nos ma-chos do que nas fêmeas. Assim, o embrião macho produz fatores endócrinos, como a hor-mona anti-Mülleriana, que causa um desenvolvimento anormal do trato reprodutor feminino. As primeiras anomalias no desenvolvimento do aparelho reprodutor da fêmea acontecem entre os dias 49 e 52 após a fecundação (Jost et al., 1972). Quando o embrião fêmea é ex-posto à hormona anti-Mülleriana, o seu aparelho reprodutor não sofre um desenvolvimento
19
completo resultando em tratos cegos, perda de células germinativas do tecido ovárico e in-capacidade de produzir estradiol. Assim sendo, fêmeas freemartins são estéreis (Müller, 1992), e não são viáveis para serem utilizadas como novilhas de reposição do efetivo.
Cerca de 92% das fêmeas gémeas heterozigóticas são freemartins (Buoen et al., 1992). Deste modo, cerca de 8% são férteis, presumivelmente porque houve uma falha na fusão das membranas embrionárias ou porque a fusão ocorreu depois do período crítico de diferenciação dos órgãos reprodutivos femininos, durante o desenvolvimento embrionário (Buoen et al., 1992). Já Gregory et al. (1996) verificaram que mais de 95% das fêmeas gé-meas heterozigóticas eram freemartins.
Alguns estudos demonstraram que a gemelaridade diminui o número de novilhas de reposição disponíveis numa exploração (Erb e Morrison, 1959; Nielen et al., 1989; Day et al., 1995). A ideia de que o freemartinismo é o principal fator para esta diminuição é falsa, pois este decréscimo no número de novilhas de reposição por parto deve-se, em grande parte, ao aumento da mortalidade perinatal dos vitelos gémeos, e de ser mais frequente o nasci-mento de vitelos do sexo masculino (Ryan e Boland, 1991). Seria de esperar que o sexo dos vitelos gémeos correspondesse a uma proporção de 50% M: 50% F.
Num estudo realizado por Silva del Rio et al. (2007), foi registado 51,9% M: 48,1% F, e apesar de terem nascido mais vitelos machos em partos gemelares, o desvio à proporção esperada não foi superior ao registado para vitelos machos nascidos em partos simples (53,3%; P>0,05). Para além disto, seria de esperar que o sexo dos vitelos por parto fosse segundo a proporção de 25% MM: 50% MF: 25% FF, mas tal não se verificou, obtendo-se uma proporção de 30,1% MM: 43,6% MF: 26,3% FF. Segundo Johansson et al. (1974), des-vios nesta proporção poderão indicar uma maior incidência de gémeos monozigóticos.
Silva del Rio et al. (2007) referem, que num estudo anterior (Silva del Rio et al., 2006), foi observada uma incidência de gémeos monozigóticos reduzida, sendo de apenas 5% de todos os gémeos nascidos. Johansson et al. (1974) estimaram que a frequência de gémeos monozigóticos variava entre 13,5 e 25% dos gémeos do mesmo sexo (MM e FF), dependendo da raça da vaca. Já Erb e Morrison (1959) estimaram que a frequência de gé-meos monozigóticos em vacas da raça Holstein-Frísia foi de 13,7% dos gégé-meos do mesmo sexo (MM e FF) e 7,4% de todos os gémeos nascidos.
O freemartinismo também representa problemas de fertilidade para os touros (Dunn
et al., 1979). A baixa fertilidade pode ser devida a hipoplasisa testicular (Bongso et al., 1981)
ou ao aparecimento de células germinativas (espermatogónias) quimeras (Rejduch et al., 2000).
20
Um outro problema adicional é a presença de fêmeas resultantes de partos simples, que mais tarde revelam ser freemartins. Isto resulta da morte do macho após a fusão das membranas embrionárias, mas antes do parto (Wijeratne et al., 1977). Contudo, como a ocorrência deste tipo de situação é muito rara, o impacto económico é muito limitado.
2.6.2. Impacto da gemelaridade nas vacas
2.6.2.1. Produção de leite na subsequente lactação
A literatura existente indica que durante a lactação, na qual as vacas se encontram gestantes de gémeos, estas produzem mais leite do que vacas com gestações simples (Me-adows e Lush, 1957; Wood, 1975; Kay, 1978; Chapin e Van Vleck, 1980; Nielen et al., 1989; Kinsel et al., 1998), apesar de terem uma lactação menos persistente (Syrstad, 1977; Nielen
et al., 1989).
A maioria dos autores concorda que as vacas com partos gemelares sofrem uma di-minuição na produção de leite na subsequente lactação.
Syrstad (1977) estimou uma diminuição na produção de leite de 195 kg na lactação subsequente ao parto gemelar, para um efetivo com uma média de produção entre 5373 e 5393 kg de leite.
Contrariamente, outros estudos revelaram um aumento na produção de leite em situ-ações de gemelaridade. Registos recolhidos durante vários anos, os quais incluíram mais de 1,5 milhões de partos, indicaram que vacas de explorações britânicas com produções entre 4000 a 6000 kg por lactação, tiveram um aumento na produção de leite de 2 a 131 kg após parirem gémeos (Wood, 1984). No entanto, este estudo não considerou vacas de elevada produção com partos gemelares.
Kay (1978) não registou diferenças na produção de leite entre partos simples e ge-melares para vacas que parissem pelo menos uma vez gémeos, durante a sua vida produti-va.
É interessante observar que os pequenos ruminantes e os bovinos de carne produ-zem mais leite após parirem gémeos. Esta descoberta tem sido associada ao maior desen-volvimento mamário causado pelos elevados níveis de hormonas presentes na circulação sanguínea durante a gestação em ovinos (Hayden et al., 1979; Manalu et al., 2000), e pelo aumento da frequência de amamentação em bovinos com gémeos (Wyatt et al., 1977; Guer-ra-Martinez et al., 1990).
21
2.6.2.2. Duração da gestação
Vários estudos revelaram que existe uma diminuição da duração da gestação em gestações gemelares, conduzindo a uma antecipação do parto com consequente diminuição do peso dos vitelos à nascença (Tabela 5). A duração da gestação em vacas com gestação gemelar é de, aproximadamente, 5 a 7 dias mais curta relativamente à de vacas com gesta-ção simples (Turman et al., 1971; Bellows et al., 1974; Anderson et al., 1982; Echternkamp e Gregory, 1999).
Tabela 5 - Duração da gestação entre vacas com gestação gemelar e simples.
Parâmetro Gestação gemelar Gestação simples Valor de P
Duração da gestação (dias) (abortos incluídos)
n= 308 268,7 ± 20,36
n= 958
278,7 ± 11,83 P<0,001 Duração da gestação (dias)
(abortos excluídos)
n= 272 274,8 ± 5,26
n= 926
280,4 ± 5,00 P<0,001 Fonte: Adaptado de Nielen et al. (1989).
Hendy e Bowman (1970) recolheram dados de várias raças de bovinos desde 1934 até 1964, e verificaram uma diminuição na duração da gestação de 1,5 a 10 dias para vacas com gestação gemelar. Estudos mais recentes indicaram que para a raça Holstein havia uma diminuição na duração da gestação de 5,2 dias (Cady e Van Vleck, 1978), 5,7 dias (Ni-elen et al., 1989) e 8 dias (Eddy et al., 1991) em vacas com gestação gemelar, relativamente a vacas com gestação simples.
As diferenças registadas no período de gestação entre vacas com gestação gemelar e simples, provavelmente, contribuirão para algumas das diferenças no peso dos vitelos (Gregory et al., 1996).
Algumas das causas que têm sido apontadas para explicar esta redução do período de gestação são o elevado peso dos fetos, visto que são mais do que um, as elevadas ne-cessidades nutricionais e uma maior resposta hormonal inerente à gestação (Hendy e Bowman, 1970; Nielen et al., 1989).
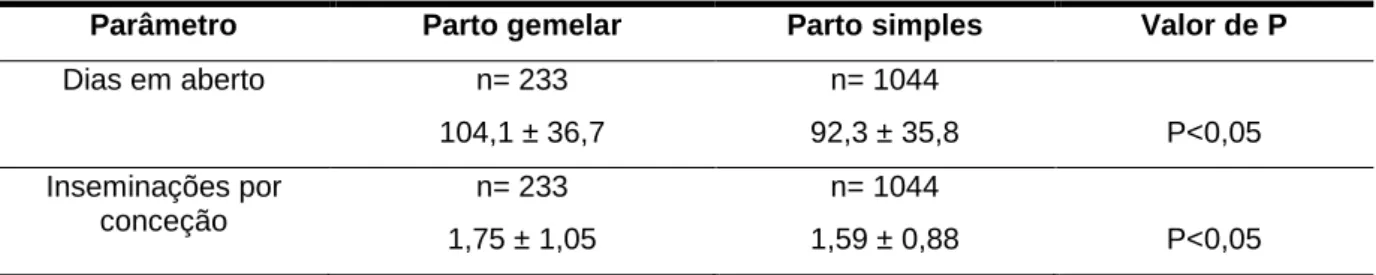
A diminuição do período de gestação poderá resultar num aumento da incidência de retenção placentária (Eddy et al., 1991), assim como limita a oportunidade de a vaca benefi-ciar das estratégias nutricionais e de maneio, adotadas no pré-parto.
22
2.6.2.3. Distócia
A ocorrência de nados-mortos e morte perinatal são mais comuns em partos distóci-cos. Por isso, uma situação de distócia reduz a viabilidade perinatal dos vitelos, especial-mente em gémeos (Gregory et al., 1996; Echternkamp e Gregory, 2002).
Um ensaio realizado em explorações leiteiras de Inglaterra e País de Gales, entre 1963 e 1965, que incluiu registos de 59804 vitelos, revelou uma incidência de distócia de 27,2%, sendo a segunda maior causa de morte dos vitelos até às oito semanas de idade (Sellers et al., 1968). Resultados de uma campanha de vigilância, realizada nos EUA, reve-laram que a incidência de distócia variava entre 16 e 28% para vacas primíparas, e entre 6 e 11% para vacas multíparas (Berger, 1994). Apesar do efeito da paridade, a incidência de distócia é maior em vacas com gestação gemelar, e varia na causa, comparativamente a vacas com gestação simples (Cady e Van Vleck, 1978; Gregory et al., 1996).
Numa outra experiência realizada num efetivo de vacas de carne cruzadas, do USDA MARC, em Clay Center, Nebraska, EUA, a incidência de distócia foi de 46,9% (n=1014) para vacas com gestação gemelar, e 20,6% (n=3370) para vacas com gestação simples (P<0,01; Echternkamp e Gregory, 1999).
Outros estudos não encontraram diferenças significativas na percentagem de vacas que necessitavam ou não de assistência, aquando do parto, entre partos simples (5,0% e 45,6%, P>0,05; respetivamente) e partos gemelares (7,0% e 46,4%, respetivamente; Eddy
et al., 1991; Mee, 1991).
Geralmente, os estudos realizados têm sido consistentes na identificação dos pro-blemas associados à gemelaridade, com exceção da incidência de distócia.
Kirkpatrick (2002) afirma que esta inconsistência pode ser explicada por dois cená-rios diferentes. Por um lado, a gemelaridade reduz a incidência de distócia causada pelo tamanho dos fetos, pois estes apresentam um menor tamanho no momento do parto, mas por outro lado, aumenta a incidência de distócia causada pelo aumento da frequência de apresentações anormais dos mesmos, no canal do parto. Segundo Mee (1991), a frequência de apresentações anormais para partos gemelares é superior à de partos simples. Para além disto, o risco de distócia em vacas com gestação gemelar poderá aumentar, devido a contrações uterinas mais fracas associadas ao enorme tamanho do útero (Cady e Van Vleck, 1978; Markusfeld, 1987).
23
2.6.2.4. Retenção placentária
A incidência de retenção placentária aumenta após um parto gemelar (Turman et al., 1971; Bellows et al., 1974; Marcusfeld, 1987; Nielen et al., 1989; Gregory et al., 1996).
Pfau et al. (1948) observaram, em vacas da raça Holstein-Frísia, uma incidência de retenção placentária superior em vacas com partos gemelares do que em vacas com partos simples. Em três diferentes estudos realizados na Holanda, Inglaterra e Irlanda, foi registada uma diferença significativa (P<0,05) na incidência de retenção placentária entre vacas de partos simples (6,1%, 2,0% e 3,3%) respetivamente, e vacas de partos gemelares (34,6%, 16,0% e 19,3%) respetivamente (Nielen et al., 1989; Eddy et al., 1991; Mee, 1991).
Echternkamp e Gregory (2002) verificaram uma incidência de retenção placentária de 1,9% para vacas com partos simples (n=3370) e de 27,9% para vacas com partos geme-lares (n=1014; P<0,01; Tabela 6).
Tabela 6 - Incidência de retenção placentária em vacas com partos simples e gemelares a.
Variável N.º de partos % de partos com retenção
placentária Tipo de parto (P<0,01)
Simples 3370 1,9
Gemelar 1014 27,9
Tipo de parto X distócia (P<0,01)
Simples sem distócia 2630 1,7
Simples com distócia 740 2,5
Gemelar sem distócia 528 23,7
Gemelar com distócia 486 32,7
Fonte: Adaptado de Echternkamp e Gregory (2002). a Dados de Echternkamp e Gregory (1999).
Echternkamp et al. (1987) observaram que a indução do parto prematuramente em vacas com gestação simples, também aumentava a incidência de retenção placentária.
Tendo em conta que uma gestação gemelar é cerca de uma semana inferior, relati-vamente a uma gestação simples, então esta poderá levar ao aumento da incidência de retenção placentária (Echternkamp e Gregory, 2002).
Outros autores especulam que um período de gestação menor e/ou um fornecimento nutricional inadequado durante a gestação, poderão ser possíveis explicações para uma maior incidência de retenção placentária verificada em vacas com partos gemelares (Eddy